










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.33 no.1 Bogotá Jan./June 2011
Reproducción continua bajo un régimen de precipitación bimodal en el anolis Andino (Anolis mariarum) de Antioquia, Colombia
LAURA C. RUBIO-ROCHA
BRIAN C. BOCK
VIVIAN P. PÁEZ
Departamento de Biología, Edificio 320, Ciudadela Universitaria Meléndez, Universidad del Valle. Cali, Colombia/Department of Ecology and Evolutionary Biology, 569 Dabney Hall, 1416 Circle Drive, University of Tennessee. Knoxville, Tennessee, 37996-1610, USA. lacaruro@gmail.com
Instituto de Biología, Universidad de Antioquia, AA 1226. Medellín, Colombia. brianbock1@gmail.com, vivianpaez1@gmail.com
ABSTRACT
Reproductive activity was studied in two populations of Anolis mariarum during rainy and dry season months in the Antioquia department of Colombia. Minimum size at maturity was comparable at the two sites for both males (37-39 mm SVL) and females (44 mm SVL). At the population level, reproduction was continuous, with the majority of adult males and females reproductively active even during the dry season months. Juvenile size distributions also were uniform across seasons, consistent with the conclusion that recruitment is not pulsed in these populations. However, there was some evidence that certain females may lower their reproductive rates during the dry season, especially at the site receiving less total annual precipitation (1700 mm). These results further support accumulating evidence that populations of Anolis species inhabiting the "climatic equator" region, where the annual precipitation regime is bimodal, are capable of maintaining continuous reproduction even when annual precipitation amounts are relatively low. In contrast, Anolis populations in areas receiving comparable amounts of annual precipitation during a single rainy season tend to cease reproductive activity during the longer dry season each year.
Key words. Bimodal precipitacion, Climatic equator, Polychrotidae, Reproductive cycles, Sexual maturity.
RESUMEN
Se estudió la actividad reproductiva de dos poblaciones de Anolis mariarum durante meses pertenecientes a las estaciones seca y lluviosa. El tamaño mínimo de madurez sexual fue comparable en los dos sitios, tanto para machos (37-39 mm LRC), como para hembras (44 mm LRC). Al nivel de población, la reproducción fue continua, encontrándose la mayoría de los adultos machos y hembras reproductivamente activos incluso en los meses de la estación seca. La distribución del tamaño de los juveniles también fue uniforme a lo largo de todos los meses, consistente con la conclusión de que el reclutamiento no es estacional en estas poblaciones. Sin embargo, se encontró evidencia de que algunas hembras pueden reducir su tasa reproductiva durante la estación seca, especialmente en la localidad que recibe menor precipitación total anual. Estos resultados apoyan la evidencia acumulada de que poblaciones de especies de Anolis habitando la región del "ecuador climático", en donde el régimen de precipitación anual es bimodal, son capaces de mantener una reproducción continua incluso cuando la cantidad de precipitación anual es relativamente baja, mientras que poblaciones de Anolis en áreas que reciben cantidades comparables de precipitación anual, pero durante una sola estación de lluvias, tienden a interrumpir la actividad reproductiva durante la estación seca prolongada cada año.
Palabras clave. Precipitación bimodal, Ecuador climático, Polychrotidae, Ciclos reproductivos, Madurez sexual.
Recibido: 01/09/2010
Aceptado: 10/01/2011
A number of lizard lineages have evolved a suite of life history traits that includes small body size, relatively little growth after maturity, invariant clutch size of one or two eggs per clutch, and short inter-clutch intervals (Vitt 1986, Shine & Greer 1991). While most other lizard species exhibit seasonal reproduction (Vitt & Caldwell 2009), many of the species that have adopted this life history strategy reproduce continuously throughout the year. Thus, comparison of the reproductive ecologies of lizard species with small fixed clutch sizes, such as geckos or anoles, offers an opportunity to examine hypotheses concerning the factors that contribute to the evolution of seasonal vs. continuous reproduction.
Watling et al. (2005) reviewed the literature on 32 populations of 17 species of mainland neotropical lizards with fixed clutch sizes, including eight Anolis species, to inspect for environmental effects on reproductive phenology. They concluded that, after controlling for phylogenetic effects, seasonal breeding was significantly related to the annual amount of precipitation at a site, but not to the duration of the dry season. For example, in the Anolis species they examined, seven of eight populations from habitats receiving more than 2600 mm of precipitation a year exhibited continuous reproduction, while all eight populations experiencing less than 2600 mm of annual precipitation exhibited seasonal reproduction (Watling et al. 2005).
However, all the Anolis populations included in their meta-analysis were Central American, where a well defined primary dry season occurs, with a secondary decline in precipitation during a "veranillo" (or midsummer drought) in the middle of the rainy season (Rand & Rand 1982, Magaña et al. 1999, Small & de Szoeke 2007). This second dry season never approaches the primary dry season in intensity or duration, and Central American Anolis that reproduce seasonally either reduce or completely halt reproductive effort only during the primary dry season (Watling et al. 2005 and references within). As latitude decreases, however, the intertropical convergence zone produces the "climatic equator" region (Borchert et al. 2005), where genuinely bimodal patterns of annual precipitation occur. The climatic equator in South America occurs slightly north of the geographic equator. In northwest South America it is centered at approximately 4.5 °N, while in northeast South America it runs SW to NE through Guyana from 2-6 °N (Borchert et al. 2005).
