










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.33 no.2 Bogotá Jul./Dec. 2011
Palm communities in two forests of Chocó, Colombia
GIOVANNY RAMÍREZ-MORENO
GLORIA GALEANO
Instituto de Investigaciones Ambientales del Pacífico (IIAP), carrera 6 No. 27-39, Barrio Huapango, Quibdó, Chocó, Colombia. gramirez@iiap.org.co
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, D. C., Colombia. gagaleanog@unal.edu.co
RESUMEN
Estudiamos la riqueza, la abundancia, la composición y la estructura de las comunidades de palmas en dos sitios de la selva central del Chocó, Colombia. Se establecieron 14 transectos (1.5 ha) en Tutunendo y 12 transectos (1 ha) en Angostura, y en cada uno se registraron e identificaron las palmas, incluyendo todos los tamaños (plántulas, juveniles y adultos); además, se indagó sobre nombres comunes y usos de cada especie, entrevistando a 16 hombres mayores de 45 años y conocedores del bosque. La riqueza de palmas encontrada en las localidades estudiadas (29 especies en 1.5 ha y 23 especies en 1 ha), está entre las más altas registradas para el Neotrópico, y corrobora el patrón de distribución de riqueza ligado a altas precipitaciones. La mayoría de las especies están representadas por pocos individuos, mientras que unas pocas son las más abundantes y frecuentes. Se identificó un grupo de especies (Wettinia quinaria, Welfia regia, Iriartea deltoidea, Socratea exorrhiza, Oenocarpus bataua, en el dosel, y Geonoma cuneata, en el sotobosque), que son particularmente importantes a nivel ecológico, y que caracterizan y modelan la vegetación, no sólo del área de estudio, sino también, al parecer, a lo largo de la región del Chocó Biogeográfico. Con base en los resultados y en otros datos florísticos, se plantea que, si bien las palmas son importantes a todo lo largo de la región, la mayor abundancia y riqueza parece presentarse hacia la zona central, que coincide con un aumento en el gradiente de precipitación. Veintisiete de las 33 especies encontradas tenía algún uso en una de las siete categorías de uso, lo cual muestra la importancia de las palmas para las poblaciones locales.Palabras clave. Estructura, Chocó biogeográfico, composición, diversidad, palmas.
ABSTRACT
We studied the richness, abundance, composition and structure of the palm communities in two localities of the central Chocó, Colombia. We made 14 transects (1.5 ha) in Tutunendo and 12 transects (1 ha) in Angostura. In each transect we counted and identified all palm individuals, including all size classes (seedlings, juveniles, and adults); additionally, local names and uses for the species were documented through interviews with 16 knowledgeable men, older than 45 years. Palm richness (29 species in 1.5 ha and 23 species in 1 ha) is among the highest recorded in the Neotropics and supports a richness pattern associated with high rainfall. Most species were represented by a few individuals while a few species were highly abundant and dominant. A group of species (Wettinia quinaria, Welfia regia, Iriartea deltoidea, Socratea exorrhiza, Oenocarpus bataua, in the canopy, and Geonoma cuneata in the understory) is particularly ecologically important because it characterizes and shapes the vegetation at the study sites and probably also throughout the Biogeographic Chocó region. Based on these results and on other floristic data, we suggest that although palms are important at all levels throughout the Biogeographic Chocó region, their greatest abundance and richness seem to occur towards the central zone, which corresponds with an increase in the rainfall gradient. Twenty-seven of the 33 species had some use in one of seven use categories, which stresses the importance of palms to local people.
Key words. Chocó Biogeographic region, composition, diversity, palms, richness, structure.
Recibido: 29/10/2010
Aceptado: 04/10/2011
INTRODUCCIÓN
Las palmas son uno de los grupos de plantas más importantes a nivel neotropical, no sólo por su riqueza, abundancia y papel fundamental en los ecosistemas en donde crecen, sino también por su uso actual y potencial por parte de las comunidades humanas (Henderson et al. 1995). Por esta razón, uno de los temas de investigación más activo actualmente está relacionado con los patrones de distribución y abundancia de esta riqueza en las diferentes escalas, cuyo entendimiento puede aportar herramientas para la conservación y el manejo de este recurso. A nivel del Neotrópico, en los últimos años se han realizado varios análisis tratando de entender este fenómeno (Clark et al. 1995, Bjorholm et al. 2005, 2006, 2008, Normand et al. 2006, Svenning et al. 2008, Balslev et al. 2011, Kristiansen et al. 2011, entre otros), y en algunos de ellos (Bjorholm et al. 2005, 2006) se señala el noroccidente de Colombia como una de las áreas donde se presenta mayor riqueza de palmas. En efecto, Colombia es uno de los países más ricos en palmas a nivel mundial y Neotropical, y gran parte de esta riqueza se encuentra concentrada en las tierras bajas del Pacífico o Chocó Biogeográfico (Galeano & Bernal 2010). En estudios generales sobre los bosques de esta región (Rangel 2004, Gentry 1986, Faber-Langendoen & Gentry 1991, Galeano et al. 1998a,b, García et al. 2002, entre otros), siempre se destaca la importancia de las palmas, tanto por su riqueza como por su abundancia. Sin embargo, para la región no hay estudios específicos sobre la estructura, riqueza, abundancia y composición en comunidades de palmas que incluyan todas las formas de vida y los estados de desarrollo de los individuos, de tal suerte que permitan una visión más completa sobre la riqueza y conformación de las comunidades de palmas. Por otro lado, siguiendo el mismo patrón neotropical, en el Chocó biogeográfico las palmas parecen representar un recurso importante para sus pobladores (Bernal & Galeano 1993, García et al. 2002), a juzgar por algunos estudios publicados (Castro et al. 2004, Valois & Pino 2004), en los cuales se ha encontrado que las palmas constituyen la familia de plantas con mayor número de usos en algunas áreas. En este contexto, este trabajo pretende contribuir al conocimiento de las comunidades de palmas del Chocó biogeográfico, mediante el estudio detallado de la riqueza, la abundancia, la composición, la estructura y los usos dados en dos sitios de muestreo en la selva central del Chocó. Se espera que la información resultante aporte al entendimiento de los patrones de distribución de las palmas en el Neotrópico y en Colombia, pero que también sirva de apoyo para la toma de medidas de conservación y manejo para este grupo de gran importancia en los bosques del Chocó y en las dinámicas sociales y culturales de las comunidades humanas.
