










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.33 no.1 Bogotá Jan./June 2011
Mangrove forests in a salinity gradient at Cispata bay - Boca Tinajones, department of Córdoba-Colombia
DENISSE VIVIANA CORTÉS-CASTILLO
J. ORLANDO RANGEL-CH.
Posgrado en Biología. Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C., Colombia. dvcortesc@unal.edu.co
Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C., Colombia. jorangelc@unal.edu.co, jorangelc@gmail.com
RESUMEN
Siguiendo un gradiente de salinidad desde la Bahía de Cispatá hasta Boca Tinajones en el departamento de Córdoba (Colombia), se caracterizó la vegetación según la composición florística y aspectos de la estructura de acuerdo con los lineamientos de la escuela fitosociológica de Braun-Blanquet. La riqueza florística se expresa en 40 especies, 38 géneros y 23 familias. La familia más importante fue Fabaceae con once especies. La vegetación se clasificó en tres (3) alianzas, una de ellas nueva: Fico dendrocidae-Rhizophorion manglis all.nov., y seis (6) asociaciones: Pelliciero rhizophorae-Rhizophoretum manglis ass.nov., Lagunculario racemosae-Conocarpodetum erecti Peinado et al. 1995, Lagunculario racemosae-Rhizophoretum manglis Peinado et al. 1994, Rhizophoretum manglis Cuatrecasas 1958, Priorio copaiferae-Rhizophoretum manglis ass.nov., y Annono glabrae-Ficetum dendrocidae ass.nov. La distribución diferencial de las especies indica que las variaciones de salinidad del agua y la estabilidad del suelo son los principales factores que condicionan el establecimiento y el desarrollo de la vegetación. En este sentido la vegetación de las alianzas Lagunculario racemosae-Rhizophorion manglis y Rhizophorion occidentalis se encuentra en lugares con alta salinidad y suelos estables, mientras que la de la alianza Fico dendrocidae-Rhizophorion manglis se establece en lugares con baja salinidad y suelos permanentemente inundados.
Palabras clave. Bosques de mangle, biodiversidad, Caribe colombiano, gradiente de salinidad, fitosociología.
ABSTRACT
Following the phytosociological school of Braun-Blanquet, the vegetation along a salinity gradient, from Cispatá Bay to Boca Tinajones at department of Córdoba (Colombia), was characterized. Floristic richness is represented by 40 species, 38 genera, and 23 families. The most important family was Fabaceae with eleven species. Vegetation was classified in three (3) alliances, one of them new: Fico dendrocidae-Rhizophorion manglis all.nov.; six (6) associations: Pelliciero rhizophorae-Rhizophoretum manglis ass.nov., Lagunculario racemosae-Conocarpodetum erecti Peinado et al. 1995, Lagunculario racemosae-Rhizophoretum manglis Peinado et al. 1994, Rhizophoretum manglis Cuatrecasas 1958, Priorio copaiferae-Rhizophoretum manglis ass.nov., and Annono glabrae-Ficetum dendrocidae ass.nov. The differential distribution of the species indicates that salinity of water and soil stability are the main factors influencing the establishment and development of vegetation. In this sense, vegetation of the Lagunculario racemosae-Rhizophorion manglis alliance is located in places with high salinity and stable soil, while Fico dendrocidae-Rhizophorion manglis and Rhizophorion occidentalis alliances are found in places with low salinity and soils permanently flooded.
Key words. Mangrove forests, biodiversity, Caribbean region, salinity gradient, phytosociology.
Recibido: 14/11/2010
Aceptado: 01/06/2011
Los manglares constituyen un complejo de ecosistemas boscosos, caracterizados por ubicarse en litorales tropicales y subtropicales (Walsh 1974) sobre sitios planos, suelos fangosos y aguas relativamente tranquilas en bahías, estuarios, lagunas costeras, ensenadas y esteros (Sánchez-Páez et al. 2000a). En la composición florística de las fitocenosis figuran varias especies de árboles denominados mangles que tienen diferentes tolerancias a la salinidad (Kathiresan 2005). Desde la costa, el manglar puede penetrar hacia el interior, siguiendo el curso de los ríos hasta donde las características del medio acuático, la consolidación del sustrato y la competencia con la vegetación de agua dulce se lo permiten. Se localizan en sectores donde las condiciones se caracterizan por temperaturas altas y constantes; normalmente están restringidos a latitudes entre los 30° norte y 30° sur (West 1956, Kathiresan 2005); aun así, existen formaciones de manglar en zonas que exceden estos límites latitudinales (Bermudas y Japón al norte; Nueva Zelanda, Australia y Sudáfrica al Sur), generalmente debido a factores ambientales locales como la corriente cálida del mar, la tensión de la salinidad y la acción del oleaje (Choudhury 1996).
En América, los manglares se localizan desde el Sur de Estados Unidos en el estado de Florida (29°53'Norte) hasta el Sur de Brasil (25°30'Sur) en la costa Atlántica y desde México (30°15'Norte) hasta Perú (en el estuario de Río Tumbes 3°40'Sur) en la costa Pacífica (Prahl 1990, Yáñez & Lara 1999, Sánchez-Páez et al. 2004). La superficie total de los manglares de América es de aproximadamente 4 133 216 hectáreas (41 332 km2), de las cuales América del Norte y Centro América, con las islas del Caribe suman 2 102 886 hectáreas y América del Sur cobija 2 030 330 hectáreas (Sánchez-Páez et al. 2004).
En Colombia, los manglares cubren aproximadamente 371 000 hectáreas, de las cuales 283 000 se localizan en la costa Pacífica y 88 000 en la costa Caribe (Sánchez-Páez et al. 2000b). El desarrollo de los manglares en el Caribe colombiano se da en estrechas franjas inundables (Prahl 1990) que se extienden desde el Golfo de Urabá (Antioquia) hasta Bahía Turkakas en la alta Guajira (Monroy-C. 2000). Los bosques de mejor desarrollo se presentan en el departamento de Córdoba, mientras que los del delta-exterior del río Magdalena (hacia la Isla de Salamanca y la Ciénaga Grande de Santa Marta) se consideran los de mayor extensión (Prahl 1990, Botero & Mancera 1996, Monroy-C. 2000). Además de las áreas continentales aparecen manglares en la Isla Fuerte, el Archipiélago de San Bernardo, la Isla de Tierra Bomba, las Islas del Rosario y en San Andrés y Providencia, estos últimos son los que cuentan con la menor cobertura, apenas 197.3 hectáreas (Yáñez & Lara 1999).
Entre los primeros estudios realizados en los manglares de Colombia incluyendo a los del Caribe, figuran los de Cuatrecasas (1958) en torno a la composición florística y a la descripción de rasgos ecológicos asociados a estos ambientes. Posteriormente los trabajos se enfocaron principalmente hacia la zonificación de los manglares del Caribe y al diseño de estrategias y lineamientos para el manejo sostenible y la conservación de estos ecosistemas (Sánchez-Páez et al. 1997, Sánchez-Páez et al. 2000a, Sánchez-Páez et al. 2000b). Trabajos en fenología, dinámica y regeneración de los manglares fueron realizados por Ulloa et al. (1998); estudios relacionados con el cambio en la estructura de la vegetación por efecto de la salinidad intersticial fueron realizados por Rivera-Monroy et al. (2006) para los manglares de la Ciénaga Grande de Santa Marta. Específicamente para el área de estudio, se destacan los aportes de Olaya-Hernández (1991) en donde se presenta una zonificación de los manglares de Bahía de Cispatá, así como el análisis del aprovechamiento de los recursos forestales y el de Sánchez-Páez et al. (2005), que consistió en el diseño de un plan de manejo integral de la zona de uso sostenible.
