










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.33 no.1 Bogotá Jan./June 2011
Estudios en ejemplares tipo de Frullania subgénero Meteoriopsis. VI. Filiación subgenérica de algunas especies asiáticas asignadas previamente al subgénero Meteoriopsis
JAIME URIBE-M.
Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C., Colombia. juribem@.unal.edu.co
ABSTRACT
A type revision of the Asiatic species previously assigned to Frullania subgen. Meteoriopsis revealed that they were erroneously included in that subgenus. Evidence presented here shows that F. seriatifolia Steph., F. ternatensis Gottsche, F. trichodes Mitt. and F. vaginata (Sw.) Nees are members of F. subg. Thyopsiella, and F. grandiclava Steph. and F. orientalis Sande-Lac. of F. subg. Frullania sect. Orientalis.
Key words. Frullania subgenus Frullania, Frullania subgenus Meteoriopsis, Frullania subgenus Thyopsiella, Hepaticae, Asia, nomenclature, taxonomic revision.
RESUMEN
La revisión de los ejemplares tipo de especies asiáticas asignadas a Frullania subgén. Meteoriopsis, reveló que ellas habían sido erróneamente incluidas en ese subgénero. Aquí se presenta evidencia que muestra que F. seriatifolia Steph., F. ternatensis Gottsche, F. trichodes Mitt. y F. vaginata (Sw.) Nees son miembros de Frullania subg. Thyopsiella, F. grandiclava Steph. y F. orientalis Sande-Lac. de F. subg. Frullania sect. Orientalis.
Palabras clave. Frullania subgénero Thyopsiella, Frullania subgenus Meteoriopsis, Frullania subgenus Frullania, Hepaticae, Asia, nomenclatura, revisión taxonómica.
Recibido: 08/12/2010
Aceptado: 10/06/2011
INTRODUCTION
Spruce (1884) described the subgenus Meteoriopsis as one of his six subgenera within Frullania, for the inception of species with pendent growth. Since then, many species have been assigned to subg. Meteoriopsis Spruce. Stephani (1911) assigned 53 species from all over the Tropics to this subgenus, including 15 species from tropical Asia and Oceania. In 1924, Stephani described eight further new species in subg. Meteoriopsis, but several of them were subsequently transferred to other subgenera.
Verdoorn (1930) in his treatment of Frullania from the Indomalayan archipelago recognized 11 species in subg. Meteoriopsis including several that were newly assigned to this subgenus. Verdoorn also presented a discussion on the morphological limits of the subgenus, and proposed several characters as being diagnostic for subg. Meteoriopsis, such as stem and branches very long, growing almost unlimited ["Stamm und Hauptäste sehr verlängert, fast unbegrenzt weiter wachsend"]. Characters associated with leaf-lobes, leaf-lobules or leaf bases were not considered.
In 1972, Hattori published the new species Frullania schusterana which he assigned to the subg. Meteoriopsis. He also discussed the merits of Meteoriopsis as a subgenus and suggested that it corresponded with sect. Vaginatae of subg. Thyopsiella Spruce (the latter as subg. "Frullania"). In 1974, however, Hattori accepted Meteoriopsis again as a good subgenus (following Verdoorn 1930) while placing F. schusterana Hatt. in subg. Thyopsiella.
Uribe & Gradstein (2003) redefined F. subg. Meteoriopsis Spruce to include species with two auricles at leaf-base and with leaves convoluted around the stem when dry (not spreading widely when moistened). Frullania peruviana Gottsche was selected as the lectotype of the group. Subsequently, Uribe (2008) monographed subg. Meteoriopsis and accepted seven species in the subgenus, all of them from tropical America (Figure 2, clade 32; Uribe 2008). Species from the Old World previously assigned to subgenus Meteoriopsis were placed in other subgenera or left unclassified. The relationships of subg. Meteoriopsis to other members of the genus Frullania were analyzed based on a phylogenetic analysis of the genus using 36 morphological characters. The analysis showed that subg. Meteoriopsis is monophyletic and sister to the monophyletic subg. Thyopsiella. Together two groups formed a monophyletic group, clade 29, consisting of 2 lineages (Uribe 2008; Fig. 2): the Thyopsiella clade with 3 species (F. atrata, F. brasiliensis, F. intumescens) and the Meteoriopsis clade with 7 species (F. convoluta, F. darwinii, F. dulimensis, F. grandifolia, F. peruviana, F. phalangiflora, F. weberbaueri).