ÁREA DE ESTUDIO
El presente estudio se realizó en dos localidades en el centro del Departamento de Chocó.
1. Tutunendo. Ubicada en el corregimiento de Tutunendo, municipio de Quibdó, 05°44´41.7Norte -76°32´24.2Oeste, a 150 m de elevación, con temperatura megatermal (25.7¬27.9ºC), y una precipitación promedio anual de 10000 mm (Poveda et al. 2004). La zona presenta una topografía quebrada y con numerosos cursos de agua; los suelos son ácidos y de fertilidad moderada. El área está cubierta de bosque en diferentes estados de intervención; en las áreas mejor conservadas presenta un dosel de 20¬-25 m de altura, con especies arbóreas como chanó (Humiriastrum procerum (Little) Cuatrec.), trúntrago (Vitex cooperi Standl.), carrá (Huberodendron patinoi Cuatrec.), y algarrobo (Hymenaea sp.).
2. Angostura. Ubicada en la parte alta del río San Juan, municipio de Tadó, corregimientos de Angostura y Playa de Oro, 05°17´26 N-76°26´0.73 O, a unos 160 m de altitud, con una temperatura media anual de 23.5¬-27.5ºC, y una precipitación promedio anual de 8000 mm (Poveda et al. 2004). El bosque original de la zona presenta un dosel de 20¬-25 m de altura, donde sobresalen algunas especies como Brosimum utile (Kunth) Oken ex J. Presl., Huberodendron patinoi Cuatrec., Campnosperma panamensis Standl. y Carapa guianensis Aubl., entre otras. La zona ha sido fuertemente transformada por la actividad minera, y ahora está notoriamente fragmentada. El estudio se realizó en los relictos boscosos.
MÉTODOS
Muestreo de las comunidades de palmas. El muestreo se realizó en transectos que se distribuyeron de manera sistemática, teniendo en cuenta que la distancia mínima entre transectos fuera de 20 m, que no se sobrelaparan, ni existieran en su interior zonas muy alteradas como caminos amplios o claros de bosque. En Tutunendo se realizaron 14 transectos, diez de 100x5 m (5000 m2) y cuatro de 500x5 m (10000 m2), para un área total de 1.5 ha. En Angostura se establecieron 12 transectos, diez de 100x5 m (5000 m2) y dos de 500x5 m (5000 m2), correspondientes a un total de 1 ha. La delimitación de los transectos se hizo utilizando una cuerda como eje central y dos varas de madera de 2.5 m a lado y lado de la cuerda. En cada transecto se hicieron colecciones de todas las especies, las cuales fueron depositadas en el Herbario Chocó, de la Universidad Tecnológica del Chocó, en Quibdó, y en el Herbario Nacional Colombiano (COL), del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, en Bogotá. En cada transecto se registraron todas las palmas, incluyendo todos los tamaños (plántulas, juveniles y adultos), según los parámetros de Kahn & Castro (1985). Se definieron como arbustivas las palmas menores de 4 m, que alcanzan su máximo desarrollo a nivel del sotobosque; como arbóreas, aquellas que sobrepasan 5 m de altura (usualmente mayores de 10 m de altura) y que alcanzan su máximo desarrollo a la altura del dosel; acaules, las que no poseen tallo aéreo; y trepadoras, aquellas que presentan estructuras o modificaciones para alcanzar la parte alta del bosque.
Usos de las palmas. Se obtuvo información sobre nombres comunes y usos, directamente en campo, de 16 personas en total (sabios y guías), todos hombres mayores de 45 años, nativos de las localidades y reconocidos por los habitantes como conocedores del bosque.
Análisis de la información
Riqueza, abundancia y composición de especies. Se realizaron curvas especies-área, utilizando los estimadores Singletons, ACE, Chao 1 y Michel Mean (Colwell 2004), y se aplicaron intervalos de confianza (IC) del 95%. La diversidad, la equidad y la dominancia fueron analizadas con los índices de Margalef, Pielou y Simpson, y la diversidad entre localidades (beta) fue evaluada con el índice de Jacard para datos cuantitativos (Moreno 2001, Villareal et al. 2004). Se utilizaron los programas estadísticos Estimates 7.5 (Colwell 2004) y PAST 1.94b (Hammer et al. 2001).
Estructura de la comunidad. Para este parámetro se tuvo en cuenta la importancia ecológica de las especies, la cual se obtuvo utilizando el Índice de Valor de importancia (IVI), calculado según Villareal et al. (2004), como la sumatoria de los valores relativos de la densidad, la frecuencia y la dominancia relativa. Sin embargo, en este estudio el IVI se calculó sólo con la sumatoria de la densidad relativa y la frecuencia relativa. Teniendo en cuenta que las áreas de muestreo tomadas en cada localidad fueron diferentes (1.5 ha para Tutunendo y 1.0 ha para Angostura), para realizar análisis comparativos de las variables (riqueza y abundancia) entre las mismas y con otros estudios en el trópico, se tomó una unidad base de 0.1 ha.
Usos de las palmas. Los usos registrados para las palmas se agruparon en siete categorías de uso, siguiendo a Paniagua et al. 2010 y a Cook (1995).
RESULTADOS
Riqueza y abundancia de especies. Los índices ecológicos para las comunidades de palmas de Tutunendo y Angostura se muestran en la tabla 1.