Los bosques de mangle del departamento de Córdoba son los terceros en extensión para la región Caribe de Colombia y comprenden los sectores de la antigua y nueva desembocadura del río Sinú en los municipios de San Antero y San Bernardo del Viento. El área del presente estudio se ubica entre la Bahía de Cispatá y la zona de Boca Tinajones y comprende los bosques de mangle y los bosques inundables (bosques estuarinos), asociados al gradiente de salinidad (desde la orilla del mar -sectores salobres- hacia el río -sectores de agua dulce-).
Desde el punto de vista geológico, el área de estudio se encuentra ubicada en un territorio de gran complejidad estructural, en donde convergen las placas de Suramérica y del Caribe, que en el Terciario produjeron las dos provincias estructurales más importantes de la zona: El cinturón del Sinú, localizado en la parte oeste de la zona costera y el cinturón de San Jacinto, que se encuentra hacia el este (Sánchez-Páez et al. 2005). Las unidades geomorfológicas identificadas en el área tienen diferente origen e incluyen geoformas fluviales, marinas y deltaicas. El perfil del suelo decrece desde la orilla del río o del mar hacia el interior de los bosques, donde se pueden formar leves depresiones con aguas estancadas (Olaya et al. 1991). Las condiciones físico-químicas del agua en caños y ciénagas varían en las épocas de lluvia, tornándose salobres durante el periodo seco.
El patrón de distribución de la precipitación, es de tipo unimodal-biestacional con un monto anual de 1337.4 mm y un periodo lluvioso que comprende desde mayo hasta noviembre y coincide con el aumento del caudal del río Sinú. El clima según la clasificación de Thornthwaite es semiseco con ligero o sin superávit de agua en la época de verano, los valores de evapotranspiración potencial del periodo seco indican una deficiencia de agua de 515.8 mm que se hace notoria en febrero (Rangel & Arellano 2010). La temperatura promedio anual es de 28°C, oscila entre 32.9°C en el mes de mayo y 26.7 °C en abril (Sánchez-Páez et al. 2005).
Los ensambles de vegetación que se encuentran son relativamente recientes, ya que la zona de estudio estuvo sujeta a los cambios de curso que experimentó el río Sinu en los últimos siglos. Palacios (2011) en su estudio paleoecológico, encontró que los manglares de la zona son de establecimiento reciente (menores a 2000 años). En la parte más antigua del sedimento de la bahía, la señal de polen que se encontró se asocia con varios tipos de vegetación actualmente frecuentes en las ciénagas del Caribe (Rangel et al. 2010). Los fenómenos de intrusión de las aguas marinas y la salinización de los suelos incidieron fuertemente en la composición y distribución actual de la vegetación y de la flora del lugar. Sumado a esto, estudios realizados en sistemas cenagosos del departamento y en áreas aledañas han mostrado que existe un grado considerable de similitud entre las biotas cordobesa y del Chocó biogeográfico, que hace pensar en Córdoba como un ecotono entre ambientes secos del Caribe y húmedos del Chocó (Rangel et al. 2010, Avella & Rangel, en imprenta).
En el presente estudio se caracterizó la vegetación a lo largo del gradiente de salinidad (desde la orilla del mar hacia la entrada del río Sinú) en la bahía de Cispatá en el departamento de Córdoba (Fig. 1) y se relacionaron de manera preliminar los cambios en las unidades de vegetación con condiciones de salinidad ya que todavía no se tienen suficientes datos en conductividad y salinidad intersticial para poder establecer de manera contundente su relación con los cambios en la vegetación.
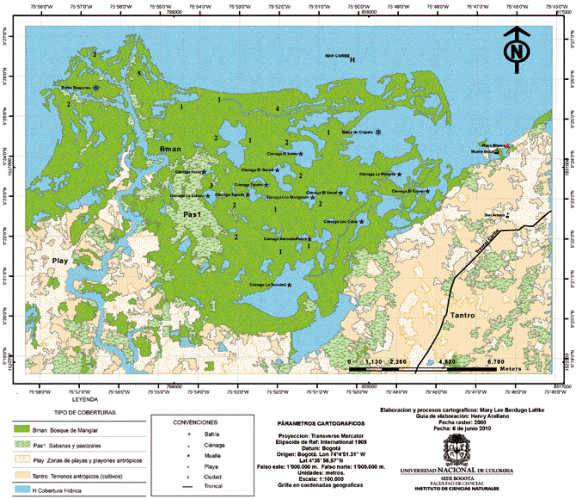
Figura 1. Mapa del área de estudio. Los números indican la cantidad de levantamientos realizados en cada sector. El color azul indica levantamientos en bosques estuarinos, el color fuchsia es para bosques de mangle.
METODOLOGÍA
La elección de los puntos de muestreo se realizó desde la orilla del mar (sectores salobres) hacia el río Sinú (sectores de agua dulce), con el objetivo de obtener un transecto que describiera el gradiente de salinidad. Una vez delimitados los puntos de muestreo, se realizaron levantamientos (parcelas) de 500 m2, área recomendada por Rangel & Velásquez (1997) para el trabajo en bosques de tierras bajas. En total se realizaron 28 levantamientos de 500 m2, 4 de 300 m2, 2 de 400 m2 y 1 de 200 m2, equivalentes a 1.62 ha entre los municipios de San Antero y San Bernardo del Viento. En cada levantamiento se elaboró un inventario de las especies presentes. Adicionalmente, se tomó información de localidad, coordenadas, altitud, fecha, numero de parcela (letra S), cobertura del dosel, porcentaje estimado de hojarasca en el suelo. Los atributos de campo que se evaluaron en la vegetación fueron diámetro a la altura del pecho (DAP), cobertura (m2), altura total (m). De todas las especies inventariadas se tomaron muestras botánicas bajo el código de colección DVC (Denisse Viviana Cortés) para su identificación en el Herbario Nacional de Colombia (COL), reconocido en el Index Herbariorum donde igualmente se depositaron.
Así mismo, se tomaron muestras de suelo en cada levantamiento, equivalentes a un kilogramo, con el fin de analizar las características físico-químicas básicas que fueron evaluadas en el laboratorio de suelos de la Facultad de Agronomía de la Universidad Nacional. Se realizaron mediciones de la conductividad en los cuerpos de agua aledaños a los puntos de muestreo al ser éste un parámetro indicativo de la salinidad con un Multiparámetro HI-98129. En la mayoría de los sectores, la toma de los datos de conductividad se hizo en dos épocas del año, ya que el régimen de lluvias puede afectar éste parámetro.