Based on a molecular-phylogenetic analysis of the genus Frullania using four molecular markers, Hentschel et al. (2009) established a much wider subgeneric concept of F. subg. Meteoriopsis which included many previous members of subg. Thyopsiella Spruce, in three different clades. The newly defined subgenus differed from subg. Thyopsiella s.str. (including the type, F. tamarisci) by the lack of ocelli and its tropical distribution. Subgenus Meteoriopsis Spruce was subsumed in a clade named "sect. Intumescentes", containing 6 former members of Thyopsiella (F. aculeata, F. atrata, F. brasiliensis, F. intumescens, F. moritziana, F. pittieri) and 1 of genuine Meteoriopsis (F. peruviana). Thus defined, sect. Intumescentes corresponded with Frullania clade 29 recovered by Uribe (2008) on morphological grounds. However, only 4 species of clade 29 were studied by Hentschel et al. (2009), 3 of the Thyopsiella clade and 1 of the Meteoriopsis clade.
Hentschel et al. (2009) postulated that their sect. Intumescens largely corresponded to the subg. Meteoriopsis as circumscribed by Spruce (1884). This statement is erroneous. Spruce recognized four species in his subgenus Meteoriopsis: F. atrata, F. atrosanguinea, F. peruviana and F. aculeata. Of these, F. aculeata is actually a member of subg. Thyopsiella sensu Spruce, all others belong in subg. Meteoriopsis sensu Spruce (Uribe & Gradstein, 2003; Uribe, 2008; but specimens identified by Spruce as "F. atrata" proved to be misidentified F. peruviana or F. convoluta, and F. atrosanguinea appeared to be a synonym of F. peruviana). In contrast, all species included in sect. Intumescentes by Hentschel et al. (2009) are members of subg. Thyopsiella sensu Spruce with exception of F. peruviana.
Since only 1 out of 7 species of F subg. Meteoriopsis Spruce (sensu Uribe 2008) was studied by Hentschel et al. (2009), sampling in the latter study was too low to decide on the status of this subgenus. Moreover, support of the clade of F subgen. Meteoriopsis in Hentschel et al. (2009) was low (68) compared with the support of other clades, and no morphological characters were provided for the separation of the three sections in Meteoriopsis sensu Hentschel et al. Therefore, I think that it is better to keep using the concept of subgen. Meteoriopsis as defined by Uribe (2008) until more species of this group are analyzed by molecular methods.
The purpose of this present paper is to describe six Asiatic species previously assigned to subg. Meteoriopsis (F. grandiclava, F. orientalis , F. seriatifolia, F. ternatensis, F. trichodes, F. vaginata) and discuss their subgeneric affiliation.
SPECIES DESCRIPTIONS
1. Frullania (subg. Thyopsiella) vaginata (Sw.) Nees, in Gottsche, Lindenberg & Nees, Syn. Hepat. : 465 (1844). Jungermannia vaginata Swartz, Amoenitates Acad. Ed. II, vol. X, App.: 115 (1781); Frullania vaginata Dumort., 1835, Rec. Obs.: 13 (1835), nom. inval. (Art. 32.1 ICBN). Type: Java, without collector (holotype, S!). Fig. 1 A-F
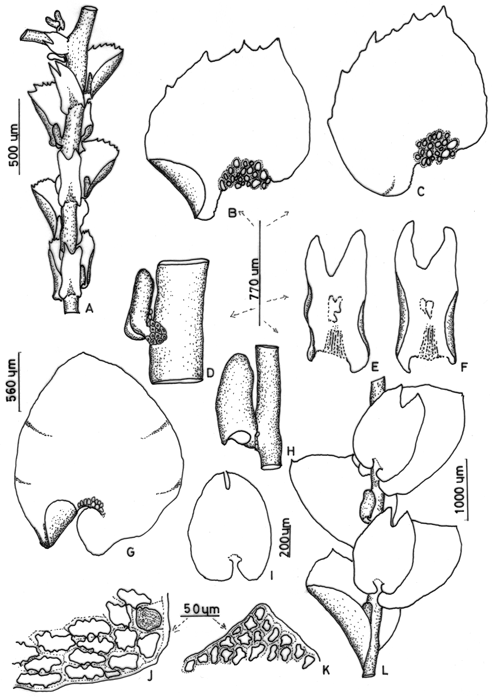
Figure 1. Frullania vaginata (Sw.) Nees - A. Part of shoot, ventral view, note the distant underleaves; B. C. Leaves of stem with dorsal base not auriculate; D. Ventral lobules with laminate stylus; E. F. Underleaves. Frullania orientalis Sande-Lac. - G. Leaf-lobe; H. Leaf-lobule with mouth and peak; I. Underleaf; J. Basal leaf cells; K. apical leaf cells; L. Part of shoot, ventral view. (A-F from the type of F. vaginata (Sw.) Nees. G-L from the type of F. oblicua Steph.).