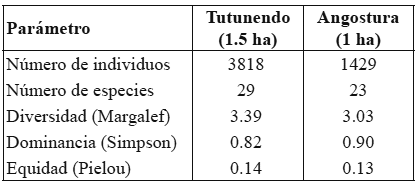
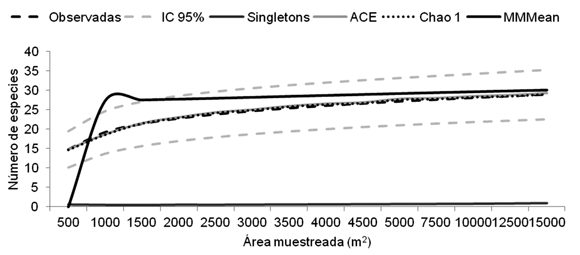
La diversidad representada mediante el índice de Margalef se vio fuertemente influenciada, en ambas comunidades, por el alto número de individuos y la gran abundancia de unas pocas especies. En ambas comunidades dominaron pocas especies (Wettinia quinaria, Welfia regia, Socratea exorrhiza y Geonoma cuneata), situación que se reflejó en el alto valor del índice de Simpson, mientras la equidad fue muy baja, también en ambas comunidades, afectada directamente por la poca abundancia de cerca de diez especies con menos de 25 individuos. El número de especies en los 14 transectos de Tutunendo fue muy alto, incluso para los cinco primeros transectos (2500 m2), donde se registraron 23 especies, correspondientes al 79% de la saturación del área de muestreo, según el indicador Michel M (Figura 1).
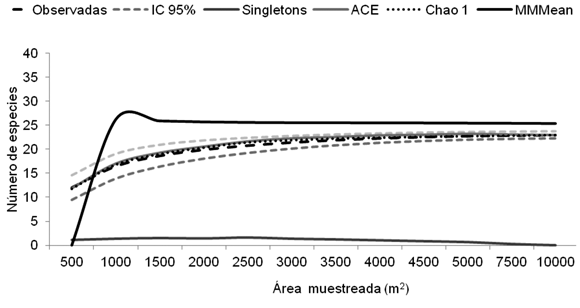
La curva mostró un leve aumento desde el transecto 6 al 10, donde se observaron valores progresivos que fueron desde 24 hasta 26 especies acumuladas (83 y 90% de la saturación de la comunidad de palmas). En Angostura (Figura 2), las especies observadas en el transecto número uno (12) aumentaron significativamente a medida que se incrementó el esfuerzo de muestreo: en 3500 m2 se registraron 21 especies, es decir, 91% de la saturación del área para esta comunidad, según el indicador Michel M (MM); la curva mostró una estabilidad a partir de los 4500 m2, donde se observaron 23 especies que saturan el área total según el estimador MM, y se mantuvo constante hasta los 10 000 m2.
Composición de la comunidad de palmas. Las especies de palmas encontradas en Tutunendo y Angostura, con sus correspondientes autores, y sus valores de importancia ecológica se muestran en la tabla 2. En 14 transectos (1.5 ha) de Tutunendo se registraron 3818 individuos (1233 adultos), pertenecientes a 18 géneros y 29 especies. En cuanto a la hectárea estudiada en Angostura (10 transectos de 100 x5 m y 2 de 500x5m), se registraron 1429 individuos (529 de ellos adultos), pertenecientes a 15 géneros y 23 especies. Entre las dos localidades se registraron 33 especies, 10 de las cuales (30%) fueron exclusivas de Tutunendo y 4 (12%) se registraron solo en Angostura; las restantes 19 especies (58%) fueron comunes a las dos localidades (Índice de Jacard = 0,58). A nivel genérico, 15 de los 18 géneros registrados son compartidos por las dos localidades. Los valores de importancia ecológica de las especies (tabla 2) muestran que en Tutunendo y Angostura, 62% y 42% de los individuos de palmas, respectivamente, estuvo constituido por sólo tres especies (Wettinia quinaria, Welfia regia y Socratea exorrhiza).
Las especies con mayor índice de valor de importancia tanto en Tutunendo como en Angostura, fueron W. quinaria, W. regia, S. exorrhiza, G. cuneata y Pholidostachys dactyloides; estas cinco especies no solo son las más frecuentes, apareciendo en casi todos los transectos, sino que también dominan por sus altas densidades; el 50% y 45% del total del IVI en Tutunendo y Angostura, respectivamente, se concentró en estas cinco especies. En cuanto a las especies raras, se destacan Desmoncus cirrhifer, presente sólo en Tutunendo, con un único individuo; e Iriartea deltoidea, muy abundante en Tutunendo, pero con sólo 25 individuos en Angostura.
Distribución de hábitos de crecimiento y estructura. La riqueza de palmas, tanto en Tutunendo como en Angostura, se concentró en gran parte a nivel del sotobosque, donde se encontraron 15 y 13 especies, respectivamente, más de la mitad de ellas de hábito cespitoso. En el dosel de Tutunendo y Angostura, se encontraron 11 y 8 especies de porte arbóreo, respectivamente, más de 67% de ellas de tronco solitario; sólo se encontraron 3 palmas acaules en ambos sitios, y sólo una especie trepadora (Desmoncus cirrhifer), presente solo en Tutunendo. Con respecto a la estructura de las poblaciones de comunidades las palmas en Tututnendo y Angostura, el mayor porcentaje de individuos se encontró en la categoría Juveniles (40%, 46%, respectivamente), seguido de los adultos (32% y 37%) y por último de las plántulas (28%, 17%).
Usos de las palmas
De las 33 especies registradas en las localidades de estudio, 27 tuvieron registros de uso por los habitantes de las comunidades. Los usos se agruparon en las siguientes siete categorías (tabla 3): Materiales (19 spp.), que incluye implementos para caza y pesca y utensilios para uso doméstico y de trabajo; Construcción (17 spp.), que incluye techos, paredes y pisos y construcciones relacionadas y además la construcción de embarcaciones; Alimentación Animal (6 spp.) y Alimentación Humana (4 spp.); Medicinal (6 spp.); Social, incluyendo cosméticos y objetos ceremoniales (3 spp.); y finalmente, una especie usada como Insecticida.
Se registraron seis partes de las palmas que son utilizadas: tallo (21 spp.), hojas (13 spp.), frutos y semillas (10 spp.), brácteas (2 spp.) y raíces (1 spp.). La especie con mayor número de usos registrados fue Oenocarpus bataua, ya que se registró en seis de las siete categorías, con el aprovechamiento de cuatro de sus órganos (tallo, hojas, brácteas y frutos).
