










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCaldasia
versão impressa ISSN 0366-5232versão On-line ISSN 2357-3759
Caldasia v.34 n.1 Bogotá jan./jun. 2012
MARÍA RASAL-SÁNCHEZ
JOEL TRONCOS-CASTRO
CARLOS LIZANO-DURÁN
OSCAR PARIHUAMÁN-GRANDA
DAVID QUEVEDO-CALLE
CONSUELO ROJAS-IDROGO
GUILLERMO E. DELGADO-PAREDES
Jardín Botánico de Valencia. Universitat de València, Valencia, España. maria.rasal@cerai.org
Asociación Chira - Centro de Formación Binacional, Mallares, Sullana, Piura, Perú. jtroncos@asociacionchira.org; carlidur@hotmail.com; pgranda@hotmail.com;
Facultad de Ciencias Biológicas, Universidad Nacional Pedro Ruiz Gallo, Ciudad Universitaria, Juan XXIII 391, Lambayeque, Perú. crojasi2002@yahoo.es; guidelg2001@yahoo.es
RESUMEN
Entre los meses de febrero y marzo de 2008 se realizaron inventarios que permitieron conocer el estado actual de la composición florística y la estructura de la vegetación del bosque montano neotropical de Lanchurán (4°38'S, 79°42'W), provincia de Ayabaca, región Piura, Perú. Se siguió el método propuesto por Gentry para muestreo de plantas leñosas en dos sitios, Los Molinos y La Antena, para un total de 0.5 ha; éstos fueron escogidos por sus características fisiográficas y florísticas. En ambos sitios el análisis de suelo reveló niveles bajos de materia orgánica, nitrógeno y potasio y niveles medios de fósforo asimilable. Se registraron, en el sitio Los Molinos, 399 individuos/0.25 ha con diámetro a la altura del pecho ≥ 2.5 cm, correspondientes a 41 especies, 33 géneros y 25 familias, a lo que se suma una vegetación herbácea conformada por 82 especies, 75 géneros y 48 familias; y en el sitio La Antena, 86 especies, 67 géneros y 41 familias. Las familias presentes con mayor valor ecológico son comunes a la mayoría de los bosques montanos del norte y noroeste del Perú, entre ellas Asteraceae, Lauraceae, Melastomataceae, Rubiaceae y Solanaceae. Para el sitio Los Molinos el Área Basal fue 89.23 m2, destacando la Lauraceae con la mayores valores de cobertura relativa, Índice de Valor de Importancia y Valor de Importancia para la Familia con 38.9, 73.1 y 70.8, respectivamente. Se destaca la acción antrópica sobre la diversidad florística en ambos sitios de estudio.
Palabras clave. Análisis de suelo, Bosque montano neotropical, Diversidad florística, Índice de Valor de Importancia.
ABSTRACT
Floristic inventories were carried out from February to April 2008 to know the current state of the floristic composition and vegetation structure of the neotropical montane forest of Lanchurán (4o38'S, 79o42'W), province of Ayabaca, Piura Region, Peru. The method proposed by Gentry to study woody plants was followed in two localities that were selected because of their physiographic and floristic characteristics, Los Molinos and La Antena (0.5 ha total). In both localities the soil analysis showed low levels of organic matter, nitrogen, and potassium, and medium levels of available phosphorus. In Los Molinos, a total of 399 individuals/0.25 ha with ≥ 2.5 cm diameter at breast height were found (41 species in 33 genera and 25 families). Additionally, a herbaceous vegetation consisting of 82 species (75 genera and 48 families) was recorded. In La Antena, a total of 86 species in 67 genera and 41 families were found. The plant families with high ecological value, which are also common in others montane forest of the northern and northwestern Peru, are the followings: Asteraceae, Lauraceae, Melastomataceae, Rubiaceae, and Solanaceae. In Los Molinos the basal area was 89.2 m2, with Lauraceae having the highest values in relative cover, Importance Value Index and Importance Family Value (38.9, 73.1, and 70.8, respectively). The anthropic action stood out as an influencing factor over the floristic diversity found in both sites studied.
Key words. Soil analysis, Neotropical montane forest, Floristic diversity, Importance Value Index.