En las tablas de composición florística de las unidades de vegetación, los valores de cobertura (%) en cada levantamiento corresponden al estrato donde la especie presentó el valor máximo. Para el cálculo de la cobertura relativa (%), se siguieron los planteamientos de Rangel & Velásquez (1997). Los estratos se determinaron teniendo en cuenta a Rangel & Lozano (1986), así: Arbóreo inferior (Ai) 25-12 m, Arbolito o subarbóreo (Ar) 12-5 m, Arbustivo (ab) 5-1,5 m y Herbáceo (h) 1,5-0,25 m. Otro parámetro que se tuvo en cuenta fue el Índice de Valor de Importancia que se calculó usando la fórmula propuesta por Cottan & Curtis (1956) y Finol (1976) I.V.I = Densidad relativa (%) + Dominancia relativa (%) + Frecuencia relativa (%). Para mayor detalle del uso de éste índice en el Caribe colombiano se puede consultar a Rangel et al. (2009). Se estimó la riqueza de especies para cada una de las asociaciones o diversidad alfa, mediante la fórmula, Riqueza de especies de cada asociación= número de especies/unidad de área (m2); de esta manera, fue posible comparar la riqueza entre unidades que incluyeron inventarios de diferente área.
La definición de las unidades de vegetación se realizó con base en los planteamientos de la escuela fitosociológica de Braun-Blanquet (1979), teniendo en cuenta también algunas aportaciones de investigadores nacionales como Rangel et al. (2010). Se utilizó el método automatizado TWINSPAN con el programa PC-ORD para generar la matriz fitosociológica. Los nombres de las categorías fitosociológicas se designaron siguiendo el Código Internacional de Nomenclatura Fitosociologica (Weber et al. 2000, traducción oficial Izco & Del Arco, 2003).
RESULTADOS
Composición florística
La base de datos presenta un total de 3843 registros que corresponde a 35 levantamientos, éstos incluyen información sobre 23 familias, 38 géneros y 40 especies.
Sintaxonomía-arreglo fitosociológico
En un área muestreada de 16200 m2 (1.62 ha) que abarca localidades de dos municipios, la vegetación se clasifico en tres (3) alianzas y seis (6) asociaciones que se adscribieron a dos órdenes y una clase. Las formaciones vegetales establecidas para el sector de Bahía Cispatá y Boca Tinajones se pueden definir en dos grandes grupos:
-Bosques de mangle: vegetación de porte arbóreo con elementos que superan 12 m de altura; se desarrollan en sectores del litoral, ciénagas y bordeando caños de agua salada. Las especies dominantes son Rhizophora mangle, Avicennia germinans y Laguncularia racemosa. En este grupo también se incluyen a los bosques dominados por Conocarpus erectus y Pelliciera rhizophora.
-Bosques estuarinos: vegetación de porte arbóreo con elementos que no superan los 10 m de altura, se desarrollan sobre suelos inestables en zonas circundantes a las ciénagas sin entrada directa de agua salada. Las especies dominantes son Clathrotopis macrocarpa, Ficus dendrocida, Prioria copaifera y Montrichardia linifera.
El arreglo sintaxonómico tentativo es el siguiente:
Clases: Rhizophoro-Avicennietea germinantis Borhidi & Del Risco, in Borhidi 1991 (sensu Reyes & Acosta 2007) ó Rhizophoro manglis-Laguncularietea racemosae Peinado et al. 1995
Orden: Rhizophoro manglis-Laguncularietalia racemosae Peinado et al. 1995
Alianza Lagunculario racemosae-Rhizophorion manglis Peinado et al. 1995
Lagunculario racemosae-Rhizophoretum manglis Peinado et al. 1994
Pelliciero rhizophorae-Rhizophoretum manglis Cortés & Rangel ass. nov
Lagunculario racemosae-Conocarpodetum erecti Peinado et al. 1995; enmendó Cortés & Rangel en esta contribución
Orden Rhizophoretalia Cuatrecasas 1958
Alianza Rhizophorion occidentalis Cuatrecasas 1958 (sensu Reyes & Acosta 2007)
Rhizophoretum manglis Cuatrecasas 1958
Clase y orden no definidos
Alianza: Fico dendrocidae-Rhizophorion manglis Cortés & Rangel all. nov
Typus Priorio copaifera-Rhizophoretum manglis Cortés & Rangel ass. nov
Annono glabrae-Ficetum dendrocidae Cortés & Rangel ass. nov
Alianza Lagunculario racemosae-Rhizophorion manglis Peinado et al. 1995
Número de levantamientos: 26
Tabla 1
Composición florística. En la zona de estudio, la especie exclusiva de esta alianza es Pelliciera rhizophora; como especies selectivas se encuentran a Acrostichum aureum y Samanea saman; como especies acompañantes figuran Ficus bullenii, Terminalia catappa, Gliricidium sepium, Maclura tinctorea y Paullinia fuscencens. Solamente se encontró una especie epifítica, Tillandsia flexuosa. Entre las especies características-dominantes figuran Rhizophora mangle, Avicennia germinans y Laguncularia racemosa.
Fisionomía. En la alianza se reúne vegetación boscosa, conformada principalmente por manglar de borde. Las ramas superiores con hojas medianas se entrecruzan en el dosel generando una cobertura cercana al 70%; las ramas de altura media se mezclan con elementos trepadores, como Blepharodon mucronatum, Solanum jamaicense, Mesechites trifida y Sarcostema aff. glaucum. En estos bosques se diferencian cuatro estratos, herbáceo, arbustivo, de arbolitos y arbóreo inferior. Los valores más altos de cobertura se presentan en los estratos de arbolitos y arbóreo inferior, las especies dominantes son Rhizophora mangle (33 estrato arbolitos y 17% estrato arbóreo inferior), Laguncularia racemosa (10 y 9% respectivamente) y Avicennia germinans (9 y 8% respectivamente). El estrato arbustivo presenta una cobertura baja (6.1%) con dominio de R. mangle 3% y Conocarpus erectus 1%. En el estrato herbáceo domina el helecho Acrostichum aureum 1.4% de cobertura, aunque también se presentan otras especies como Eclipta prostrata, Wedellia calycina con coberturas inferiores al 2%.
Distribución y ecología. La vegetación se establece entre los 4 y los 10 m. El bosque se ubica principalmente en la zona de litoral, en sectores con alta influencia de las mareas o en terrenos circundantes a los cursos de agua, con suelos permanentemente encharcados. Los sectores que presentan esta vegetación son: Departamento de Córdoba: Municipio de San Antero (Ciénaga Navío, Ciénaga La Zona, Ciénaga Garzal, Caño Palermo, Ciénaga el Tábano, Caño Lobo), San Bernardo del Viento (Ciénaga Mireya, Caño Corea, Ciénaga Corea). Los suelos presentan un pH promedio de 4.8 que se interpreta como un suelo fuertemente ácido, el contenido de materia orgánica (CO%) fue de 14% y de Sodio (Na meq/100g) 124. La textura es Franco Limosa (FL), predomina el Limo con 56%, seguido de la Arena 24% y la Arcilla 20%. La conductividad registrada en estos sectores se encuentra entre los 20 µS/cm en la época de lluvias y hasta 36 200 µS en el periodo seco (tabla 6).
Pelliciero rhizophorae -Rhizophoretum manglis ass. nov.
Typus: S-24
Numero de levantamientos: cuatro (4)
Tabla 1
Composición florística. Se diferencia a Pelliciera rhizophora como la especie exclusiva de esta asociación. Las especies dominantes son Avicennia germinans y Rhizophora mangle y como acompañante se encuentra Acrostichum aureum.