DISCUSIÓN
Riqueza de especies. La riqueza de palmas encontrada en las dos localidades estudiadas (29 especies en 1.5 ha y 23 especies en 1 ha, para un total de 33 especies en 2.5 ha), corrobora que la riqueza registrada por Gentry (1986) para un transecto de un área cercana, en Tutunendo (17 especies de palmas en 0.1 ha.) no era un hecho aislado, y confirma la alta riqueza de palmas en la región del Chocó. Los valores de riqueza de palmas encontrados en este estudio están entre los más altos registrados hasta el momento para el Neotrópico (Balslev et al. 2011), superados sólo por varios registros de Iquitos y áreas cercanas en el Occidente de la Amazonia (Vormisto et al. 2004, Montúfar & Pintaud 2006), y de los bosques húmedos de Costa Rica (Balslev et al. 2011). Esta alta riqueza de especies encontrada apoya algunas de las hipótesis que se han planteado sobre los patrones de distribución de las palmas en el Neotrópico (Bjorholm et al. 2005, Svenning et al. 2008), que indican que los factores más determinantes de la riqueza de palmas en el Neotrópico son la alta precipitación, la disponibilidad de agua a lo todo largo del año, las altas temperaturas y las bajas latitudes, factores que se han encontrado también ligados a la riqueza en otros organismos como las aves (Rahbek & Graves 2001). La alta diversidad de los géneros Geonoma y Bactris encontrada en las localidades del centro del Chocó, confirma un patrón fitogeográfico de todas las tierras bajas y húmedas del Neotrópico, pues estos géneros han sido reportados también como los más diversos en otras áreas del Neotrópico (Henderson et al. 1995). Por otro lado, el comportamiento de los estimadores de riqueza Chao 1 y ACE y su relación con los datos de las especies observadas, sugiere que los valores de riqueza observados son representativos y que el área evaluada fue suficiente para registrar la riqueza específica de las localidades. Es interesante destacar que la cifra de la riqueza de palmas encontrada en las dos localidades de estudio resulta impresionante si se compara con los listados de palmas de Galeano & Bernal (2004, 2010), pues las palmas encontradas (33 especies en sólo 2.5 ha) corresponden al 48% del total de especies de palmas (69) registradas para todo el departamento del Chocó. Este hecho concuerda con la hipótesis discutida por Kristiansen et al. (2011) y Balslev et al. (2011), que plantea que la diversidad alfa (el alto número de especies en una localidad) está influenciada por la diversidad gama (alta riqueza de especies en una región), lo cual parecería constituir un patrón general en el Neotrópico (Valencia et al. 1994, Balslev et al. 1998).
Abundancia. La diferencia en la densidad de palmas entre las dos localidades (en promedio 254 individuos por 0.1 ha en Tutunendo y 143 individuos por 0.1 ha en Angostura) podría ser explicada por los siguientes factores: a. Distintos regímenes de lluvia: 11000 mm en Tutunendo y cerca de 8000 mm en Angostura, y b. La variación fisionómica del bosque, dado que en Tutunendo la cobertura vegetal estuvo constituida por una matriz de bosque continuo, mientras que en Angostura se presentó un bosque en su mayoría discontinuo, conformado por parches que soportan las palmas de la localidad, donde posiblemente no son tan eficientes los procesos de dispersión de semillas y establecimiento de nuevos individuos, situación que se evidenció con la abundancia de especies como Wettinia quinaria e Iriartea deltoidea, que fueron compartidas por las dos localidades pero con mayores abundancias para Tutunendo. Con todo, la comparación de las densidades de palmas encontradas en el centro del Chocó, con otros datos disponibles (individuos con DAP = 10 cm) para otras áreas del Chocó biogeográfico, muestran diferencias importantes. En la zona central del Chocó (donde la precipitación fluctúa entre 8000 y más de 10000 mm anuales) se registraron 32 y 70 individuos de palmas DAP = 10 cm por 0.1 ha (para Angostura y Tutunendo, respectivamente), y en otro estudio en la localidad de Salero, también en el centro del Chocó, Asprilla et al. (2003) registraron un promedio de 27 palmas = 10 cm DAP por 0.1 ha. En contraste, en la Costa Pacífica del Chocó (ca. 7000 mm de lluvia anual), Galeano (2001) registró un promedio de 12 individuos de palmas con DAP = 10 cm por 0.1 ha, y en el Bajo Calima (sur del Chocó Biogeográfico, con 6800 mm de precipitación anual), Faber-Langendoen & Gentry (1991) registraron un promedio de 13 individuos de palmas con DAP = 10 cm por 0.1 ha. Podría pensarse entonces que la abundancia de palmas aumenta hacia el centro de la región del Chocó. Este patrón de aumento de densidad de palmas hacia los bosques centrales, también parecería entonces estar relacionado positivamente, al igual que la riqueza de palmas, con la precipitación. La baja densidad de algunas especies (Aiphanes acaulis, Chamaedorea deneversiana, Desmoncus cirrhifer, Hyosphate elegans, Mauritiella macroclada, Prestoea decurrens y Geonoma chococola, entre otras), probablemente esté relacionada con las altas densidades de las especies comunes y generalistas (Wettinia quinaria, Welfia regia, Geonoma cuneata, entre otras), en concordancia con lo planteado por Pitman et al. (2001), quienes sugieren que las especies oligárquicas dominantes pueden ser generalistas ecológicos, en contraste y compitiendo con las especies raras, que son ecológicamente especializadas.