Plants of large size, up to 20 cm long and 0.7 mm wide including leaves, dark reddish to brown, irregularly once or twice pinnate, growing more or less pendent. Branches 1-2 cm long. Stems to 90 µm wide, in cross section. Leaf-lobes remote, ovate-orbicular, obliquely spreading, with strongly incurved ventral margin, plane, ovate, asymmetrical, 740 x 600 µm, apex subacute (short acuminate on branch leaves), dorsal margins dentate, dorsal base auriculate, ventral base not auriculate, insertion line slightly curved. Leaf cells: apical cells 10-12 x 8-9 µm, median cells 12-14 x 9-11 µm, basal cells 23-33 x 8-12 µm, all cells with thick, sinuous walls and with trigones and intermediate thickenings, pale orange, basal ocelli distinct, 10-17 per leaf-lobe. Lobules cylindrical to clavate, 220-230 x 100-110 µm, contiguous and almost parallel to the stem, with arched and crenulate mouth. Stylus composed of a row of 3 uniseriate cells and of a rounded disc of 6-8 cells long and 5 cells wide. Underleaves ovate-rectangular, distant, slightly wider than the stem, 540-560 x 230-240 µm, margins strongly recurved, bifid to 1/3 the length, segments subtriangular, apex acute or acuminate, diverging, base short auriculate, insertion line straight. Branch appendages: first branch underleaf divided to the base into a lanceolate, bifid segment and a saccate segment, first leaf transformed into two saccate lobules. Plants dioicous. Androecia not seen. Gynoecia lateral on primary branches, or terminal on stems, the bracts and bracteoles in three series, bracts unequally bifid, the segments acute or acuminate, margin dentate. Perianth not seen.
Distribution Java, Sumatra. Stephani (1911) reported this species from Borneo and New Guinea, and Menzel (1988) listed the species in his Borneo catalogue based on literature compilation. However, according to Verdoorn (1930) and Hattori (1976a) this species is restricted to Sumatra and Java, and is absent of Borneo and New Guinea.
Comments Verdoorn (1930) assigned F. vaginata to subg. Meteoriopsis, mostly on the basis of its long stems, whereas Hattori (1976a) treated it as a member of Frullania subg. Thyopsiella. Uribe (2008) excluded the species from subg. Meteoriopsis and did not assign it to subgenus by lack of study of the type. Following examination the holotype, I now confirm that the species belongs to subg. Thyopsiella.
2. Frullania (Frullania) orientalis Sande Lac. Nederl. Kruidk. Arch. IV: 94 (1855). Type: Java, Teysmann s. n. (holotype, L!). Fig. 1 G-L
Frullania obliqua Steph., Sp. Hepat. IV: 616 (1911). Type: Nova Guinea, Moroka, Loria s.n. (holotype, G!).
Plants of medium to large size, up to 10 cm long and 2 mm wide including leaves, loosely branching, branch long, to 5 cm long, distant, dark reddish brown to almost black, irregularly pinnate, growing more or less pendent. Stems to 200-500 µm wide in cross section. Leaf-lobes distant, rounded, entire, convolute around the stem when dry and wet, plane, ovate, symmetrical, 1850-1900 x 1550-1575 µm, apex obtuse or rounded, to slightly acute, margins entire, dorsal base auriculate or sometimes subtruncate, ventral base auriculate, auricles sometimes superposing each other, mostly the leaves subtending branches, insertion line arched. Leaf cells: apical cells 12.5-17.5 x 10-12.5 µm, median cells 30-42.5 x 10-17.5(-20) µm, basal cells 50-67.5 x 35-45 µm, all cells with thick, sinuous walls and with trigones and intermediate thickenings. Leaf-Lobules cylindrical to clavate, often forming at mouth a distinct short beak, 210-240 x 60-90 µm, contiguous and parallel to the stem, occasionally slightly inclined to the stem. Stylus filiform, erect, linear, three cells long, with a terminal, 50 µm long slime papilla. Underleaves ovate, distant, slightly wider than the stem, 270-560 µm x 220-360 µm, margins recurved, undulate, bifid to 1/4 the length, segments acute or obtuse, diverging, base short auriculate, insertion line straight. Branch appendages: first branch underleaf divided to the base into a lanceolate segment and a saccate segment, first leaf transformed into two saccate lobules. Plants dioicous. Androecia not seen. Gynoecia terminal on the stem, the bracts and bracteoles in three series, bracts unequally bifid, the segments acute or acuminate, entire. Perianth pyriform, 500 µm long, with three keels, the ventral keel narrow.