Ramírez-P et al. (1991) studied the reproductive phenology of a population of Anolis heterodermus located at 4.8 °N (in the climatic equator zone) in Cundinamarca, Colombia, where only 880 mm of annual precipitation falls in two distinct wet seasons (April to June and September to November). Contrary to the predictions of the analysis of Watling et al. (2005), reproduction in this population was continuous. More recently, Ardila-M et al. (2008) reported evidence of reproduction occurring during both the end of the second wet season of 2004 and the first dry season of 2005 (November to February) in a population of Anolis tolimensis located at 4.6 °N in Cundinamarca, Colombia where a mean of 1005 mm of precipitation occurs each year (D. Ardila, pers. comm.), further suggesting that selection acts differently on reproductive phenology in Anolis species from the climatic equator region.
Here we describe the reproductive phenology of two populations of Anolis mariarum (Polychrotidae) located at 6.2-6.4 °N in Antioquia, Colombia, where there also is a bimodal precipitation regime, to further examine the effects of the amount and distribution of precipitation on the reproductive phenology of Anolis lizards.
MATERIALS AND METHODS
Anolis mariarum occurs in low vegetation in open (usually human disturbed) areas in the Cordillera Central of northern Colombia, from 1300 to 2700 m elevation (Páez et al. 2002, Palacio-B et al. 2006). In this study, we documented reproductive activity in a population inhabiting a roadside area in the municipality of San Pedro de los Milagros (hereafter, "San Pedro", 06°25'07'' N; 75°35'55'' W) at 2400 m elevation that has a mean annual temperature of 14 °C and mean annual precipitation of 1700 mm. We also studied reproductive phenology in a second population inhabiting a suburban neighborhood (San Antonio de Prado) in the municipality of Medellín (hereafter, "San Antonio", 06°11'02''N 075°40'09 W) at 2200 m elevation with a mean annual temperature of 16 °C and mean annual precipitation of 2600 mm (climatological data provided by Empresas Públicas de Medellín). We collected individuals from October 2008 (middle of the second rainy season of the year) until March 2009 (beginning of the first rainy season of the year), for a total of three rainy and three dry season months. Twenty individuals (in approximately equal sexual proportions) were hand captured each month from each population and sacrificed by cardiac injection of 2% xilocain. For each individual, body mass was weighed (+ 0.001 g) on a digital balance and snout-vent length (SVL) was measured (+ 0.01 mm) using a digital caliper. The body cavity was opened with a mid-ventral incision and the following data were recorded with the same precisions as above: for females, ovarian mass, diameter of the largest ovarian follicle, number of vitellogenic and non-vitellogenic follicles, number, mass, length and width of oviductal eggs, and presence or absence of abdominal fat bodies; for males, left testicle length, width, and mass, presence or absence of epididimal convulations, and presence or absence of abdominal fat bodies (Serrano-C et al. 2007). All specimens were fixed in 10% formalin, preserved in 70% ethanol, and deposited in the Museo de Herpetología of the Universidad de Antioquia (MHUA-R 11809-11903, 11952-11965, 12125-12237).
Studies of reproductive cycles ideally should establish minimum body sizes at first reproduction for both males and females, to help avoid confusing sexually immature individuals with individuals that have interrupted their reproductive effort. To more critically establish size at first reproduction in these populations, we selected individuals from a range of body sizes of each sex in each population and used histology to examine their reproductive tracts (32 males and 29 females from San Pedro and 28 males and 23 females from San Antonio). We removed the left reproductive tract (ovaries and oviduct for females and testicles and epididimus for males) from each individual. These tissues were fixed in Bouin's solution, embedded in paraffin, sectioned into 3 µm layers, and stained with hematoxylin-eosin. These histological preparations were used to corroborate the reproductive states previously determined by examination of gross anatomy. In males, spermatogenesis was confirmed by the presence of secondary spermatocytes (spherical cells with a set of homologous chromosomes still joined by the chromatids), spermatids (elongated cells that still had not completed the spermatogenesis process), or sperm cells (with complete head, intermediate section and flagella). In females, initiation of vitellogenesis was indicated by a change in the epithelial follicular morphology; in previtellogenesis it was stratified and polymorphic, while with the initiation of vitellogenesis it was reduced to constitute a cap of flattened cells. Given that in this stage, the follicular cells accumulate nutritious substances to form vitelline, it also was possible to observe vitelline plaques (grains of vitelo; Estrada & Uribe 2002).