Composición. A pesar de estar separadas por cerca de 86 km y de corresponder, según Forero & Gentry (1989), a regiones fitogeográficas diferentes (Selva Pluvial Central en Tutunendo, y Subregión del San Juan en Angostura), con regímenes de precipitación diferente), las comunidades de palmas de estos dos sitios mostraron una gran similitud en la composición: el 83% de los géneros y el 58% de las especies fueron compartidos, de tal forma que por lo menos en términos de composición de las comunidades de palmas, parecería no haber diferencias entre esas regiones fitogeográficas. Los datos encontrados muestran que debido a las altas densidades y frecuencias, algunas palmas como Wettinia quinaria, Welfia regia, Iriartea deltoidea, Geonoma cuneata, Geonoma calyptrogynoidea y Pholidostachys dactyloides, modelan la estructura de las comunidades de palmas en las áreas estudiadas, contribuyendo de forma directa a la arquitectura y estructura de la vegetación. Por otro lado, dado el gran aporte de biomasa y de disponibilidad de hábitats y frutos para la fauna local, es de suponer que estas palmas constituyan además, un recurso importante en los niveles tróficos locales. Situación similar se reporta para otros bosques: en el sur del Chocó Biogeográfico (Bajo Calima,Valle el Cauca), las especies de palmas arbóreas dominantes son Socratea exorrhiza, Wettinia quinaria y Welfia regia (Faber-Langendoen & Gentry 1991); en la Costa Pacífica de Chocó se registró Welfia regia como la especie arbórea más abundante (Galeano 2001); en una síntesis de la región Pacífica, Rangel (2004), identifica a Wettinia quinaria, Welfia regia e Iriartea deltoidea como elementos importantes de ecosistemas y de asociaciones biológicas a lo largo del Chocó Biogeográfico. Con base en esta información, proveniente de diferentes áreas en sitios extremos de las zonas bajas del Chocó Biogeográfico y en otros datos para el centro del Chocó (Asprilla et al. 2003, Mosquera et al. 2007), se puede concluir, no sólo que las palmas en su conjunto son claves en los ecosistemas, sino también que hay un grupo de especies de palmas que son comunes, abundantes y frecuentes a lo largo y ancho de la región. Esto coincide también con la hipótesis de oligarquía propuesta por Pitmann et al. (2001). Estas especies incluyen, por lo menos, a Wettinia quinaria, Welfia regia, Socratea exorrhiza, Iriartea deltoidea y Oenocarpus bataua a nivel del estrato arbóreo, y a Geonoma cuneata, a nivel del sotobosque. Además, la reiterada presencia de los individuos de estas especies indica que, por lo menos a nivel regional, tienen un alto potencial para colonizar diferentes ambientes, resultado de alta producción de semillas, dispersores eficientes, buena germinación y capacidad para establecerse y persistir. Por lo menos para Iriartea deltoidea y las especies de Wettinia se ha señalado que en los bosques donde crecen, la alta precipitación genera un abundante epifitismo que ocasiona caída de grandes ramas, y una alta tasa de caída de árboles, lo cual produce pequeños claros que favorecen el desarrollo de esas especies (Galeano & Bernal 2010). Por otra parte, al comparar los datos de composición, abundancia y riqueza de las comunidades de palmas de las dos localidades (Tutunendo y Angostura), ubicadas en el centro del Chocó Biogeográfico colombiano, con los datos del bosque húmedo de Costa Rica, se observa una gran similaridad. De los 15 géneros que reporta Wang (2008) para los bosques húmedos de la estación biológica La Selva, 10 géneros (63%) son compartidos. La similitud también se observa a nivel de riqueza y composición de especies: de las 24 especies reportadas para los muestreos en Costa Rica, 10 son compartidas con Tutunendo y Angostura. En los muestreos de Wang (2008) se observó también un dominio en la comunidad de palmas de las especies de dosel Welfia regia y Socratea exorrhiza; así mismo, en el sotobosque se comparten las especies Geonoma cuneata, Geonoma deversa y Bactris hondurensis. Estas similitudes en la flora de palmas permiten apoyar la hipótesis planteada, entre otros, por Lellinger & de La Sota (1978), Galeano et al. (1998a, 1998b), Galeano (2001) y Mora et al. (2006), quienes sugieren que el Chocó Biogeográfico se extiende hasta las tierras bajas de Costa Rica, por la similaridad estructural y florística y por los elementos compartidos a lo largo de los bosques de tierras bajas de Centro América.
Estructura de las comunidades de palmas. Aunque la delimitación por clases de tamaño utilizada en este estudio fue demasiado gruesa para permitir inferencias detalladas, la estructura de la población encontrada tanto en Tutunendo como en Angostura, con un muy bajo porcentaje de plántulas con respecto a los juveniles y adultos, podría sugerir que existen problemas de reclutamiento a nivel general en la comunidad, pues lo que se esperaría para una comunidad de palmas en buen estado es que haya más plántulas que juveniles y que adultos, es decir, que la estructura muestre una curva en forma de J invertida. Sin embargo, esta estructura no fue común para todas las especies: el comportamiento estructural de las poblaciones de las especies de palmas más importantes de las dos localidades, mostró que Wettinia quinaria no solo es dominante (36% y 21% de los individuos de las palmas de Tutunendo y Angostura, respectivamente) y el elemento de dosel más frecuente, apareciendo en todos los transectos, sino que también posee una población bien estructurada, con un modelo de distribución de J-invertida típica de poblaciones sanas y en crecimiento. La situación fue un poco diferente para las poblaciones de otras especies importantes por sus densidades y frecuencia como Welfia regia, Socratea exorrhiza, Pholidostachys dactyloides e Iriartea deltoidea; es decir, que los problemas de reclutamiento de la comunidad están reflejando los problemas de reclutamiento de estas especies. Esta situación se puede explicar desde varios escenarios: a) problemas de reclutamiento a nivel de la comunidad debido a la fragmentación (más fuertes en Agostura), pues las plántulas son la clase de tamaño más vulnerable a este proceso (Scariot 1999); y b. La forma de aprovechamiento y las partes utilizadas, si tenemos en cuenta que para las dos localidades se registra un alto potencial de uso de tallos y frutos, lo que puede estar influyendo directamente en la producción de semillas, el reclutamiento de las mismas y el establecimiento al interior de los bosques. Específicamente, el bajo reclutamiento observado para especies como Iriartea deltoidea y Socratea exorhiza, muy posiblemente está relacionado con el uso de los troncos de estas especies en construcción. Para las otras especies de las cuales no se utilizan los troncos, el inadecuado corte de los tallos para colectar las hojas, podría ser el factor que esté provocando el bajo reclutamiento. Aunque la situación aún no es tan dramática, de continuar con los procesos de fragmentación y formas de uso, a largo plazo se podrían generar cambios en la composición de la comunidad de palmas ocasionados por la colonización parcial o total de especies generalistas con