Distribution - New Guinea, Taiwan, Philippines, Sulawesi, Borneo, Java, Vietnam, Malayan Peninsula, Sumatra, Sri Lanka, and other, smaller Asian and Pacific islands (Hattori, 1976b). In addition, the species has been reported from Fiji (Campbell, 1971), China (Yunnan) (Piippo et al., 1998) and India (Parihar, 1962).
Comments - This species was treated by Verdoorn (1930) within a new section "Orientales", along with F. nigricaulis (Reinw., Blume & Nees) Nees. However, Hattori (1972) transferred this species to subg. Frullania ("Trachycolea") sect. Ornitocephalae Verd. Based on study of the type material, I confirm that the species belongs to subg. Frullania. Following Hentschel et al. (2009), the species may be placed in subg. Frullania sect. Orientales.
3. Frullania (subg. Frullania) grandiclava Steph., Sp. Hepat. 4: 676 (1911). Type: New Caledonia, VII 1909, Le Rat s.n. (holotype, G!). Fig. 2 A-H
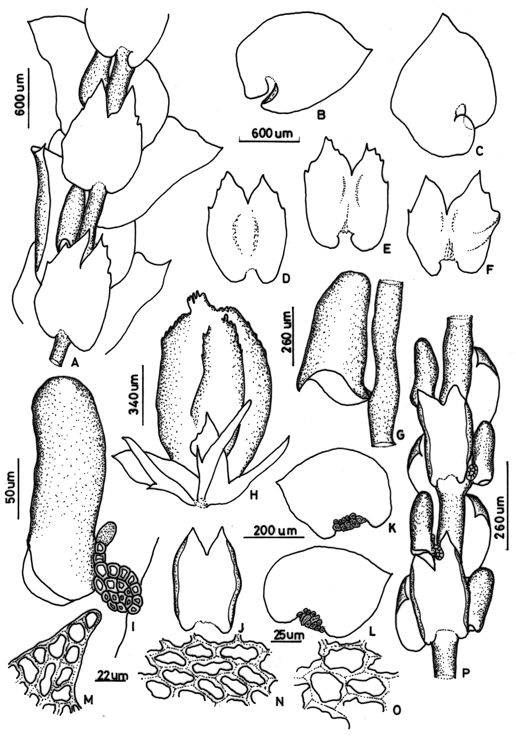
Figure 2. Frullania grandiclava Steph. - A. Part of shoot, ventral view; B. C. Leaves-lobes; D, E, F. Underleaves; G. Leaf-lobule, with mouth and peak; H. Perianth. Frullania seriatifolia Steph. I. Leaf-lobule, with stylus laminate; J. Underleaf; K. L. Leaves with basal ocelli; M. Apical leaf cells; N. medial leaf cells; O. basal leaf cells; P. Part of shoot, ventral view. (A-H from the type of F. grandiclava Steph.; I-P from the type of F. seriatifolia Steph.).
Plants slender, medium to large size, to 10 cm long and 2 mm wide including leaves, reddish brown. Irregularly pinnate, branches widely spreading, usually to 5 mm long, branches remote. Stems to 175 µm wide in cross section. Leaf-lobes loosely imbricate to contiguous, slightly squarrose, convoluted around the stem when dry and spreading when wet, ovate, 1250-1325 - 1000-1300 µm, margin entire, incurved to ventral side, apex acute to short apiculate, base cordate, dorsal base widely auriculate and ventral base auriculate or truncate, ventral auricle shorter than dorsal one. Insertion line curved. Leaf-lobe cells: apical cells 12-20 x 10-15µm, median cells 25-37 x 17-20µm and basal cells 35-45 x 25-32 µm, cell wall thick, sinuous, trigones conspicuous, intermediate thickenings present, walls of basal cells more reddish than the others cells. Leaf-lobules clavate to cylindric, 520-600 x 250-370µm, with mouth rostrate with a long beak, contiguous and parallel to the stem. Stylus short, filiform, five cells long, with terminal slime papilla. Underleaves distant, elliptical or subcuadrate, concave, bifid to 1/3 of length, 1000-1125 x 770-825 µm, sinus wide, lobes triangular with acute apex, one or two marginal tooth, base cordate, insertion line very short and straight. Branch appendages: first branch underleaves (BUL1) ventral segment laminar, ovate, bifid to ½ of length, insertion line very long and curved. First branch leaf-lobes not seen. Plants dioicous. Androecia not seen. Gynoecia on short lateral branches, bracts and bracteoles entire and bifids. Perianth 2/3 exerted, ovate to elliptical, 1100-1200 x 600-650 µm, with three keels, ventral keel prominent, long beak at apex, rough by scattered tubercles or scale-like outgrowths.