Testicular, ovarian, and body mass data and whole testes diameter, follicle diameter, and SVL measurements were log10 transformed and the reproductive variables were regressed on the body size variables to yield size-adjusted residuals to permit inspection of evidence of reproductive cycles in these populations (Ramírez-B. & Vitt 1997). The residual values were compared using ANOVAs, with month and study site as main effects, using Tukey post-hoc tests when significant differences between months or sites were found.
Body size and mass of sexually immature individuals were compared among seasons using Mann-Whitney tests (sexes pooled). Body size and mass of adult females in the four reproductive categories (non-reproductive, vitellogenic, gravid with one egg, and gravid with two eggs) were compared using a Kruskall-Wallis test, with multiple median post-hoc tests when appropriate. Body size and mass of reproductive and non-reproductive adult males were compared using Mann-Whitney tests. When sample sizes permitted, X2 tests were used to inspect for heterogeneity in proportions of reproductive males and females in the two populations or in the dry vs. rainy seasons, using only individuals larger than the minimum size at maturity documented for each population using histological analyses.
Spearman correlations were used to inspect for a relationship between gravid female body and egg mass (Ramírez-B & Vitt 1997). Reproductive effort, defined as the amount of reproductive biomass relative to body mass, was calculated by dividing the mass of oviductal eggs by female body mass, and Mann-Whitney tests were used to inspect for differences in reproductive effort between seasons and study sites. Finally, mean daily precipitation values for the dry season months (December, January, and February) and rainy season months (October, November, and March) were compared using Mann-Whitney tests. Overall amounts of precipitation at the two study sites also were compared using the same test. Means are presented + 1 SD unless otherwise indicated. All statistical analyses were conducting with STATISTICA (Ver. 8.0; StatSoft, 2007) using a = 0.05 as the criteria for statistical significance.
RESULTS
Significantly more precipitation fell in the rainy season months (SP, X=5.90+6.3, n=92; SA, X=9.21+9.9, n=92) than in the dry season months at both San Pedro (X=2.03+3.5, n=90; Mann-Whitney U=2301, p<0.001) and San Antonio (X=3.95+6.5, n=90; Mann-Whitney U=2360, p<0.001). San Pedro (X=3.99+5.4, n=182) received significantly less precipitation than San Antonio (X=6.61+8.78, n=182) during the study period (Mann-Whitney U=13848, p=0.007; Fig. 1).
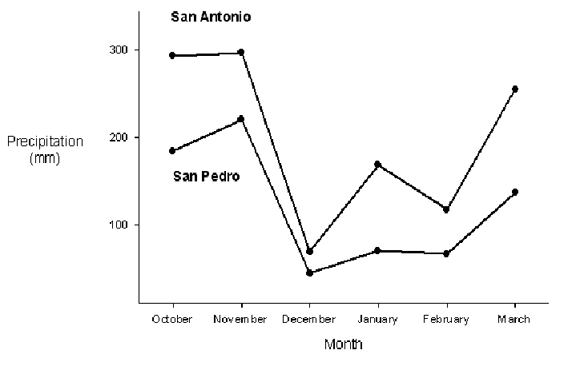
Figure 1. Mean daily precipitation from October 2008 to March 2009 at the San Pedro and San Antonio study sites.
In San Pedro, a total of 52 males and 63 females were collected, with no evidence of sexual body size dimorphism (SVL, Mann-Whitney U=1498, p=0.43). In San Antonio, 51 males and 57 females were collected, again with no significant sexual body size differences (SVL, Mann-Whitney U=1246.5, p>0.10). There also were no significant differences between sites in body size for either sex (males, SVL, Mann-Whitney U=1226.5, p>0.10; females SVL, Mann-Whitney U=1742.5, p>0.10).
Females in San Pedro with SVLs from 44.2 to 56.3 mm contained vitellogenic follicles or were gravid, and in San Antonio, females of 44.0 SVL or larger also were sexually mature, so the same minimum size at maturity (44 mm) was established for both populations in the analyses of reproductive seasonality (Fig. 2).
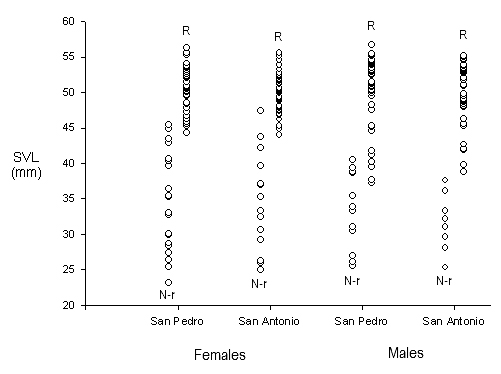
Figure 2. Reproductive status as a function of body size in females and males from San Pedro and San Antonio. N-r = Non-reproductive, R = Reproductive.
In San Pedro, males began spermatogenesis at a minimum of 34.8 mm SVL, with meiosis occurring in the spermatogonias but no spermatids observed in the seminiferous tubules at this body size. Sexually mature males with sperm in the lumen of the seminiferous tubules ranged from 37.1 to 56.7 mm SVL (Fig. 2). Thus, to inspect for seasonality of reproduction, only males of 37 mm SVL or greater were included in the analyses. In San Antonio, males began spermatogenesis at a minimum SVL of 32.8 mm and sexually mature males ranged from 38.7 to 55.1 mm SVL (Fig. 2), so only males of 39 mm SVL or greater were included in the analyses of reproductive seasonality.