Fisionomía. Bosque de ribera en el sector de Caño Salado, con elementos arbóreos de 13 m de altura y emergentes de 15 m; dosel no homogéneo, con copas altamente ramificadas. En el paisaje general, resaltan las raíces zanconas de algunos elementos (figura 2). El estrato arbóreo inferior presenta pocos individuos (36) y dominan en cobertura Rhizophora mangle 25% y Avicennia germinans 5%. El estrato de arbolitos tiene la mayor cobertura y se encuentra representado principalmente por individuos de Pelliciera rhizophora 18% y Laguncularia racemosa 1%. En el estrato herbáceo aparece Acrostichum aureum 1.5%, así como elementos de la regeneración de P. rhizophorae.

Figura 2. Aspectos de las asociaciones vegetales caracterizadas en la zona de estudio. Fotos: D. Cortés.
Lagunculario racemosae-Conocarpodetum erecti Peinado et al. 1995; enmendó Cortés & Rangel en esta contribución.
Numero de levantamientos: 5
Tabla 1
Composición florística. Especie característica de esta asociación es Conocarpus erectus cuya distribución se restringe a sectores de transición entre la playa y el interior del bosque. Entre las especies dominantes figura Laguncularia racemosa.
Fisionomía. Reúne vegetación boscosa y de matorral, en sectores de transición entre la playa y el bosque. La vegetación boscosa presenta un dosel discontinuo y elementos arbóreos que se ramifican a los pocos metros del suelo. El estrato de arbolitos, con un gran número de individuos (241) domina en cobertura y se encuentra representado por Laguncularia racemosa 10%, Avicennia germinans 9% e Hippomane mancinella 3%. Los estratos arbóreo inferior (con 6 individuos) y arbustivo (con 237 individuos) se encuentran representados por Conocarpus erectus 4% y Rhizophora mangle 1%.
Distribución y ecología. La vegetación de este tipo se encuentra en sectores estuarinos, de playa o bordeando ciénagas hacia la actual desembocadura del río Sinú; Departamento de Córdoba: Municipio de San Bernardo de Viento (Ciénaga Corea, Boca Mireya, Vereda Chiqui, Vereda Camino Real). Los sitios presentan signos evidentes de intervención antrópica, relacionados principalmente con la expansión de la frontera agrícola y la extracción de productos forestales (se destaca la entresaca de Conocarpus erectus). Los suelos presentan una textura franco-arenosa y son propensos a inundarse en la época de lluvias (de mayo a noviembre). La conductividad registrada para estos sectores es la más alta y oscila entre 18 200 y 36 200 µS/cm (tabla 6), ya que hacia la nueva desembocadura, la vegetación de este tipo se encuentra fuertemente influenciada por las mareas.
Sintaxonomía. Esta asociación originalmente adscrita a la alianza Lagunculario racemosae-Avicennion germinantis Peinado et al. 1995, se transfiere a la alianza Lagunculario racemosae-Rhizophorion manglis Peinado et al. 1994. Como se nota en la tabla 1 de Peinado et al. (1995), la asociación Lagunculario racemosae-Conocarpodetum erecti Peinado et al. 1995, enmendó Cortés & Rangel en esta publicación, se caracteriza por el dominio en cobertura de Conocarpus erectus, Laguncularia racemosa y valores bajos de Rhizophora mangle y Avicennia germinans. Aunque en nuestra tabla de vegetación, Rhizophora mangle y Avicennia germinans presentan valores altos de cobertura, contrario a las cifras que aparecen en la tabla 1 de Peinado et al. (1995), los valores exhibidos por L. racemosa justifican en nuestro concepto la transferencia.
Lagunculario racemosae-Rhizophoretum manglis Peinado et al. 1994.
Número de levantamientos: 17
Tabla 2
Composición florística. Esta asociación tiene como especies dominantes a Rhizophora mangle, Laguncularia racemosa y Avicennia germinans; especies acompañantes son Tabebuia rosea y Acrostichum aureum.
Fisionomía. Manglar de borde con 227 individuos en el estrato arbóreo inferior, en donde se destacan Rhizophora mangle 35% y Laguncularia racemosa con 30% de cobertura. En el estrato de arbolitos dominan Rhizophora mangle 60% y Avicennia germinans 30%; y en el estrato arbustivo Rhizophora mangle con 6.2% de cobertura. Acrostichum aureum fue la única especie registrada para el estrato herbáceo con 4% de cobertura.
Distribución y ecología. La vegetación de este tipo se encuentra representada en: Departamento de Córdoba- Municipio de San Antero (Ciénaga Navío, Ciénaga La Zona, Ciénaga Garzal, Caño Palermo, Ciénaga El Tábano, Caño Lobo) y San Bernardo (Caño soldado, Boca Mireya). Los suelos presentan un pH de 4.6, contenido de materia orgánica (CO%) 9.29 y de Sodio 3.68 (meq/100g). Tienen una textura Franca (F), predomina la Arena (46%), seguida por el Limo (42%) y la Arcilla (12%). La conductividad registrada tuvo un máximo de 35200 µS/cm en época seca y un mínimo de 0.23 en época de lluvias (tabla 6).
Sintaxonomía. Esta asociación guarda semejanza en su composición florística con la asociación Avicennio germinantis-Rhizophoretum manglis Reyes & Acosta 2007, manglares en áreas dispersas de costas cenagosas y en esteros con corriente fuerte. Sin embargo, en los manglares cubanos, no aparecen registros de Laguncularia racemosa, especie que por el contrario caracteriza a nuestra fitocenosis.
Alianza Rhizophorion occidentalis Cuatrecasas 1958.
Rhizophoretum manglis. Cuatrecasas 1958.
Número de levantamientos: 2
Tabla 3
Composición florística. La única especie que se presenta en esta asociación es Rhizophora mangle.
Fisionomía. Bosque de borde con dominancia absoluta de Rhizophora mangle, si bien esta asociación presenta los estratos arbóreo inferior, de arbolitos y arbustivo su fisionomía no es muy compleja, al tratarse de un bosque monoespecífico.
Distribución y ecología. La vegetación de este tipo se encuentra en: Departamento de Córdoba: Municipio de San Antero (Ciénaga Navío, Caño Salado). Suelos inestables, permanentemente inundados y con alta influencia de las mareas. La conductividad registrada para esta asociación va desde 2500 a 2.07 µS/cm (tabla 6).
Clase y orden no definidos
Alianza Fico dendrocidae-Rhizophorion manglis all. nov.
Typus: Priorio copaifera-Rhizophoretum manglis
Número de levantamientos: 7
Tabla 4
Composición. Especies exclusivas de la vegetación de esta alianza son Clathrotropis macrocarpa, Montrichardia linifera, Melothria pendula, Annona glabra, Ficus bullenii, Malmea anomala, Myrcia sp., Neea sp., Prioria copaifera, Casearia aff. aculeata, Erythrina fusca, Crescentia cujete y Cucumis melo. Especies preferentes son Mussatia hyacinthina y Ficus dendrocida; como especies acompañantes figuran Gliricidia sepium, Samanea saman, Inga aff. alba, Mesechites trífida, Dalbergia brownei, Tabebuia rosea y Solanum sect. micracantha. En la vegetación de la alianza, las especies son principalmente de ambientes semi-inundados y ciénagas, aunque también se presentan elementos halófitos de amplia valencia ecológica como Rhizophora mangle, Acrostichum aureum y el bejuco Paullinia fuscecens.