buena estructura poblacional como Wettinia quinaria, mientras las especies naturalmente más escasas como Aiphanes acaulis, Chamaedorea deneversiana, Geonoma chococola, que necesitan ambientes conservados, desaparecerían, afectando directamente la riqueza de las palmas y a su vez la vida silvestre que aprovecha el recurso que estas ofrecen. Aunque las dos localidades presentan problemas a nivel general de la estructura de las comunidades de palmas, éstos son más acentuados en Angostura, lo que podría estar relacionado con una mayor fragmentación del área en relación con Tutunendo, ya que según lo encontrado por Baez & Balslev (2007) en bosques del occidente ecuatoriano, el efecto de borde en los bosques fragmentados también contribuye a la alteración de las comunidades de palmas. Con relación a los hábitos de crecimiento, los datos son claros en demostrar que las palmas ocupan todos los estratos del bosque, lo cual también podría interpretarse como un tipo de mecanismo que permite a numerosas especies de palmas diferentes coexistir simpátricamente, tal como se ha señalado para aves y otros organismos (Terborgh 1992). Sin embargo, el hábito de crecimiento con mayor riqueza en las localidades de estudio fue el arbustivo (palmas del sotobosque), que estuvo dominado por Geonoma calytrogynoidea, G. cuneata, G. deversa, G. divisa y G. stricta; Bactris coloniata y B. hondurensis; Pholidostachys dactyloides y P. pulchra. Esta dominancia y riqueza de palmas a nivel del sotobosque es, al parecer, un patrón común también para los bosques de la Amazonia (Kahn et al. 1988, Balslev et al. 2010). Por otro lado, el hábito acaule fue poco diverso, con dos especies poco abundantes, Aiphanes acaulis y Attalea allenii, situación que se puede atribuir a la necesidad estricta de estas especies de ambientes particulares con más humedad (A. acaulis), o sitios abiertos y claros de bosque (A. allenii), los cuales se presentaron poco en los transectos. El hábito trepador fue el más escaso, con un único individuo de Desmoncus cirrhifer presente en Tutunendo; esta baja representatividad podría estar relacionada con las especificidades ecológicas para su germinación y establecimiento, baja dispersión de semillas o dispersión limitada; las observaciones de esta especie realizadas por Castro et al. (2004), Valois et al. (2004) y Ramírez (2007), en bosques de localidades cercanas, también sugieren que es una especie escasa y de distribución dispersa. Por otro lado, en las dos localidades dominaron las palmas de tallo solitario (18 de las 33 especies), la mayoría de tamaño arbóreo, lo que muestra concordancia con lo observado por Ruokolainen & Vormisto (2000), quienes lo relacionan con mecanismos ecológicos que permiten a las palmas arbóreas tener acceso a dispersores efectivos (aves pequeñas, móviles y que recorren mayores distancias) de forma más fácil y frecuente. En las dos localidades estudiadas se destacaron especies solitarias como Wettinia quinaria, W. equalis, W. radiata, Welfia regia, Socratea exorrhiza y Oenocarpus bataua, que presentaron grandes infrutescencias con numerosos frutos. En contraste, especies cespitosas como Bactris barronis, B. brongniartii, B. coloniata, B. hondurensis, Geonoma cuneata y G. deversa, entre otras, fueron observadas con una baja producción de frutos, lo cual posiblemente es compensada con la producción de nuevos individuos a partir de tallos subterráneos y de brotes. Una situación similar fue discutida por Balslev et al. (2010, 2011), quienes la interpretaron como una estrategia reproductiva de división de la comunidad en gremios diferentes y complejos, que permitirían, en última instancia, la coexistencia de una mayor riqueza de las especies en una comunidad dada.
Usos de las palmas. Las siete categorías de uso asignadas a 27 de las 33 especies encontradas, muestran la importancia y la variada utilización de las palmas para las poblaciones locales. El panorama de aprovechamiento en ambas localidades muestra que especies con un número menor de usos registrados (uso de dos órganos en dos o tres categorías de uso), como es el caso de Iriartea deltoidea (barrigona), y Welfia regia (amargo), son ampliamente apreciadas por los pobladores y por ello son más frecuentemente empleadas en las labores cotidianas. Sin embargo, los datos estructurales de las poblaciones de palmas usadas podrían sugerir que el uso de las especies está teniendo un impacto negativo. Por ejemplo, para Oenocarpus minor y O. bataua, se observó una estructura poblacional que evidencia una situación dramática, con más adultos (68% y 58%, respectivamente) y juveniles (29% y 40%, respectivamente) que plántulas (3% y 2%, respectivamente), lo que se puede explicar dado que la categoría de uso alimenticia requiere del aprovechamiento de los frutos maduros, lo que podría incidir directamente sobre un muy deficiente reclutamiento de estas especies en las zonas estudiadas. De hecho, es bien conocido que los frutos de estas dos especies están entre los más apetecidos en toda la región del Chocó y que desafortunadamente, en la mayoría de los casos, las palmas son derribadas para cosechar los frutos. Esta situación plantea la urgencia de hacer estudios sobre la dinámica de estas especies que son más utilizadas, como Iriartea deltoidea, Welfia regia, Socratea exorrhiza, Oenocarpus bataua y Oenocarpus minor, con el fin de elaborar planes de manejo y modelos productivos que permitan el uso y la conservación a largo plazo sus poblaciones. En conclusión, los resultados de este estudio corroboran que la comunidad de palmas del Chocó es una de las más ricas del Neotrópico y apoyan las hipótesis relacionadas con la distribución de la riqueza y la abundancia de palmas asociadas a la precipitación. También se evidenció la versatilidad de uso de las palmas para los pobladores del Chocó. Se recomienda el desarrollo de estudios detallados sobre las comunidades de palmas en otros sectores del Chocó Biogeográfico, con el fin de refinar el conocimiento y probar las hipótesis existentes.
AGRADECIMIENTOS
Esta contribución hace parte de la tesis de Maestría en Ciencias-Biología, Universidad Nacional de Colombia, Línea Biodiversidad y Conservación del primer autor. Agradecimientos al Instituto de investigaciones del Pacífico (IIAP) y a su director William Klinger, por el apoyo a esta investigación en todas sus fases; a la Universidad Nacional de Colombia y a su Instituto de Ciencias Naturales por el apoyo institucional y logístico. El primer autor agradece a los profesores Rodrigo Bernal, Orlando Rangel, Jaime Aguirre, Diego Giraldo, Orlando Rivera, Agustín Rudas, Olga Montenegro, y Argenis Bonilla por la formación impartida, base para el desarrollo de esta investigación. A los profesores Orlando Rivera y Agustín Rudas, Henrik Balslev y a dos evaluadores anónimos por la revisión crítica de versiones anteriores de este manuscrito. A los integrantes del Grupo de Investigación en Palmas Silvestres Neotropicales. A Luz del Carmen Moreno, Lady Vargas, Zulmary Valoyes, Hamleth Valois quienes apoyaron incondicionalmente, tanto en campo, como en el análisis de los resultados.