There were no differences among immature individuals collected during the dry vs. rainy seasons in terms of body size (dry season, X=33.30+6.76, n=28; rainy season, X=34.08+5.08, n=26; Mann-Whitney U=334.0, p>0.10) or mass (dry season, X=0.98+0.62, n=28; rainy season, X=0.95+0.40, n=26; Mann-Whitney U=344.0, p>0.10). In sexually mature males, there was a significant positive relationship between SVL and testicular diameter (San Pedro, R2=0.83, F1,50=259.4, p<0.001; San Antonio, R2=0.84, F1,49=269.45, p<0.001) and between body mass and testicular mass (San Pedro, R2=0.89, F1,49=433.05, p<0.001; San Antonio, R2=0.66, F1,45=87.54, p<0.001). Absolute testes size did not vary among months (testicular diameter, San Pedro, F5,46=0.75, p>0.10; San Antonio, F5,45=1.96, p=0.10; testicular mass, San Pedro, F5,45=2.23, p>0.10; San Antonio, F5,41=2.22, p=0.07). Size-corrected two-way Anovas also failed to show differences among months, sites, or significant interactions in relative testes size or mass.
In San Pedro, 18 of the 63 females collected were immature. Of the sexually mature females, 13.3% were vitellogenic and 82.2% were gravid with one or two oviductal eggs (Fig. 3). Immature females never comprised more than 15% of the females examined in a given month. In San Antonio, 13 of the 57 females examined were immature, with 25.% of the mature females vitellogenic and 72.7% gravid (Fig. 3), again with immature females never constituting more than 15% of the total females examined in a month. There were no differences in the proportion of gravid females encountered in the dry and rainy seasons in San Pedro (X21=0.04; p>0.10) or in San Antonio (X21=1.37; p>0.10), nor were there differences in the proportion of females that were gravid in the two populations (X21=1.15; p>0.10). The three non-reproductive adult-sized females were all within 3 mm of the minimum size at reproductive maturity established for these populations and were encountered during the dry season months of January and February (Fig. 3).
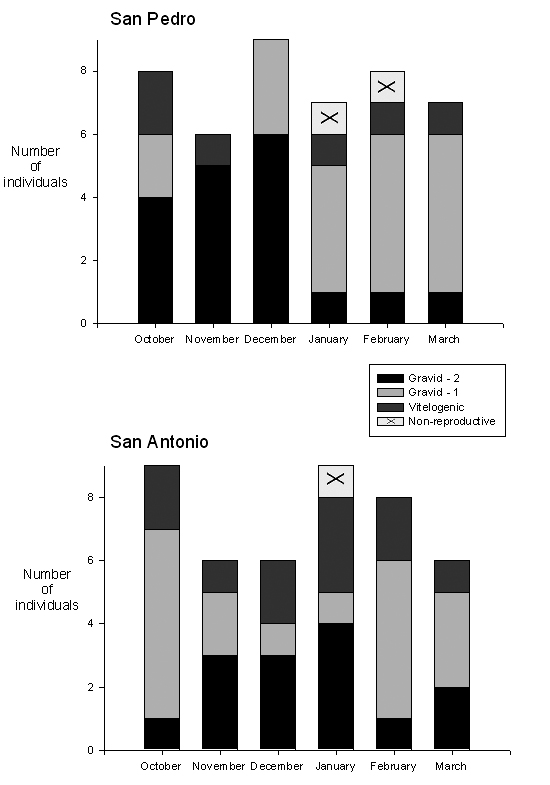
Figure 3. Reproductive status of adult females (SVL = 44 mm) by month at each site. Gravid-1 = females with 1 oviductual egg; Gravid-2 = females with 2 oviductual eggs.
In San Pedro, there was a significant positive relationship between female body size (SVL) and gonad size (diameter of the largest ovarian follicle (R2=0.81, F1,61=290.85, p<0.001) and gravid body mass was positively correlated to gonadal mass (R2=0.53, F1,44=50.42, p<0.001). In San Antonio, female body size and ovarian diameter also were positively correlated (R2=0.84, F1,54=250.31, p<0.001), but body and gonad mass were not (R2=0.02, F1,42=1.05, p>0.10). Size-corrected gonad size and mass did not vary between months at either site, nor were either site-month interaction significant (Gonad size, 2-way ANOVA, all p's >0.10, Gonad mass, 2-way ANOVA, all p's >0.10).