Fisionomía. Esta alianza recoge vegetación boscosa de formaciones de humedales (ciénagas) y zonas inundables; los troncos en la mayoría de los elementos arborescentes son de porte delgado y alcanzan los 12 m de altura. El estrato arbóreo inferior es de cubrimiento bajo aportado por Rhizophora mangle con 19% de cobertura; por el contrario, el estrato de arbolitos alcanza mayor vigor destacándose Ficus dendrocida 5% y Clathrotropis macroparca 1.4%. En el estrato arbustivo se presentan Montrichardia linifera 4.7% y Ficus dendrocida 3.5%. En el estrato herbáceo se encuentran elementos de la regeneración del bosque, así como helecho Acrostichum aureum en menor abundancia. Se incluye dentro de esta alianza a la asociación Annono glabrae-Ficetum dendrocidae que presenta una baja representación de Rhizophora mangle, Montrichardia linifera y Ficus dendrocidae. Estudios posteriores en bosques estuarinos deben permitir definir mejor las afinidades corológicas y sintaxonómicas de esta asociación.
Distribución y ecología. La vegetación de este tipo se encuentra representada en: Departamento de Córdoba: Municipio de San Bernardo del Viento (Ciénagas Coroza, Ostional, sector de El Guarumo). Se establece en los márgenes de las ciénagas y cursos de agua dulce (incluyendo caños taponados); los suelos presentan un pH de 4.3, contenido de materia orgánica (CO%) 18.6, probablemente relacionado con la alta cobertura de hojarasca y troncos; estos suelos presentan inundación en los periodos de lluvia. La conductividad mínima registrada fue de 0.15 µS/cm (tabla 6) en época de lluvias y máxima de 8310 µS en época seca.
Priorio copaiferae-Rhizophoretum manglis ass. nov.
Typus: S-23
Número de levantamientos: 4
Tabla 4
Composición. Especies exclusivas son Prioria copaifera, Crescentia cujete, Inga cf. alba; como especies acompañantes se encuentran a Ficus dendrocida, Montrichardia linifera, Dalbergia brownei, Solanum sp. sect. micracantha. Son especies preferentes Mussatia hyacinthina y Clathrotropis macrocarpa.
Fisionomía. Bosque ralo con individuos que alcanzan los 12 m de altura. En el estrato arbóreo inferior se encuentra a Rhizophora mangle con 35% de cobertura, en el estrato de arbolitos a Prioria copaifera 10%, Tabebuia rosea 3.7%, Ficus sp. 3%, Myrcia sp. 2.4%; en el estrato arbustivo Ficus dendrocida 1.6% y Montrichardia linifera 1.7% y en el herbáceo al helecho Acrostichum aureum con 11% de cobertura.
Distribución y ecología. La vegetación de este tipo se encuentra representada en: Departamento de Córdoba: Municipio San Bernardo del Viento (Ciénagas Ostional y Ciénaga La Coroza). El suelo tiene una textura Franco arenosa (FA), con predominio de Arena 62%, Limo 26% y Arcilla10%. La conductividad mínima registrada para esta asociación fue de 1550 µS/cm (tabla 6).
Annono glabrae-Ficetum dendrocidae ass. nov.
Typus: S-32
Número de levantamiento: 3
Tabla 4
Composición. Especies exclusivas de esta asociación son Erythrina fusca, Annona glabra, Malmea anómala y Casearia aff. aculeata. Como especies acompañantes se encuentran a Bactris guineensis, Clathrotropis macrocarpa, Ficus dendrocida y Montrichardia linifera.
Fisionomía. Bosquete con elementos arbóreos que no superan los 10 m de altura, los troncos son de gran porte en algunos individuos y además presentan estructuras espinosas en la corteza. La ramificación de los troncos se da a los pocos metros del suelo y el dosel es discontinuo, presentando una cobertura del 60%. El estrato arbóreo inferior se encuentra representado por un solo individuo de Erythrina fusca con cobertura de 0.4%, el estrato de arbolitos se encuentra dominado por Erythrina fusca 7.3%, Ficus dendrocida 6.9% y Annona glabra 3.7%. En el estrato arbustivo se encuentran Montrichardia linifera 8.8 % y Ficus dendrocida 5.8% de cobertura.
Distribución y ecología. La vegetación de este tipo está representada en: Departamento de Córdoba: Municipio San Bernardo del Viento (Ciénaga La Coroza y sector El Guarumo). Se establece sobre terreno con depresiones que puede sufrir proceso de inundación en la época de lluvias. La cobertura de hojarasca en el suelo es alta (70% aproximadamente). En ocasiones esta vegetación se observa al margen de caños cerrados o madres viejas de agua dulce únicamente. La conductividad máxima registrada para este tipo de vegetación fue de 1467 µS en época seca (tabla 6).
SINECOLOGÍA
Riqueza
El valor más alto en número de especies para los estratos arbóreo inferior y arbolitos se encontró en la alianza Fico dendrocidae-Rhizophorion manglis, doce especies en el estrato arbóreo inferior y cuatro en el de arbolitos, en los sitios con vegetación de esta alianza se registraron los valores más bajos en conductividad. Por otra parte, Lagunculario racemosae-Rhizophorion manglis presentó siete especies en el estrato arbóreo inferior y cinco en el estrato de arbolitos. A nivel de asociación, Priorio copaiferae-Rhizophoretum manglis tuvo el mayor número de especies (ocho y tres) mientras que Rhizohoretum manglis presentó el valor más bajo con una sola especie.
Estructura
La asociación Lagunculario racemosae-Rhizophoretum manglis mostró el valor más alto en número de individuos (1102), el más bajo se registró en la asociación Annono glabrae-Ficetum dendrocidae (71). En general, los valores por debajo del promedio (345) son predominantes. Los valores más altos en número de individuos para el estrato de arbolitos se encontraron en Lagunculario racemosae-Rhizohoretum manglis con 925. En la mayoría de los casos, la especie con el mayor número de individuos fue Rhizophora mangle (Cortés, 2011).
Índice de Valor de Importancia (IVI)
El Índice de Valor de Importancia (IVI) mostró a Rhizophora mangle como la especie más importante, alcanzando valores hasta de 300 en la asociación Rhizophoretum manglis. En la alianza Lagunculario racemosae-Rhizophorion manglis las especies con los valores más altos (tabla 5) fueron Rhizophora mangle (127.5), Laguncularia racemosa (68.9) y Avicennia germinans (66.3); la vegetación de la alianza Fico dendrocidae-Rhizophorion manglis mostró como especies importantes a Rhizophora mangle (86.7), Ficus dendrocidae (39.8), Prioria copaifera (33.5) y Erythina fusca (26.5).
Altura del dosel y Área basal (tabla 6)
De acuerdo con los datos recolectados, la altura promedio para los bosques de mangle fue de 8 m, con valores que oscilan entre 5 y 20 m; la ausencia de árboles en estratos superiores puede estar relacionada con factores como la inestabilidad del suelo, salinización del sustrato y también la extracción de recurso maderero. El área basal promedio fue de 6.3 m2 (oscilando entre 2.8 a 16 m2); los valores más altos fueron registrados en la asociación Lagunculario racemosae-Rhizophoretum manglis (16 m2), la cual, a pesar de encontrarse en un sector con alta salinidad presentó los bosques mejor desarrollados con elementos de gran porte.