Distribution - Known only from the type locality from New Caledonia.
Comments: Verdoorn (1930) and Hattori (1977) treated F. grandiclava as a synonym of F. capillaris without justification. In my study I found that F. grandiclava differs from F. capillaris as described by Hattori by its toothed underleaves, leaves longer than wide and leaf-lobules smaller, therefore F. grandiclava is treated here as a separate species, not as a synonym of F. capillaris.
By its underleaves with a toothed margin, lobule with a rostrate mouth and long beak, and perianth with scattered tubercles, F. grandiclava is clearly a member of subg. Frullania sect. Orientalis.
4. Frullania (subg. Thyopsiella) seriatifolia Steph., Hedwigia 33:167 (1894). Type: Nova Guinea, Mt. Yule, without collector (holotype, G! 13663). Fig. 2 I-P
Frullania yulensis Steph., Spec. Hepat. IV: 536 (1911). Type: New Guinea, Mt. Yule, McGregor s.n. (holotype, G!).
Plants slender, short size, to 1,5 cm long and 300 µm wide including leaves, reddish brown. Pinnately branched; widely spreading, usually to 2 mm long, branches remote. Stems to 100 µm wide in cross section. Leaf-lobes distant, patent, ovate to suborbicular, plane, 450-490 x 330-350 µm, margin entire, apex acute to acuminate, dorsal base widely auriculate and ventral base truncate. Insertion line straight. Leaf-lobe cells: apical cells 7-12 x 5-7µm, median cells 12-20 x 5-8µm and basal cells 18-22 x 6-10 µm, cell wall thick, sinuous, trigones conspicuous, intermediate thickenings present, walls of basal cells more sinuous than the others cells. Leaf-lobules cylindric to long cylindric, 207-210 x 62-65µm, distant to the stem, forming an angle of 45° to the stem (patent). Stylus short, filiform, three cells long, with terminal slime papilla and laminal portion at base. Underleaves long distant, subcuadrate to rectangular, concave, margins strongly recurved, bifid to 1/3 to 1/4 of length, slightly wider than the stem, 310-350 x 200-210 µm, sinus narrow, lobes triangular with subacute apex, base truncate, insertion line long and straight. Branch appendages: first branch underleaves (BUL1) ventral segment laminar, ovate, bifid to 1/3 of length. First branch leaf-lobes not seen. Plants dioicous. Androecia on short lateral branches, capitate, three pair of bracts. Gynoecia not seen. Perianth not seen.
Distribution - Known only from type locality, New Guinea.
Comments - Stephani (1894, 1911) and Uribe (2008) placed this species in subg. Diastaloba, Verdoorn (1930) assigned it to Meteoriopsis and Hattori (1973) to subg. Thyopsiella. Hattori also concluded that F. yulensis Steph. is a synonym of F. seriatifolia. My study of the type specimens confirm the conclusions of Hattori (1973).