In females, gravid body weight was correlated with oviductal egg weight (San Pedro, Spearman R=0.33, p=0.04; San Antonio, Spearman R=0.45, p=0.009; Fig. 4). Reproductive effort (oviductal egg mass/gravid female body mass) was greater in San Pedro (San Pedro: 0.10+0.04, n=37; San Antonio:0.08+0.02, n=32; Mann-Whitney U=402, p=0.02), and was greater in the rainy season in San Pedro (rainy season, 0.12+0.03, n=20; dry season, 0.09+0.03, n=17; Mann-Whitney U=93, p=0.01), but not different between seasons in San Antonio (rainy season, 0.08+0.02, n=16; dry season, 0.09+0.3, n=16; Mann-Whitney U=116, p>0.10).
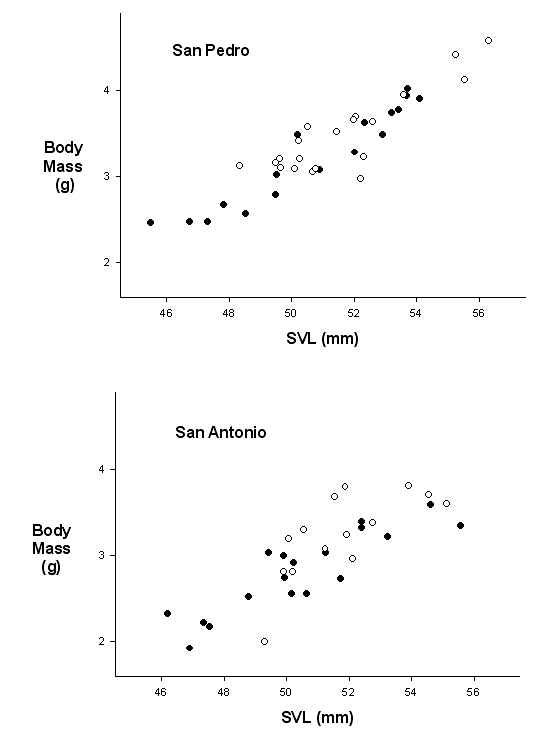
Figure 4. Correlation of SVL vs. gravid female body mass at the two study sites. Open circles are females carrying two oviductal eggs and shaded circles are females carrying only one oviductal egg.
Histological inspection of reproductive tracts corroborated the correct assignment of sex via gross anatomy in 104 cases (60 males and 44 females). Six immature males were incorrectly classified as females based on gross anatomy and one immature female was incorrectly classified as a male based on gross anatomy. On 11 occasions, males classified as being reproductively active based on the presence of convolutions in the epididymis were not actually producing sperm, while all males classified as possessing an unconvuluted epididymis were confirmed by histology to not be producing sperm.
DISCUSSION
Adult males in both populations were reproductive (producing viable sperm) during both the dry and rainy seasons, with comparable testicular sizes and masses in all months sampled. One problem with using macroscopic measures such as testicular mass to establish reproductive cycles is that differences might merely reflect the degree of hydration of the gonads (Vitt 1986) or variation in the intensity of sperm production (García-C et al. 1993). However, histological analyses also confirmed that males were reproductive with viable sperm during the entire study in both sites, despite the significant differences in the amounts of precipitation between months.
With adult females, there was also no evidence that the decrease in precipitation caused reproduction to cease at either site, with no differences detected in gonad size or mass during the two seasons, and with vitellogenic or gravid females abundant during all months. However, there was some suggestion that precipitation may have influenced reproductive rates. The only three non-reproductive females larger than the minimum size at first reproduction detected in this study were all encountered during the dry season (Fig 2). In addition, at the dryer site of San Pedro, there also was a smaller proportion of females carrying two eggs simultaneously in the dry season months, while females often carried two eggs in both the rainy and dry seasons at San Antonio. We assume the presence of females with two oviductal eggs indicates high rates of egg production during favorable periods, rather than a case of egg retention during harsh environmental conditions, as has been reported in other Anolis species (Huey, 1977), because only large females were shown to carry two eggs (Fig. 4). Small adult females just beginning to reproduce presumably are required to invest in both reproduction and growth, and thus would be expected to exhibit lower rates of egg production than larger females.
Finally, in pre-reproductive individuals, body size distributions were homogeneous during the different months of this study, further arguing that recruitment in these populations was continuous rather than pulsed. In addition, recently hatched individuals (less than 30 mm SVL) were captured in all months of the study. Thus, all of our evidence argues that reproduction is continuous in A. mariarum during the first dry season of the year. The second dry season in June to August is no longer or more severe than the dry season we studied, so we assume reproduction is continuous during that period as well. Ramirez-P et al. (1991) found reproduction to be continuous during both annual dry seasons for Anolis heterodermus in the climatic ecuator region, while Ardila-M et al. (2008) documented continuous reproduction during the first dry season of the year in Anolis tolimensis from the same region, and made the same assumption that we make here that it is reasonable to assume it is continuous in the second dry season each year as well.
The accumulating evidence for continuous breeding by Anolis species in the climatic equator region (Ramírez-P et al. 1991, Ardila-M et al. 2008; this study) suggests that Watling et al. (2005) were premature in arguing that total annual precipitation, but not length of the dry season, is what determines reproductive cycles in populations of mainland neotropical lizard species with fixed clutch sizes. When annual precipitation is limited, but occurs bimodally, individuals apparently are capable of sustaining reproductive investment throughout the relatively short pair of dry seasons each year.