DISCUSIÓN-CONSIDERACIONES FINALES
Composición florística
En el sector de la Bahía de Cispatá y Boca Tinajones, se registraron 23 familias, 38 géneros y 40 especies, distribuidas en dos formaciones vegetales que incluyen a los bosques de mangle y al bosque estuarino alrededor de las ciénagas. Cuando se cuenta únicamente a las especies arbóreas presentes en las parcelas, se pueden encontrar 15 familias, 20 géneros y 20 especies. En ambos casos, los valores de riqueza que se encontraron son mayores a los que se habían mencionado para el sector en estudios anteriores, en los cuales sólo se mencionaron nueve familias, once géneros y once especies, (Olaya-H et al. 1991; Sánchez-Páez et al. 2005). Esta diferencia puede estar relacionada con el menor área de estudio abarcada en los estudios mencionados anteriormente y con el nivel de detalle de los mismos.
Sintaxonomía
Las contribuciones de Delgadillo et al. (1992), Peinado et al. (1994, 1995), Reyes & Acosta (2003, 2007), se suman a las clásicas contribuciones de Cuatrecasas (1947, 1958a, 1958b) para ofrecer un marco detallado de la composición y ecología de los manglares en el continente americano, sin embargo la delimitación sintaxonómica de las diferentes unidades, en nuestro concepto no está clara y se presta a confusión en el proceso de tipificación. No es nuestra intención entrar a solucionar el asunto, razón por la cual en nuestro arreglo fitosociológico hemos incluido unidades muy antiguas como las mencionadas por Cuatrecasas (1947, 1958a, 1958b) con excelentes descripciones de la ecología pero carentes de tablas de vegetación tal como lo estipula el código de nomenclatura fitosociológica (Weber et al. 2000). Estas unidades sin embargo, fueron aceptadas por otros investigadores como Borhidi & del Risco (in Borhidi 1991), y Reyes & Acosta (2003, 2007). De las contribuciones de Cuatrecasas (1958a, 1958b) surgieron nombres como Rhizophorion occidentalis; Avicennion occidentalis que posiblemente fueron utilizados al interpretar las descripciones originales que se referían al distrito occidental en la distribución de las especies de mangles. Según Cuatrecasas (1958a, 1958b) todos los manglares del mundo constituirían "un gran orden de asociación": Rhizophoretalia y en jerarquías menores menciono al Rhizophorion para incluir las sinecias definidas por cada especie de Rhizophora, por ejemplo, Rhizophoretum manglis. Las unidades propuestas a partir de la década del 90 por los fitosociologos españoles que caracterizaron la vegetación halofita de Baja California siguen rigurosamente el código internacional de nomenclatura fitosociológica y en consecuencia hay nuevas unidades propuestas. Queda como una tarea para todos los interesados en la fitosociología de los manglares tratar de ordenar el tema.
Las asociaciones de mangles diferenciadas se lograron adscribir a dos órdenes Rhizophoretalia Cuatrecasas 1958 y a Rhizophoro mangle-Laguncularietalia racemosae Peinado et al. 1995, donde se incluye la alianza Lagunculario racemosae-Rhizophorion manglis Peinado et al. 1995 con las asociaciones Lagunculario racemosae-Rhizophoretum manglis Peinado et al. 1994, Pelliciero rhizophorae-Rhizophoretum manglis Cortés & Rangel ass. nov y la asociación Lagunculario racemosa-Conocarpodetum erecti Peinado et al. 1995 que se transfiere a esta alianza por las razones expuestas previamente.
Los bosques estuarinos de la alianza Fico dendrocidae-Rhizohorion manglis all. nov. cuyas relaciones sintaxonómicas a nivel de clase y orden no se lograron definir, engloban a dos asociaciones, Priorio copaiferae-Rhizophoretum manglis y Annono glabrae-Ficetum dendrocidae en las cuales es notoria la dominancia de Rhizophora mangle, especie que por su plasticidad ecológica es capaz de ocupar espacios con diferentes condiciones de salinidad, inundación y sustrato.
Corología-Sinecología
Cuando se compara la composición florística de la alianza Lagunculario racemosae-Rhizophorion manglis con otros sintaxones similares mencionados para la cuenca Caribe, se observa que los manglares de Córdoba son florísticamente similares a comunidades reseñadas para Guayana Inglesa, Surinam (Lyndenman 1953), Venezuela (Medina & Barboza 2003), Cuba (Reyes & Acosta 2007), México (Delgadillo et al. 1992, Peinado et al. 1994; 1995) y Puerto Rico; quizá una de las principales diferencias es la presencia de Pelliciera rhizophora en los manglares de Córdoba.
Comunidades vegetales con composición florística similar a la de las asociaciones Lagunculario racemosae-Rhizophoretum manglis y Rhizophoretum manglis han sido mencionadas en otras localidades del Caribe colombiano como en el Golfo de Urabá-Antioquia (Corpourabá 2003), Ciénaga Grande de Santa Marta (Álvarez-León et al. 2004), Tayrona (Bastidas & Corredor 1977), Ciénaga (Cleef 1984) y Chengue-Magdalena (Sánchez-Páez et al. 1997), Barranquitos-Bolívar (Sánchez-Páez et al. 1997), La Caimanera-Sucre (Sánchez-Páez et al. 1997), Caño Tijó-Córdoba (Sánchez-Páez et al. 1997).
Comunidades con Pelliciera rhizophora en el Caribe solamente han sido mencionadas para los departamentos de Antioquia (CORPOURABA 2003), Magdalena (Prahl 1990) y Córdoba (Olaya-H et al. 1991, Sánchez-Páez et al. 1997, Sánchez-Páez et al. 2005). Esta comunidad se encuentra mejor representada en la costa pacífica colombiana, donde está acompañada por Rhizophora mangle, Pterocarpus officinalis y Mora megistosperma (Rangel 2004).
Bosques con predominio de Conocarpus erectus fueron mencionados para Antioquia (Corpourabá 2003) y Córdoba (Sánchez-Páez et al. 1997), aunque en los departamentos de Atlántico y La Guajira también hay registros de esta especie pero en densidades muy bajas.
En la Costa Pacífica colombiana, Rangel (2004) también señaló la presencia de manglares con dominancia de Rhizophora mangle, Avicennia germinans, Laguncularia racemosa y Pelliciera rhizophora.
No se conocen registros de asociaciones como Priorio copaiferae-Rhizophoretum manglis y Annono glabrae-Ficetum dendrocidae. Zuluaga (1987) reseñó la presencia de formaciones dominadas por Prioria copaifera que denominó Prioretum copaiferae, las cuales forman extensas selvas con elementos arborescentes de 30 m de altura en el área del P.N.N. Katios. En el departamento de Córdoba, Rangel et al. (2010) mencionaron la presencia de individuos de P. copaifera e igualmente describieron bosques en áreas no inundables alrededor de los complejos cenagosos, con dominancia de Annona punicifolia, Pithecellobium lanceolatum y Ficus dendrocida. En el departamento del Magdalena, Álvarez-León et al. (2004) caracterizaron bosques mixtos de ambientes pantanosos en la Ciénaga Grande de Santa Marta, cuya composición florística a nivel de género es similar a la reseñada para la alianza Fico dendrocidae-Rhizophoretum manglis. Estos resultados permiten considerar a la asociación Annono glabrae-Ficetum dendrocidae como una comunidad de transición entre los bosques altamente influenciados por las ciénagas (inclusive con influencia marina) y los bosques no inundables.