Recibido: 14/09/2011
Aceptado: 09/05/2012
INTRODUCCIÓN
Los bosques montanos, bosques andinos o bosques montanos de los Andes tropicales, se extienden por varios países de América del Sur como Perú, Ecuador, Colombia, Bolivia y Venezuela. La importancia de estos bosques radica en que son reservorios de biodiversidad y fundamentales en la provisión de servicios ecosistémicos principalmente vinculados al agua, a la regulación climática regional y a la captura y almacenamiento de carbono; son catalogados como ecosistemas frágiles y estratégicos por estar situados en zonas de recarga de cuencas hidrográficas que proveen agua que benefician a más de 40 millones de personas en Bolivia, Ecuador y Perú (Cuesta et al. 2009). Específicamente, los bosques montanos pluviales (subandinos, andinos y altoandinos) presentan una dinámica hídrica poco convencional (Bruijnzeel 2001), donde la niebla y la lluvia, que es transportada por el viento, se convierten en un aporte adicional de agua al sistema (Tobón & Arroyave 2007).
La diversidad florística que albergan los bosques montanos tropicales ha sido estudiada y documentada por varios estudios durante los últimos años (Churchil et al. 1995, Gentry 1995, Jørgensen et al. 1995, Myers et al. 2000, Hamilton 2001, Kappelle & Brown 2001). Muchos de los cuales se realizaron para establecer modelos de distribución espacial (Condit et al. 2000), implementar planes de conservación urgente (Arroyo et al. 2008), establecer patrones de diversidad alfa (López & Duque 2010) y analizar la influencia de gradientes ambientales o ecológicos en los patrones de diversidad y composición florística (Homeier et al. 2010, Loza et al. 2010).
Los bosques montanos, en el sistema de ecorregiones naturales del Perú, propuesto por Brack & Mendiola (2004), conforman la ecorregión selva alta, específicamente los bosques de lluvia de las vertientes occidentales, donde el bosque de neblina constituye el piso intermedio de los tres pisos vegetacionales diferenciados. Los árboles alcanzan alturas de hasta 15 m y están cubiertos por epífitas: bromelias, orquídeas, helechos, musgos y líquenes; también se distingue la presencia de helechos arbóreos. La importancia de estos bosques radica en que son fuente de agua y mantienen el régimen de flujo natural para diversos ríos de las regiones Tumbes, Piura, Lambayeque y La Libertad, que hacen posible actividades agrícolas sostenibles; por otro lado, contienen numerosas especies endémicas de la flora y fauna nacionales. En efecto, en los bosques montanos de Cajamarca y Piura se han descrito varias especies nuevas de fanerógamas como Trixis montesecoensis, Miconia laciniata, Hydrocotyle sagasteguii, Nasa dillonii y Tetrapterys dillonii, en el bosque montano de Monteseco; Chrysophyllum contumacense, Critoniopsis oblongifolia y Valeriana sagasteguii, en el bosque de Cachil y registra la presencia de importantes especies forestales como Alnus acuminata, Nectandra laurel, Weinmannia elliptica y Podocarpus oleifolius (Sagástegui et al. 2003). En cuanto a la flora endémica del departamento de Cajamarca, se han registrado 32 especies endémicas en el bosque montano de Cutervo, 13 especies en Cachil y 11 especies en Monteseco y Taulis, destacando especies de los géneros Critoniopsis, Siphocampylus, Jaltomata y Larnax (Hensold 1999). Referente a la fauna nativa, con algunos mamíferos en peligro de extinción, se tiene al "oso de anteojos" (Tremarctos ornatus), "puma" (Puma concolor), "jaguar" (Panthera onca) y "tigrillo" (Leopardus pardalis); varias aves como el "quetzal de cabeza dorada" (Pharomachrus auriceps), "gallito de las rocas" (Rupicola peruviana), "guácharo" (Steatornis caripensis) y "pava de monte" (Penelope montagnii) y numerosos reptiles, batracios y peces (Sagástegui et al. 2003).