However, mean annual temperature is a confounding factor in this analysis, given that 18 of 19 Anolis populations included in Watling et al. (2005) were from locations below 1200 m elevation, while all four populations that have been studied in the climatic equator region of Colombia to date were located above 2200 m elevation. It would be interesting to know whether reproduction is continuous or seasonal in Anolis species inhabiting dry lowland areas in the climatic equator area of the llanos of Colombia or Venezuela.
Our evidence that A. mariarum breeds during the dry season, but that some females may lower their reproductive rates at these times, especially at the dryer site, also illustrates the need to consider reproductive phenology as a continuum, rather than a dichotomy (i.e., not seasonal vs. continuous breeding). For example, in the meta-analysis of Watling et al. (2005), populations of Anolis limifrons from Barro Colorado Island (BCI) in Panama were classified as breeding seasonally. It is true that recruitment is limited during the dry season at this site (Andrews et al. 1983), but this primarily results from egg dessication (Andrews & Sexton 1981) and the fact that females that attain the minimum size for breeding during the dry season continue growing but refrain from reproducing until the rainy season begins (Andrews 1989). In contrast, adult females that have begun breeding before the onset of the dry season continue to oviposit during the dry season (Andrews & Rand 1974, Andrews 1979). Thus, this population would have to be considered intermediate to cases such as Anolis cupreus in Guanacaste, Costa Rica, where reproduction ceases completely during the six-month dry season (Fleming & Hooker 1975), and Anolis mariarum, where some, but not all, females may lower their reproductive rates or refrain from initiating reproduction during the two short dry seasons each year.
Just as reproductive phenology should not be treated as a dichotomy, analyses of the influence of rainfall on reproduction also should not consider the wet-rainy season classification as a simple dichotomy. Ideally, both the severity of the dry season and the pattern of precipitation during the dry season also should be considered. For example, while the mean annual precipitation of our San Antonio site is almost identical to the mean annual precipitation on BCI, much less precipitation occurs during the dry season months on BCI than during the two dry seasons at San Antonio, and it tends to occur in major downpours separated by many days with no precipitation, rather than in short showers occurring almost weekly, as in San Antonio. Reproduction in A. mariarum may be continuous not only because the two dry seasons are shorter than the single dry season that populations of Anolis experience in Central America, but also because they are less severe and have, on average, fewer dry days interspersed between dry season precipitation events.
In previous studies of A. mariarum populations (Bock et al. 2009, 2010), mean and maximum body sizes were shown to vary among populations as a function of mean annual precipitation levels at the sites. Bock et al. (2010) found no evidence of differences in adult survivorships between the wettest and driest sites, but speculated that perhaps body sizes were larger at the dryer site due to differences in juvenile survivorship rates and/or minimum sizes of sexual maturity there. Age-specific mortality theory predicts that body sizes should differ between populations in species with indetermiant growth when their juvenile mortality rates differ, with a life history shift towards delayed maturity, longer adult life spans, and larger body sizes in sites where juvenile mortality rates are higher (Abrams & Rowe 1996, Gadgil & Bossert 1970, Law 1979, Reznick 1982, Charnov & Berrigan 1990, Reznick et al. 1990). The results of this study were curious, because the differences in annual precipitation levels among the two study sites were comparable to those of the sites studied by Bock et al. (2010; 900 mm/year), yet we found no significant adult body size differences in either sex, with both populations exhibiting large body sizes (relative to the wet site studied by Bock et al. 2010). While it is difficult to quantify juvenile (egg and sub-adult) survivorship rates in Anolis lizards, our results clearly showed that females began reproducing at the same minimum body size (44 mm SVL) at both sites. Reciprocal transplant studies will be needed to determine whether the substantial microgeographic variation in mean and maximum body size in this species represents cases of local adaptation, or is due to phenotypic plasticity, but the generalization that individuals are larger at dryer sites apparently does not always hold true.
The mimimum body sizes at first reproduction we documented in A. mariarum were lower than those recently reported for A. tolimensis (43.7 mm SVL in males and 46.4 mm in females; Ardila-M et al. 2008), a closely related species (Poe 2004) also adapted to high elevation conditions in the Andes of Colombia. This difference could be partially due to our use of histology to confirm our determinations of sexual maturity based on inspection of gross reproductive anatomy, as the histological data set caused us to reduce our estimates of minimum body sizes at maturity for both sexes. Ardila-M et al. (2008) used only inspection of gross anatomy, as is typical of most studies of reproductive cycles in lizards. We suggest that histological analysis permits a more precise determination of sex in immature individuals and of minimum size at sexual maturity, especially for males, and hence will often justify the additional effort.