En general, la zonación obtenida para los bosques de mangle y estuarinos en la bahía de Cispatá, está relacionada con los ciclos de inundación por mareas o por crecientes de los cauces de la siguiente manera: hacia las zonas de litoral con influencia directa de las mareas, dominó R. mangle, que se puede anclar a suelos inestables por medio de sus raíces zanconas; después de ésta primera franja se encuentran A. germinans con L. racemosa, en suelos más estables. Hacia los sectores de la playa, lejos del embate directo de las olas se encuentra Conocarpus erectus. En sectores aledaños a canales de agua salobres se establece Pelliciera rhizophora. Siguiendo los canales y ciénagas en dirección al río, la composición florística cambia para dar lugar a los bosques inundables que se distribuyen como parches de vegetación rodeados de ciénagas de agua dulce o con baja influencia de las mareas, donde se distinguen Ficus dendrocida, Clathrotropis macrocarpa, Pioria copaifera, Neea sp. (DVC 306) y Myrcia sp. (DVC 253) entre otras. Estos bosques que se pueden considerar como transicionales, presentan una fisionomía achaparrada y un dosel discontinuo posiblemente relacionado con el terreno inestable donde se encuentran.
AGRADECIMIENTOS
A la Vicerrectoría de Investigaciones de la Universidad Nacional y a la División de Investigación de la sede Bogotá-DIB por el apoyo al proyecto: Valoración de la biodiversidad del Caribe colombiano: síntesis del conocimiento y servicios ambientales (captura de CO2). A la comunidad de San Bernardo del Viento, especialmente al señor Jesús Espitia y familia por su colaboración y hospitalidad. A los doctores Heliodoro Sánchez (Universidad Distrital Francisco José de Caldas, Colombia), Joel O. Reyes (Cuba) y otros dos revisores que con sus sugerencias y recomendaciones ayudaron a mejorar la presentación de este manuscrito.
LITERATURA CITADA
1. ÁLVAREZ-L., R., E. CARBONÓ-D., W. TRONCOSO-O., O. CASAS-M. & P. REYES-F. 2004. La vegetación terrestre, eurihalina y dulceacuícula de la ecoregión Ciénaga Grande de Santa Marta. En: J. Garay, J. Restrepo, O. Casas, O. Solano & F. Newmark (eds). Los manglares de la Ecoregión Ciénaga Grande de Santa Marta: pasado, presente y futuro. INVEMAR - Serie de publicaciones especiales N°11: 77-96. Santa Marta. [ Links ]
2. BASTIDAS, N. & H. CORREDOR. 1977. Contribución al estudio Fitosociológico del Parque Nacional Natural Tayrona (ensenadas de Chengue y parte este de Nenguange). Tesis de biólogos. Universidad Nacional de Colombia. 75 pp. (Mimeografiada). [ Links ]
3. BORHIDI, A. 1991. Phytogeography and vegetation ecology of Cuba. Akadémiai Kiado, Budapest, 857 pp. [ Links ]
4. BOTERO, L. & J.E. MANCERA. 1996. Síntesis de los cambios de origen antropico ocurridos en los últimos 40 años en la Cienaga Grande de Santa Marta. Rev. Acad. Colomb. Cienc. 20 (78): 465-474. [ Links ]
5. BRAUN-BLANQUET, J. 1979. Fitosociología: bases para el estudio de las comunidades vegetales. H. Blume Ediciones, Madrid. [ Links ]
6. CHOUDHURY, J. K. 1996. Mangrove Forest Management. Mangrove Rehabilitation and Management Project in Sulawesi. 297 pp. [ Links ]
7. CLEEF, A.M. 1984. Synopsis of the coastal vegetation of the Santa Marta area: 423-440. In: T. van der Hammen & P.M. Ruíz. (eds). La Sierra Nevada de Santa Marta (Colombia), transecto Buritaca - La Cumbre. Studies on Tropical Andean Ecosystems 2. J. Cramer, Berlin - Stuttgart. [ Links ]
8. CORPOURABA. 2003. Caracterización y zonificación de los manglares del Golfo de Urabá, Departamento de Antioquia. Proyecto zonificación y ordenación de los manglares de Urabá, Antioquia. Corpouraba. Ministerio de Medio Ambiente. Apartadó. [ Links ]
9. CORTÉS-S., D.V. 2011. Los bosques de mangle en un gradiente de salinidad en la bahía de cispatá, departamento de Córdoba-Colombia. Tesis de Maestría. Instituto de Ciencias Naturales, Facultad de Ciencias. Universidad Nacional de Colombia. [ Links ]
10. COTTAM, G. & J.T. CURTIS. 1956. The use of distance measures in phytosociological sampling. Ecology 37(2): 451-460. [ Links ]
11. CUATRECASAS, J. 1947. Vistazo a la vegetación natural del Bajo Calima. Rev. Ac. Col. Cienc. Ex. Fis & Nat. 7: 306-312. [ Links ]
12. CUATRECASAS, J. 1958a. Introducción al estudio de los manglares. Bol. Soc. Bot. México 23: 84-98. [ Links ]
13. CUATRECASAS, J. 1958b. Aspectos de la vegetación natural de Colombia. Rev. Ac. Col. Cienc. Ex. Fis & Nat. 10(40): 221-268. [ Links ]
14. DELGADILLO. J, M. PEINADO, J. MARTÍNEZ, F. ALCARAZ & A. DE LA TORRE. 1992. Análisis fitosociológico de los saladares y manglares de Baja California, México. Acta Botánica Mexicana 19: 1-35. México. [ Links ]
15. IZCO, J. & M. DEL ARCO. 2003. (Traducción oficial). Código Internacional de Nomenclatura Fitosociológica. Universidad de la Laguna. 155 pp. España. [ Links ]
16. KATHIRESAN, K. 2005. Mangrove Ecosystems: Distribution of mangroves. En: K. Kathiresan & S.Z. Quasim (eds). Biodiversity of mangroves: 92-101. Hindustan publications. [ Links ]
17. LINDEMAN, J. C. 1953. The vegetation of the coastal region of Suriname. Drukkerij en Uitg.-Mij. v/h Kemink. 137p. Thesis of Ph.d. Utrecht. [ Links ]
18. MEDINA, E. & F. BARBOZA. 2003. Manglares del sistema del lago de Maracaibo: Caracterización fisiográfica y ecológica. Ecotrópicos 16(2):75-82. [ Links ]
19. MONROY-C., O. 2000. Estado de los manglares en Colombia. En: Informe del estado de los ambientes marinos y costeros en Colombia. INVEMAR. pp. 48-69 [ Links ]
20. OLAYA-H., H., C. MARTÍNEZ, D. LEGUIZAMO & F. PINEDA. 1991. Los bosques de mangle del antiguo delta del río Sinú (Córdoba-Colombia). Estudio de la vegetación y el aprovechamiento. Corporación Autónoma Regional de los Valles del Sinú y del San Jorge, 30 pp. Montería (Informe interno). [ Links ]