5. Frullania (subg. Thyopsiella) ternatensis Gottsche, in Gottsche, Lindenberg & Nees, Syn. Hepat. : 465 (1844). Type: Java, Hasskarl s.n. (lectotype nov., G!, designated here). Paralectotypes: Ternate, Reinwardt s.n.; Java, Belanger s.n. Fig. 3 A-F
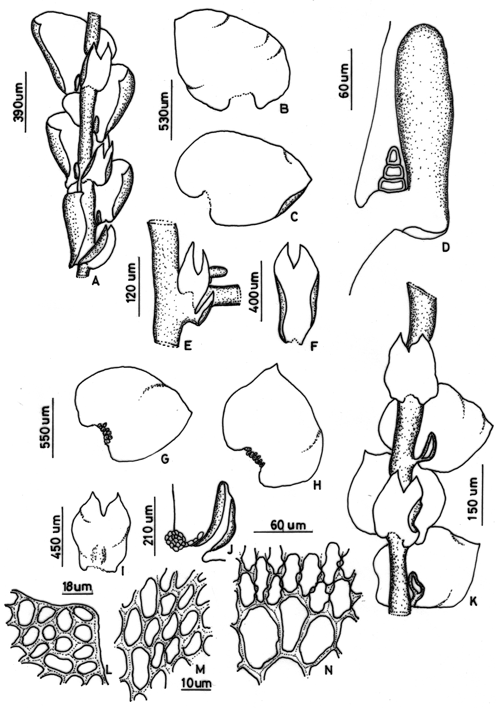
Figure 3. Frullania ternatensis Gottsche. - A. Part of shoot, ventral view; B. C Leaves; D. Leaf-lobule, with triangular stylus; E. First branch underleaf and leaf; F. Underleaf. Frullania trichodes Mitt. - G. H. Leaves; I. Underleaf. J. Leaf-lobule, with laminate stylus; K. Part of shoot, ventral view; L. Apical leaf cells; M. Medial leaf cells; N. Basal leaf cells. ( A-F from the type of F. ternatensis Gottsche. G-N from the type of F. tenuicaulis Mitten).
Frullania pendula Mitt., J. Proc. Linn. Soc., Bot. 5: 122 (1861). Type: Sri Lanka, Gardner 134 (holotype, NY!; isotypes, BM!, FH!).
F. concava Horik., Journ. Sci. Hiroshima Univ., ser. B, div. 2, 2: 238, pl. 18:19-23 (1934). Type:Taiwan: prov. Taitô, between Shinsuiei and Shûchôkyôkai, Jan. 1933, Y. Horikawa 10663b.
Plants of medium or large size, 8 to 20 cm long and 0.7 mm wide including leaves, brown, irregularly once or twice pinnate, growing more or less pendent. Branches 1-3 cm long. Stems to 60 µm wide, in cross section. Leaf-lobe more or less imbricate, ovate or triangular-ovate, convoluted around the stem when dry and spreading when wet, with strongly incurved ventral margin, plane, asymmetrical, 1200-1250 x 900-950 µm, apex acute (short acuminate on branch leaves), dorsal base auriculate, ventral base not auriculate, insertion line straight or slightly curved. Leaf cells: apical c ells 11-12 x 7-9 µm, median cells 18-25 x 8-13 µm, basal cells 27-50 x 12-23 µm, all cells with thick, sinuous walls and with confluent trigones and intermediate thickenings. Leaf-lobules cylindrical to clavate, 220-230 x 60-70 µm, 3 times as long as wide, contiguous and almost parallel to the stem, with arched and crenulate mouth. Stylus triangular, composed of a row of 3 uniseriate cells. Underleaves oblong, distant, wider than the stem, 600-800 x 200-400 µm, margins strongly recurved, bifid to 1/4 the length, segments triangular, apex acute or acuminate, diverging, insertion line straight. Branch appendages: first branch underleaf divided to the base into a lanceolate, bifid segment and a explanate segment, first leaf transformed into one saccate lobule. Plants dioicous. Androecia capitate, on lateral short branches, 1-3 pairs of bracts. Gynoecia lateral on primary branches, or terminal on stems, the bracts and bracteoles in 1-3 series. Perianth oblong, exserted, 1500 x 550 µm, 3-keeled, smooth, apex obtuse (Hattori, 1973).
Distribution - Taiwan, Philippines, Sulawesi, Borneo, Java, Vietnam, Sumatra, Malacca, Sri Lanka.
Comments: According to Verdoorn (1930) the inclined, triangular stylus (Fig. 3D) is characteristic of this species. The different sizes of the auricles at the dorsal and ventral bases of the leaf-lobe, and the shape of the lobule indicate that the species belongs in subg. Thyopsiella (Verdoorn, 1930; Hattori, 1973).
6. Frullania (subg. Thyopsiella) trichodes Mitt., Bonplandia 10: 19 (1862). Type: Fiji, Seeman s.n. (holotype, NY!). Fig. 3 G-N
Frullania vethii Sande Lac., Reiz. Onderz. Sumatra-Exped. 4 (2), Flora: 44 (1884). Type: Indonesia, Sumatra, "Barisan Gebirge bei Alahan Pandjang", 1877-1879, Veth & van Hasselt s.n. (holotype, L!).
Frullania tenuicaulis Mitt., Trans. Linn. Soc. London, Bot. 3: 203 (1899), syn. fide Hattori (1982). Type: China, Prov. Quantung, without collector (holotype, NY!);
F. picta Steph., Hedwigia 3: 166 (1894), syn. fide Kamimura (1961). Type: Indonesia, Ambon, Wawani Hila, G. Karsten s.n. (holotype, G).