LITERATURA CITADA
1. ASPRILLA, A., C. MOSQUERA & F. GARCÍA. 2003. Composición Florística de un bosque pluvial tropical (bp-T) en la parcela permanente de investigación en biodiversidad (PPIB) en Salero, Unión Panamericana, Chocó. Págs. 39-44 en F. García, Y. Ramos, J. Palacios, E. Arroyo, A. Mena, & M. González (eds.), Salero: Diversidad Biológica de un Bosque Pluvial Tropical (bp-T). Editora Guadalupe Ltda, Bogotá, D.C. [ Links ]
2. BAEZ, S. & H. BALSLEV. 2007. Edge effects on palm diversity in rain forest fragments in western Ecuador. Biodiversity and Conservation 16(7): 2201-2211. [ Links ]
3. BALSLEV, H., W. EISERHARDT, T. KRISTIANSEN, D. PEDERSEN & C. GRANDEZ. 2010. Palms and palm communities in the upper Ucayali River Valley a little-known region in the Amazon Basin. Palms 54(2): 57–72. [ Links ]
4. BALSLEV, H., F. KAHN, B. MILLAN, J-C. SVENNING, T. KRISTIANSEN, F. BORCHSENIUS, D. PEDERSEN &W. EISERHARDT. 2011. Species Diversity and Growth Forms in Tropical American Palm Communities. Botanical Review (DOI 10.1007/s12229-011-9084-x). [ Links ]
5. BALSLEV, H., R. VALENCIA, G. PAZ Y MIÑO, H. CHRISTENSEN & I. NIELSEN. 1998. Species count of vascular plants in 1-hectare of humid lowland forest in Amazonian Ecuador. Págs. 591-600. En: F. Dallmeier & J. A. Comiskey (eds.). Forest Biodiversity in North, Central and South America and the Carribean: Research and Monitoring. Man and the Biosphere Series, vol. 21. Unesco and TheParthenon Publishing Group, Paris. [ Links ]
6. BERNAL, R. & G. GALEANO. 1993. Palmas del Andén Pacífico. Págs. 220-231 en: P. Leyva (ed). Colombia Pacífico Tomo I. Editorial del Fondo FEN, Bogotá, D.C. [ Links ]
7. BJORHOLM, S., J-C. SVENNING, F. SKOV & H. BALSLEV. 2005. Environmental and spatial controls of palm (Arecaceae) species richness across the Americas. Global Ecology and Biogeography 14: 423-429. [ Links ]
8. BJORHOLM, S., J-C. SVENNING, W. J. BAKER, F. SKOV & H. BALSLEV. 2006. Historical legacies in the geographical diversity patterns of New World palm (Arecaceae) subfamilies. Botanical Journal of the Linnean Society 151: 113–125. [ Links ]
9. BJORHOLM, S., J.-C. SVENNING, F. SKOV & H. BALSLEV. 2008. To what extent does Toblers 1st law of geography apply to macroecology? A case study using American palms (Arecaceae). BMC Ecology 8: 11. [ Links ]
10. CASTRO, A., D. ABADÍA & N. PINO. 2004. Plantas Silvestres Alimenticias de uso tradicional en las comunidades de Pacurita, San José de Purré y Guayabal. Municipio de Quibdó, Chocó. Revista Institucional. Universidad Tecnológica del Chocó 18:37-42. [ Links ]
11. CLARK, D. A., D. B. CLARK., M. R. SANDOVAL & C. M. V. CASTRO. 1995. Edaphic and human effects on landscape-scale distributions of tropical rain forest palms. Ecology 76: 2581–2594. [ Links ]
12. COLWELL, R. K. 2004. EstimateS, Versión 7: Statistical Estimation of species richness and shared species from samples (Software and users Guide). Freeware for Windows and Mac OS. [ Links ]
13. COOK, F. 1995. Economic Botany Data collection standard. Royal Botanic Gardens Kew. Londres. [ Links ]
14. FABER-LANGENDOEN, D. & A. GENTRY. 1991. The structure and diversity of rainforest at Bajo Calima, Chocó, Western Colombia. Biotropica 23 (1): 2-11. [ Links ]
15. FORERO, E. & A. H. GENTRY. 1989. Lista Anotada de las plantas del Departamento del Chocó, Colombia. Biblioteca José Jerónimo Triana # 10. Instituto de Ciencias Naturales. Museo de Historia Natural. Editorial Guadalupe, Bogotá, D.C. [ Links ]
16. GALEANO, G. 2001. Estructura, riqueza y composición de plantas leñosas en el golfo de Tribugá, Chocó, Colombia. Caldasia 23 (1): 213-236. [ Links ]
17. GALEANO, G. & R. BERNAL. 2004. Catálogo de espermatofitos en el Chocó Biogeográfico. Familia Arecaceae. Págs. 135-148 en: J.O Rangel-CH. (ed), Colombia Diversidad Biótica IV: El Chocó biogeográfico/Costa Pacífica. Instituto de Ciencias Naturales Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
18. GALEANO, G. & R. BERNAL. 2010. Palmas de Colombia. Guía de campo. Editorial Universidad Nacional de Colombia. Instituto de Ciencias Naturales. Facultad de Ciencias. Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
19. GALEANO, G., J. CEDIEL & M. PARDO. 1998a. Structure and Floristic Composition of a one-hectare plot of wet forest at the Pacific coast of Colombia. Págs. 551- 567 en: F. Dallmeier & A. Comiskey (eds). Forest biodiversity in North, Central and South America, and the Caribbean: Research and Monitoring. Man and the Biosfere Series Vol 21. [ Links ]
20. GALEANO, G., S. SUAREZ & H. BALSLEV. 1998b. Vascular plant species count in a wet forest in the Chocó area on the Pacific coast of Colombia. Biodiversity and Conservation 7: 1563-1575. [ Links ]
21. GARCÍA, F., J. PALACIOS, Y. RAMOS, A. MENA, J. E. ARROYO & M. GONZÁLEZ. 2002. Composición, estructura y etnobotánica de un bosque pluvial tropical (bpT) en Salero Chocó. Revista Institucional Universidad Tecnológica del Chocó 17: 3-9. [ Links ]