In sexually dimorphic species, the larger sex often begins to produce gametes later in life than the smaller sex (Stamps & Krishnan 1997), and although sexual dimorphism has been documented in some populations of A. mariarum (Bock et al. 2009), when it does occur it is always males that are the larger sex. Yet this study showed that it is females that mature at larger sizes, and because growth rates in the two sexes are comparable (Rubio-R 2009), presumable also at older ages. The sexual bimaturation exhibited in our populations may reflect selection for delayed maturation on the part of females due to the greater energy demands of producing eggs vs. sperm, or it may reflect selection for accelerated maturation on the part of males, or both. Sexual maturation in small males may seem curious, since it has traditionally been assumed in anoles that only the larger/older territorial males procure mating opportunities (Tokarz, 1995). But a physiological puberty that greatly preceeds the size/age of functional sexual maturity has been noted in other lizards, with speculations that early maturity may be more related to hormonal effects on male-male competition and early establishment of dominance relationships than to sperm production (Pratt et al. 1994). However, a recent study using molecular techniques documented that 21% of hatchlings in a population of A. carolinensis were sired by small males who surreptitiously resided within the territories of larger males (Passek, 2002).
ACKNOWLEDGEMENTS
This project was supported with a Sustainability Grant from the Comité para la Investigación CODI of the Universidad de Antioquia and a grant from IDEA WILD. We would like to thank E. Alzate for assistance with the field work and the Alzate-Otalvaro family for offering housing at the San Antonio de Prado site.
LITERATURE CITED
1. ABRAMS, P.A. & L. ROWE. 1996. The effects of predation on age and size of maturity of prey. Evolution 50: 1052-1061. [ Links ]
2. ANDREWS, R.M. 1979. Reproductive effort of female Anolis limifrons (Sauria: Iguanidae). Copeia 1979: 620-626. [ Links ]
3. ANDREWS, R.M. 1989. Intra-populational variation in the age of sexual maturity of the tropical lizard Anolis limifrons in Panama. Copeia 1989: 751-753. [ Links ]
4. ANDREWS, R.M. & A.S. RAND. 1974. Reproductive effort in anoline lizards. Ecology 55: 1317-1327. [ Links ]
5. ANDREWS, R.M., A.S. RAND & S. GUERRERO. 1983. Seasonal and spatial variation in the annual cycle of a tropical lizard. Pp. 441-454 in: A.G.J. Rhodin & K. Miyata (eds.), Advances in Herpetology and Evolutionary Biology. Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts. [ Links ]
6. ANDREWS, R.M. & O.J. SEXTON. 1981. Water relations of the eggs of Anolis auratus and Anolis limifrons. Ecology 62: 556-562. [ Links ]
7. ARDILA-MARÍN, D.A., D.G. GAITÁN-REYES, & E.J. HERNÁNDEZ-RUZ. 2008. Biología reproductiva de una población de Anolis tolimensis (Sauria: Iguanidae) en los Andes Colombianos. Caldasia 30: 151-159. [ Links ]
8. BOCK, B.C., A.M. ORTEGA, A.M. ZAPATA, & V.P. PAEZ. 2009. Microgeographic body size variation in a high elevation Andean anole (Anolis mariarum; Squamata, Polychrotidae). Revista de Biología Tropical 57: 1253-1262. [ Links ]
9. BOCK, B.C., A.M. ZAPATA, & V.P. PÁEZ. 2010. Survivorship rates of adult Anolis mariarum (Squamata: Polychrotidae) in two populations with differing mean and asymptotic body sizes. Papéis Avulsos de Zoologia 50: 43-50. [ Links ]
10. BORCHERT, R., S.S. RENNER, Z. CALLE, D. NAVARRETE, A. TYE, L. GAUTIER, R. SPICHIGER, & P. VON HILDEBRAND. 2005. Photoperiodic induction of synchronous flowering near the equator. Supplementary Discussion 2. Bimodal flowering and bimodal rainfall seasonality near the equator (p. 1-2). Nature 433: 627-629. [ Links ]
11. CHARNOV, E.L. & D. BERRIGAN. 1990. Dimensionless numbers and life history evolution: Age of maturity versus the adult life span. Evolutionary Ecology 4: 273-275. [ Links ]
12. ESTRADA, E. & M.C. URIBE. 2002. Atlas de Histología de Vertebados. Primera Edición. Universidad Nacional Autónoma de México, Editores y Impresores FOC, SA, de CV. México, D.F. [ Links ]
13. FLEMING, T.H. & R.S. HOOKER. 1975. Anolis cupreus: The response of a lizard to tropical seasonality. Ecology 56: 1243-1261. [ Links ]
14. GADGIL, M. & W.H. BOSSERT. 1970. Life historical consequences of natural selection. American Naturalist 104: 1-24. [ Links ]