F. grebeana Steph., Sp. Hepat. 4: 537 (1911), syn. fide Kamimura (1961). Type: Indonesia, Borneo, Tamelayang, without collector (holotype, G!).
F. merrillana Steph., Sp. Hepat. 4: 613 (1911), syn. fide Kamimura (1961). Type: Philippines, Luzon, without collector (holotype, G!).
F. rubrimacula Goebel, Ann. Jard. Bot. Buitenz. 39: 51 (1928), syn. fide Kamimura (1961). Type: Indonesia, Sumatra, Fort de Kock, 1925, Goebel s.n.
Plants slender, medium size, to 4-5 cm long and 150 µm wide including leaves, deeply reddish brown. Irregulary pinnate. Branches frecuent, Frullania type, 2-4 mm long; widely spreading, usually to 2 mm long, branches remote. Stems to 100-120 µm wide in cross section. Leaf-lobes distant, widely ovate, concave, 900-975 x 1100-1125 µm, margin entire, apex acute or rounded, dorsal base slightly auriculate and ventral base truncate. Insertion slightly curved. Leaf-lobe cells: apical cells 15-18 x 10-12µm, median cells 20-25 x 12-15 µm and basal cells 35-60 x 30-50 µm, cell wall rather thick, sinuous, trigones conspicuous, intermediate thickenings present, walls of basal cells more sinuous than the others cells; basal ocelli well developed. Leaf-lobules saccate or sometimes explanate, when saccate cylindric to clavate-cylindric, 220-250 x 80-110 µm, distant to the stem. Stylus short, filiform, three or four cells long, with terminal slime papilla and laminal portion rounded at base to 18 cells. Underleaves long distant, subcuadrate to rectangular, concave, margins entire, bifid to 1/3 to 1/4 of length, wider than the stem, 500-750 x 250-550 µm, sinus wide, lobes triangular with acute apex, base truncate, insertion line long and straight. Branch appendages: first branch underleaves (BUL1) ventral segment laminar, rectangular, bifid to 1/3 of length. First branch leaf-lobes transformed into two saccate lobules. Plants dioicous. Androecia on short lateral branches, capitate to apicate, two pair of bracts. Gynoecia on lateral branches, spicate, 1000 x 250 µm, three or four pair of bracts. Perianth not seen.
Distribution - Solomon Islands, New Guinea, Japan, China, Taiwan, Philippines, Borneo, Java, Sumatra, Thailand (Sornsamran & Thaithong, 1995), Myanmar, Sri Lanka (Onraedt, 1981).
Comments: Hattori (1975) suggested that Frullania vaginata and F. vethii may be considered as locally differentiated sibling taxa at a subspecific level. However, my examination of the type specimens indicates that F. vethii is morphologically identical to F. trichodes and should be treated as a synonym of the latter species. Frullania trichodes belongs in subg. Thyopsiella and is close to F. vaginata. The latter species can be distinguished by the dentate leaf margin (mostly on dorsal side), while in F. trichodes the margins of the leaves are entire. Frullania trichodes is very variable in several characters, which is reflected in the high number of synonyms; although its identity is not completely resolved yet, I treat it here as a distinct species.
ACKNOWLEDGMENTS
I would like to thank the National University of Colombia for supporting my studies, to the Deutsche Akademischer Austausdienst (DAAD) for the scholarship that allowed me to work during the first half of 2002 at the Department of Systematic Botany of the University of Göttingen. Special thanks to Robbert Gradstein for his valuable contribution and comments on the manuscript. I also thank the curators of the cited herbaria (BM, FH, G, L, NY and S) for the loan of type specimens. Thanks to Matt von Konrat for comments to the manuscript, to Jochen Heinrichs for providing literature, and to Carlos Parra for English corrections.