En la actualidad estos bosques tienen una importancia en conservación ya que el 90% de los bosques montanos del norte del Perú han desaparecido, como consecuencia de la actividad antrópica (construcción de carreteras, adjudicación de tierras y sobreexplotación maderera, entre otras acciones), tal como lo indicó recientemente el New York Botanical Garden (Pro Naturaleza 2004). Adicionalmente, en los bosques montanos del noroccidente peruano se encuentran varias especies de Podocarpaceae, escasos representantes de las gimnospermas en la flora nacional, donde Podocarpus oleifolius "olivo", es considerado el biotipo principal y más importante en la dinámica de este ecosistema (Vicuña-Miñano 2005, Mostacero et al. 2006), que si bien es cierto, a pesar de tener una amplia distribución geográfica, en el norte del Perú existen poblaciones muy reducidas, sobreexplotadas y relegadas a pocos individuos en áreas no protegidas por el estado (Arroyo et al. 2008).
Los bosques montanos del Perú, en especial los del noroeste, pertenecientes a la zona fitogeográfica Amotape-Huancabamba, han resultado de mayor endemismo, producido por la complicada topografía y ecología de sus hábitats (Weigend 2002, Weigend et al. 2005), como lo demostraron los escasos inventarios florísticos realizados en los últimos relictos de bosques montanos del noroeste del Perú, específicamente en las regiones de Cajamarca y Piura, al oeste del río Marañón y al sur de la deflexión del Huancabamba (Gentry 1989, Dillon 1994, Dillon et al. 1995, Sagástegui et al. 2003). En la región Piura los trabajos son aún escasos contándose con el estudio realizado en el bosque Canchaque (Huancabamba) donde se han registrado 58 familias, 103 géneros y 118 especies (Sagástegui et al. 2003); el inventario rápido de biodiversidad en tres relictos de bosques montanos de Ayabaca: Cuyas (Cerro Chacas), Aypate y El Toldo (Recharte et al. 2003); el reporte de 38 especies de orquídeas distribuidas en 11 géneros en el bosque de Cuyas, destacando los géneros Oncidium y Epidendrum con el mayor número de especies (Díaz 2003); el estudio florístico y ambiental del relicto de bosque Mijal, en Morropón, reportando 76 familias, 150 géneros y 216 especies, entre helechos y angiospermas, con presencia de endemismos y nuevos registros para el Perú (Sánchez & Grados 2007); el inventario florístico de fanerógamas del bosque de Cuyas, en Ayabaca, reportando 79 familias, 161 géneros y 211 especies (Farfán 2007) y el estudio etnobotánico en varias comunidades campesinas asentadas en el bosque de Cuyas (Ambulay 2008). Recientemente, otros bosques montanos han sido objeto de estudios florísticos y etnobotánicos como es el caso de los relictos de bosques montanos en Kañaris (Lambayeque) (Llatas-Quiroz & López-Meneses 2005) y La Oscurana (Cajamarca) (Juárez et al. 2005); sin embargo, en ninguno de los estudios realizados en los bosques montanos del norte y noroeste del Perú se determinó la estructura del bosque porque no se recogió información en parcelas o transectos específicos sobre número de individuos por área muestreada, diámetro a la altura del pecho (DAP), altura de planta, entre otros datos, que permitieran determinar la densidad relativa (DER), frecuencia relativa (FER), área basal (AB) e índice de valor de importancia (IVI).
Como es conocido, la diversidad y la composición florísticas son los atributos más importantes para diferenciar o caracterizar cada complejo y/o comunidad vegetal (Matteucci & Colma 1982), habiéndose definido varias clases de diversidad, en tanto que la composición florística de un bosque es determinada por el conjunto de especies de plantas que lo componen y es tradición medir considerando la frecuencia, abundancia y dominancia de las especies (Whittaker 1975). Es por ello que en cumplimiento del trabajo de campo para el proyecto "Evaluación Florística de los Bosques de Piura", liderado por la Asociación Chira-Sullana (Proyecto Binacional Perú-Ecuador), se ejecutaron actividades con el objetivo de valorar el estado actual de la riqueza y la estructura de la vegetación del bosque montano neotropical de Lanchurán, cuyos resultados constituyen una contribución al conocimiento de las condiciones naturales del bosque para facilitar la elaboración y ejecución de planes con fines de reforestación, conservación y utilización. Este trabajo constituye el primer esfuerzo realizado en el estudio de la estructura de un bosque montano en el norte y noroeste del Perú.