15. GARCÍA-COLLAZO, R., T. ALTAMIRANO-ÁLVAREZ, & M. GÓMEZ-SOTO. 1993. Reproducción continua en Sceloporus variabilis variabilis (Sauria: Phrynosomatidae) en Alvarado, Veracruz, México. Boletín de la Sociedad Herpetológica de México 5: 51-59. [ Links ]
16. HUEY, R.B. 1977. Egg retention in some high-altitude Anolis lizards. Copeia 1977: 373-375. [ Links ]
17. LAW, R. 1979. Optimal life histories under age-specific predation. American Naturalist 114: 1-20. [ Links ]
18. MAGAÑA, V., J.A. AMADOR, & S. MEDINA. 1999. The midsummer drought over Mexico and Central America. Journal of Climate 12: 1577-1588. [ Links ]
19. PÁEZ, V.P., B.C. BOCK, J.J. ESTRADA, A.M. ORTEGA, J.M. DAZA, & P.D. GUTIÉRREZ-C. 2002. Guía de Campo de Algunas Especies de Anfibios y Reptiles de Antioquia. Multimpresos Ltda., Medellín. [ Links ]
20. PALACIO BAENA, J.A., E.M. MUÑOZ ESCOBAR, S.M. GALLO DELGADO, & M. RIVERA CORREA. 2006. Anfibios y Reptiles del Valle de Aburrá. Editorial Zuluaga Ltda., Medellín. [ Links ]
21. PASSEK, K.M. 2002. Extra-pair paternity within the female-defense polygyny of the lizard, Anolis carolinensis: Evidence of alternative mating strategies. Doctoral thesis, Virginia Polytechnic Institute and State University, Blacksburg, Virginia. [ Links ]
22. POE, S. 2004. Phylogeny of anoles. Herpetological Monographs 18: 37-89. [ Links ]
23. PRATT, N.C., J.A. PHILLIPS, A.C. ALBERTS, & K.S. BOLDA. 1994. Functional versus physiological puberty: An analysis of sexual bimaturation in the green iguana, Iguana iguana. Animal Behaviour 47:1101-1114. [ Links ]
24. RAMÍREZ-B, A. & L.J. VITT. 1997. Reproduction in the lizard Anolis nebulosus (Polychrotidae) from the Pacific coast of Mexico. Herpetologica 53: 423-431. [ Links ]
25. RAMÍREZ, M.P., J., G. DE PEREZ, P. RAMIREZ, & M.L. VARGAS. 1991. Ciclo ovárico de Phenacosaurus heterodermus (Sauria: Iguanidae) con relacion a niveles circulantes de lipoproteínas séricas y variación anual de las lluvias. Trianea 4: 513-526. [ Links ]
26. RAND, A.S. & W.M. RAND. 1982. Variation in rainfall on Barro Colorado Island. Pp. 47-59 in: E.G. Leight, Jr., A.S. Rand, & D.M. Windsor (eds.), The Ecology of a Tropical Forest: Seasonal Rhythms and Long-term changes. Smithsonian Institution Press, Washington, D.C. [ Links ]
27. REZNICK, D. 1982. The impact of predation on life history evolution in Trinidadian guppies: Genetic basis of observed life history patterns. Evolution 36: 1236-1250. [ Links ]
28. REZNICK, D.A., H. BRYGA, & J.A. ENDLER. 1990. Experimentally induced life-history evolution in a natural population. Nature 346: 357-359. [ Links ]
29. RUBIO-R, L.C. 2009. Ecología reproductiva y demografía de Anolis mariarum (Squamata: Polychrotidae) en dos localidades del departamento de Antioquia. Undergraduate thesis, Universidad del Valle, Cali. [ Links ]
30. SERRANO-C, V.H., M.P. RAMÍREZ-P, J.E. ORTEGA, & L.A. CORTES. 2007. Annual reproductive activity of Gonatodes albogularis (Squamata: Gekkonidae) living in an anthropic area in Santander, Colombia. South American Journal of Herpetology 2: 31-38. [ Links ]
31. SHINE, R. & A.E. GREER. 1991. Why are clutch sizes more variable in some species than in others? Evolution 45: 1696-1706. [ Links ]
32. SMALL, R.J.O., S.P. DE SZOEKE, & S.-P. XIE. 2007. The Central American midsummer drought: Regional aspects and large-scale forcing. Journal of Climate 20: 4853-4873. [ Links ]
33. STAMPS, J. & V.V. KRISHNAN. 1997. Sexual bimaturation and sexual size dimorphism in animals with asymptotic growth after maturity. Evolutionary Ecology, 11:21-39. [ Links ]
34. STATSOFT, INC. 2007. STATISTICA, data analysis software system, version 8.0. www.statsoft.com. [ Links ]
35. TOKARZ, R.R. 1995. Mate choice in lizards: A review. Herpetological Monographs, 8:17-40. [ Links ]
36. VITT, L.J. 1986. Reproductive tactics of sympatric gekkonid lizards with a comment on the evolutionary and ecological consequences of invariant clutch size. Copeia 1986: 773-786. [ Links ]
37. VITT, L.J. & J.P. CALDWELL. 2009. Herpetology. 3d Edition. Academic Press, Burlington, Massachusetts. [ Links ]
38. WATLING, J.I., J.H. WADDLE, D. KIZIRIAN, & M.A. DONNELLY. 2005. Reproductive phenology of three lizard species in Costa Rica, with comments on seasonal reproduction in neotropical lizards. Journal of Herpetology 39: 341-348. [ Links ]