21. PALACIOS, L. 2011. Cambios paleoambientales en el sistema estuárico bahía de Cispatá (Caribe colombiano) durante el Holoceno tardíoTesis de Magister. Universidad Nacional de Colombia-Instituto de Ciencias Naturales. Bogotá, D.C. [ Links ]
22. PEINADO, M., A. HENARES, F. ALCARAZ & J. DELGADILLO. 1995. Syntaxonomy of some halophilous communities of North and Central America. Phytocoenologia 25(1): 23-31. [ Links ]
23. PEINADO, M., F. ALCARAZ, J. DELGADILLO, M. DE LA CRUZ, J. ÁLVAREZ & J.L. AGUIRRE. 1994. The coastal salt marshes of California and Baja California. Vegetatio 110: 55-66. [ Links ]
24. PRAHL, H. C. 1990. Manglares. Villegas Editores, 200 pp. Bogotá, D.C [ Links ]
25. RANGEL-CH., J.O. & G. LOZANO. 1986. Un perfil de vegetación entre La Plata (Huila) y el volcán de Puracé. Caldasia 14 (68-70): 503-547. [ Links ]
26. RANGEL-CH., J.O. & A. VELÁSQUEZ. 1997. Métodos de estudios de la vegetación. En: J.O. Rangel-Ch. (ed). Colombia Diversidad Biótica II. Tipos de vegetación en Colombia: 59-87. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
27. RANGEL-Ch., J.O. 2004. La vegetación del Chocó biogeográfico de Colombia y zonas cordilleranas aledañas-Síntesis. En: J.O. Rangel-Ch. (ed.). Colombia Diversidad Biótica IV. El Chocó Biogeográfico-Costa Pacífica: 769-815. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
28. RANGEL-CH., J.O., A. AVELLA-M. &. H. GARAY-P. 2009. Caracterización florística y estructural de los relictos boscosos del sur del departamento del Cesar. En: J.O. Rangel-Ch (ed.) Colombia Diversidad Biótica VIII. Media y baja montaña de la Serranía del Perijá: 365-392. Instituto de Ciencias Naturales. Universidad Nacional de Colombia-Corpocesar. Bogotá, D.C. [ Links ]
29. RANGEL-CH., J.O. & H. ARELLANO. 2010. Clima. En: J.O. Rangel-Ch. (ed.) Colombia Diversidad Biótica IX. Ciénagas de Córdoba: Biodiversidad, ecología y manejo ambiental: 1-14. Instituto de Ciencias Naturales. Universidad Nacional de Colombia-CVS. Bogotá, D.C. [ Links ]
30. RANGEL-CH., J.O., H. GARAY-P & A. AVELLA-M. 2010. Bosques húmedos y secos circundantes a los complejos de humedales (ciénagas), en el departamento de Córdoba. En: J.O. Rangel-Ch (ed.) Colombia Diversidad Biótica IX. Ciénagas de Córdoba: Biodiversidad, ecología y manejo ambiental: 207-323. Instituto de Ciencias Naturales. Universidad Nacional de Colombia-CVS. Bogotá, D.C. [ Links ]
31. REYES, O.J. & F. ACOSTA. 2003. Fitocenosis presentes en las áreas costeras del Sur de la Sierra Maestra, Cuba I. Comunidades con influencia marina. Foresta Veracruzana 5(2): 1-7. [ Links ]
32. REYES, O.J. & F. ACOSTA. 2007. Principales fitocenosis en el humedal del delta del río Cauto, Cuba oriental. II. Herbazal halófito y manglares. Foresta Veracruzana 9(2): 1-7. [ Links ]
33. RIVERA-DÍAZ, O. 2010. Flora. En: J.O. Rangel-Ch. (ed.) Colombia Diversidad Biótica IX. Ciénagas de Córdoba: Biodiversidad, ecología y manejo ambiental: 121-136. Instituto de Ciencias Naturales. Universidad Nacional de Colombia-CVS. Bogotá D.C. [ Links ]
34. RIVERA-MONROY, V., R. TWILLEY, E. MANCERA, A. ALCANTARA-E., E. CASTAÑEDA-M., O. CASAS-M., P. REYES, J. RESTREPO, L. PERDOMO, E. CAMPOS, G. COTES & E. VILORIA. 2006. Aventuras y desventuras en Macondo: rehabilitación de la Ciénaga Grande de Santa Marta, Colombia. Ecotropicos 19(2): 72-93. [ Links ]
35. SÁNCHEZ-P., H., R. ÁLVAREZ-L., F. PINTO-N., A. SÁNCHEZ-A., J.C. RENJIFO, I. GARCÍA-H. & M.T. ACOSTA-P. 1997. Diagnóstico y zonificación preliminar de los manglares del Caribe Colombiano. Ministerio del Medio Ambiente, OIMT. 511 pp. Bogotá D.C. [ Links ]
36. SÁNCHEZ-P., H., G. ULLOA-D. & R. ÁLVAREZ-L. 2000a. Lineamientos estratégicos para la conservación y uso sostenible de los manglares de Colombia. Ministerio del Medio Ambiente, ACOFORE, OIMT. 290 pp. Bogotá. D.C. [ Links ]
37. SÁNCHEZ-P., H., G. ULLOA-D. & R. ÁLVAREZ-L. 2000b. Hacia la recuperación de los manglares del Caribe de Colombia. Ministerio del Medio Ambiente, ACOFORE, OIMT. 290 pp. Bogotá D.C. [ Links ]
38. SÁNCHEZ-P., H., G. ULLOA-D. & H. TAVERA-E. 2004. Manejo integral de manglares por comunidades locales. Caribe de Colombia. Ministerio del Medio Ambiente, CONIF, OIMT. 335 pp. Bogotá D.C. [ Links ]
39. SÁNCHEZ-P., H., G. ULLOA-D., H. TAVERA-E & W. GIL-T. 2005. Plan de manejo integral de los manglares de la zona de usos sostenible del sector estuarino de la Bahía de Cispatá. Departamento de Córdoba-Colombia. Corporación Autónoma Regional de los Valles del Sinú y del San Jorge (CVS), Corporación Nacional de Investigaciones y fomento forestal, OIMT. 202 pp. Bogotá D.C. [ Links ]
40. ULLOA, G., H. SÁNCHEZ-P., W. GIL-T., J.C. RENJIFO, H. RODRÍGUEZ & R. ÁLVAREZ-L. 1998. Conservación y Uso sostenible de los manglares del Caribe colombiano. Ministerio del Medio Ambiente, ACOFORE, OIMT. 224 pp. Bogotá D.C. [ Links ]
41. WALSH, G. E. 1974 Mangroves: A review. In: R.J. Reinhold & W.H. Queen (eds). Ecology of Halophytes: 51-174. Academic Press. Nueva York. [ Links ]
42. WEBER, H.E., J. MORAVEC & J.P. THEURILLAT. 2000. International code of phytosociological nomenclature. 3rd edition. Journal of Vegetation Science 11: 739-768. [ Links ]
43. WEST, R.C. 1956. Mangrove Swamps of the Pacific Coast of Colombia. Annals of the Association of American Geographers 46(1): 98-121. [ Links ]
44. YÁÑEZ-ARANCIBIA, A. & A. L. LARA-DOMÍNGUEZ. 1999. Los manglares de América Latina en la encrucijada, p. 9-16. En: A. Yáñez-Arancibia y A. L. Lara-Domínquez (eds). Ecosistemas de Manglar en América Tropical. Instituto de Ecología A.C. México, UICN/ORMA, Costa Rica, NOAA/NMFS Silver Spring MD 380 p. USA. [ Links ]
45. ZULUAGA, S., 1987. Observaciones fitoecológicas en el Darién colombiano. Pérez-Arbelaezia 1 (4-5): 85-145. [ Links ]