22. GENTRY, A. H. 1986. Species richness and floristic composition of Chocó región plant communities. Caldasia 15: 71-91. [ Links ]
23. HAMMER, O., D.A.T. HARPER & P. D. RYAN. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleontology Electronic 4(1): 9. [ Links ]
24. HENDERSON, A., G. GALEANO & R. BERNAL. 1995. Field Guide to the palms of the Americas. Princeton University Press, Princeton, New Jersey. [ Links ]
25. KAHN, F. & A. CASTRO. 1985. The palm community in a forest of central Amazonia, Brazil. Biotropica 17(3): 210-216. [ Links ]
26. KAHN, F., K. MEJIA & A. CASTRO. 1988. Species richness and density of palms in terra firme forest of Amazonia. Biotropica 20(4): 266-269. [ Links ]
27. KRISTIANSEN, T., J.-C. SVENNING, D. PEDERSEN, W. EISERHARDT, C. GRANDEZ & H. BALSLEV. 2011. Local and regional palm (Arecaceae) species richness patterns and their cross-scale determinants in the western Amazon. Journal of Ecology 99 (4) 1001-1015. [ Links ]
28. LELLINGER, D. B. & E. DE LA SOTA. 1978. The phytogeography of the Pteridophytes of the Departamento del Chocó, Colombia. Research Report for National Geographic Society, Washington, DC. [ Links ]
29. MONTÚFAR, R. & J-C-PINTAUD. 2006. Variation in species composition, abundance and microhabitat preferences among western Amazonian terra firme palm communities. Botanical Journal of the Linnean Society 151: 127-140. [ Links ]
30. MORA, M., R. BERNAL, T. CROAT & J. JÁCOME. 2006. Phytogeographic analysis of Araceae of Cabo Corrientes (Chocó, Colombia) and comparable lowland tropical American floras. Annals of the Missouri Botanical Garden 93: 359-366. [ Links ]
31. MORENO, C. E. 2001. Métodos para medir la biodiversidad. M&T–Manuales y Tesis SEA, vol. 1. Zaragoza. [ Links ]
32. MOSQUERA, L., ASPRILLA, A. & D. ROBLEDO. 2007. Diversidad florística de dos zonas de bosque tropical húmedo en el municipio de Alto Baudó, Chocó Colombia. Acta Biológica Colombiana 12 (suppl.1): 75-90. [ Links ]
33. NORMAND, S., J. VORMISTO, J.-C. SVENNING, C. GRANDEZ & H. BALSLEV. 2006. Geographical and environmental controls of palm beta diversity in paleo-riverine terrace forests in Amazonian Peru. Plant Ecology 186: 161-176. [ Links ]
34. PANIAGUA ZAMBRANA, N., M. J. MACÍA & R. CÁMARA-LERET. 2010. Toma de datos etnobotánicos de palmeras y variables socioeconómicas en comunidades rurales. Ecología en Bolivia 45(3): 44-68. [ Links ]
35. PITMAN N.C.A., J. TERBORG, M.R. SILMAN. 2001. Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology 82: 2101-2117. [ Links ]
36. POVEDA, C., C. A. ROJAS-P., A. RUDAS-LL. & O. RANGEL-CH. 2004. El chocó Biogeográfico: Ambiente físico. Págs. 1-21 en: J.O. Rangel-CH. (ed). Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
37. RAHBEK, C. & G. R. GRAVES. 2001. Multiscale assessment of patterns of avian species richness. Proceedings of the National Academy of Sciences 98: 4534-4539. [ Links ]
38. RAMÍREZ, G. 2007. Plantas útiles de dos comunidades negras en los corregimientos de Pacurita y la Troje, Municipio de Quibdó-Chocó. Revista Universidad Tecnológica del Chocó 26(2): 123-126. [ Links ]
39. RANGEL, J. O. (ed). 2004. Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica. Instituto de ciencias Naturales, Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
40. RUOKOLAINEN, K. & J. VORMISTO. 2000. The most widespread Amazonian palms tend to be tall and habitat generalists. Basic Applied Ecology 1: 97-108. [ Links ]
41. SCARIOT, A. 1999. Forest fragmentation effects on palm diversity in central Amazon. Journal of Ecology 87: 66-76. [ Links ]
42. SVENNING, J-C., F. BORCHSENIUS, S. BJORHOLM & H. BALSLEV. 2008. High tropical net diversification drives the New World latitudinal gradient in palm (Arecaceae) species richness. Journal of Biogeography 35: 394-406. [ Links ]
43. TERBORGH, J. 1992. Maintenance of diversity in tropical forests. Biotropica 24: 283-292. [ Links ]
44. VALOIS, H. & N. PINO. 2004. Etnobotánica de Cuatro Comunidades Negras del Municipio de Quibdó, Chocó - Colombia. Lyonia 7(2): 61-69. [ Links ]
45. VALENCIA, R., H. BALSLEV, & G. PAZ Y MIÑO. 1994. High tree alpha diversity in Amazonian Ecuador. Biodiversity and Conservation 3: 21-28. [ Links ]
46. VILLAREAL, H., M. ALVAREZ, S. CORDOBA, F. ESCOBAR, G. FAGUA, F. GAST, H. MENDOZA, M. OSPINA & A. M. UMAÑA. 2004. Manual de Métodos para el Desarrollo de Inventarios de Biodiversidad. Programa de Inventarios de Biodiversidad, Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt, Bogotá, D.C. [ Links ]
47. VORMISTO, J., J-C. SVENNING, P. HALL, & H. BALSLEV. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology 92: 577-588. [ Links ]
48. WANG, Y. H. 2008. Palm community structure and land cover changes in the San Juan Biological corridor, Costa Rica. Biotropica 40(1): 44-54. [ Links ]