LITERATURE CITED
1. CAMPBELL, E.O. 1971. Liverworts collected in Fiji by A. C. Smith and W. Greenwood. J. Roy. Soc. New Zealand 1:7-30. [ Links ]
2. GOTTSCHE, C. M., J. B. W. LINDENBERG & C.G. NEES VON ESENBECK. 1844-47. Synopsis Hepaticarum, p. i-xvi, 1-834. Leipzig. [ Links ]
3. HATTORI, S. 1972. Novae Guianae Hepaticae Schusteranae, III. Species Novae Frullaniaceae, J. Hattori Bot. Lab. 36:411-443. [ Links ]
4. HATTORI, S. 1973. Notes on asiatic species of the genus Frullania, Hepaticae. II. J. Hattori Bot. Lab. 37: 55-84. [ Links ]
5. HATTORI, S. 1973. Notes on asiatic species of the genus Frullania, Hepaticae. IV. J. Hattori Bot. Lab. 37: 121-152. [ Links ]
6. HATTORI, S. 1974. Notes on asiatic species of the genus Frullania, Hepaticae. V. J. Hattori Bot. Lab. 38: 185-221. [ Links ]
7. HATTORI, S. 1975. Notes on asiatic species of the genus Frullania, Hepaticae.VII. J. Hattori Bot. Lab. 39: 227-313. [ Links ]
8. HATTORI, S. 1976a. Notes on asiatic species of the genus Frullania, Hepaticae. IX. Bull. Natl. Sci. Mus., Tokyo, B 2(1):7-22. [ Links ]
9. HATTORI, S. 1976b. Notes on asiatic species of the genus Frullania, Hepaticae. X. J. Hattori Bot. Lab. 40: 461-507. [ Links ]
10. HATTORI, S. 1977. Dr. Hürlimann's collection of New Caledonian Frullaniaceae. J. Hattori Bot. Lab. 43:409-438. [ Links ]
11. HATTORI, S. 1982. A synopsis of New Guinean Frullania, Hepaticae. J. Hattori Bot. Lab. 51: 203-271. [ Links ]
12. HENTSCHEL, J., M. J. VON KONRAT, T. PÓCS, A. SCHÄFER-VERWIMP, A. J. SHAW, H. SCHNEIDER & J. HEINRICHS. 2009. Molecular insights into the phylogeny and subgeneric classification of Frullania Raddi (Frullaniaceae, Porellales). Mol. Phylogenet. Evol. 52:142-156. [ Links ]
13. KAMIMURA, M. 1961. A monograph of Japanese Frullaniaceae. J. Hattori Bot. Lab. 24:1-191. [ Links ]
14. MENZEL, M. 1988. Annotated catalogue of the Hepaticae and Anthocerotae of Borneo. J. Hattori Bot. Lab. 65: 145-206. [ Links ]
15. ONRAEDT, M. 1981. Bryophytes récoltées au Sri Lanka (Ceylan). V. J. Hattori Bot. Lab. 50: 191-216. [ Links ]
16. PARIHAR, N. S. 1962. An Annotated revised census of Indian hepaticae. Allahabad University 1961-62:1-56. [ Links ]
17. PIIPPO, S., X.-L. HE, T. KOPONEN, P. L. REDFEARN JR. & J.-X. LI. 1998. Hepaticae from Yunnan, China, with a checklist of Yunnan Hepaticae and Anthocerotae. J. Hattori Bot. Lab. 84: 135-158. [ Links ]
18. SORNSAMRAN, R. & O. THAITHONG. 1995. Bryophytes in Thailand. OEPP Biodiversity Series 2: 1-231. [ Links ]
19. SPRUCE, R. 1884. Hepaticae of the amazon and of the andes of Peru and Ecuador. Trans. & Proc. Bot. Soc. Edinburgh 15: i-xi, 1-308. [ Links ]
20. STEPHANI, F. 1894. Hepaticarum species novae VI. Hedwigia 33: 137-169. [ Links ]
21. STEPHANI, F. 1911. Species Hepaticarum, vol. 4. Geneve. [ Links ]
22. STEPHANI, F. 1924. Species Hepaticarum, vol. 6. Geneve. [ Links ]
23. THIERS, B. M. 1983. Index to the genera and species of hepaticae described by William Mitten. Brittonia 35(3): 271-300. [ Links ]
24. URIBE-M., J. & S. R. GRADSTEIN. 2003. Type studies on Frullania subgenus Meteoriopsis (Hepaticae). I. The lectotypification of the genus Frullania, F. subgen. Thyopsiella and F. subgen. Meteoriopsis, and some species transferred from subgen. Meteoriopsis to subgen. Thyopsiella. Cryptog. Bryol. 24 (3): 193-207. [ Links ]
25. URIBE-M., J. 2008. Monografía de Frullania subgénero Meteoriopsis (Frullaniaceae, Marchantiophyta). Caldasia 30(1):49-94. [ Links ]
26. VERDOORN, F. 1930. Die Frullaniaceis VII. Die Frullaniaceae der indomalesischen Inseln. Ann. Bryol., Suppl. 1:1-187. [ Links ]