MATERIALES Y MÉTODOS
Área de estudio. El área de estudio se localizó en las coordenadas 4o38' Sur y 79o42' Oeste, entre 5-10 km del distrito de Ayabaca, capital de la provincia de Ayabaca, región Piura, abarcando una extensión aproximada de 1200 ha, en una elevación entre 2200 a 2900 m, caracterizándose por una extrema humedad ambiental ocasionada por las neblinas casi continuas. Geológicamente, el área de estudio corresponde a la época Media a Inferior del periodo Cretácico de la era Mesozoica (Km-vi-c), específicamente a las formaciones Volcánico Lancones, Célica y Grupo San Pedro (Ki-sp). La geomorfología corresponde a la región de la Sierra, subregión Altoandina y como gran paisaje a la vertiente montañosa moderadamente empinada y escarpada (Vs2-d). Los suelos son del orden Inceptisoles, suborden Tropepts, predominando los grupos Dystropepts (50%) y Ustrochrepts (50%) e hidrológicamente pertenece a la subcuenca del río Quiroz (Proyecto Binacional Catamayo-Chira 2006).
De acuerdo con la clasificación de zonas de vida de Holdridge (1966) y al Mapa Ecológico del INRENA (1995), el área de estudio correspondió a la zona de vida bosque húmedo Montano Bajo (bh-MB), bioclimática Temperado Húmedo (Te-h) y pluviométrica Subhúmeda y es un área natural potencial para la conservación, integrando el Bosque protector Ayabaca - Cerro Chacas con 21 134.06 ha, (Proyecto Binacional Catamayo-Chira 2006).
Se ejecutaron levantamientos en dos sitios, Los Molinos, con aproximadamente 200 ha, corresponde a una quebrada profunda y muy húmeda, una pendiente de 40%, con grandes árboles de dosel de 15 - 20 m, pertenecientes a los estratos superior y medio, y arbustos de 7 - 12 m, con una profusa y variada vegetación herbácea; el segundo sitio de muestreo, denominado La Antena, con aproximadamente 1000 ha, corresponde a una planicie de leve a fuertemente ondulada y cubierta con una densa vegetación de árboles y arbustos no mayores de 4 m de altura.
La información climatológica de temperatura y precipitación fue proporcionada por la Estación Climatológica Principal Ayabaca (04°38' Sur, 79°43' Oeste y 2700 m de altitud), ubicada en el distrito de Ayabaca, provincia de Ayabaca y región Piura, encontrándose en el ámbito geográfico del bosque montano neotropical de Lanchurán. La temperatura máxima media anual (enero a diciembre del 2008) fue 16.9oC, variando entre 15.4oC, media de febrero y 18.7oC, media de septiembre y la temperatura mínima media anual fue 10.1oC, variando entre 10.5oC, media de enero y 9.2oC, media de diciembre. La precipitación media anual fue 119.2 mm, oscilando entre 0.6 mm en julio y 485.8 mm en febrero.
Muestreo. El trabajo de campo se realizó entre los meses de febrero a abril del 2008, siguiendo el método propuesto por Gentry (1982, 1995), con algunas modificaciones, para muestreos de plantas leñosas. Este método radica en que el censo se realiza en un área de 0.1 ha donde se establecen 10 transectos de 50 x 2 m, distribuidos al azar u ordenadamente con una distancia máxima de 20 m. Para el desarrollo de la investigación se eligieron zonas homogéneas en su fisonomía, propendiendo a la unidad de paisaje, donde se delimitaron 5 parcelas de 50 x 10 m ( 500 m2) cada una, a intervalos de 20 m en línea recta perpendicular a la base de la montaña, para un total de 2500 m2 ( 0.25 ha). En los levantamientos del sitio Los Molinos, se registraron todos los individuos leñosos de diámetro ≥ 2.5 cm a la altura del pecho (DAP = 1.3 m de la base), tomada como medida de área basal. En el sitio La Antena, debido a lo profusa e intrincada de la vegetación, el muestreo fue únicamente cualitativo. Las especies herbáceas se colectaron, tanto en las áreas de los transectos como próximas a éstos, sin seguir un patrón de muestreo definido. Todos los especímenes, fértiles y estériles, se colectaron y herborizaron para su correcta identificación en el laboratorio. El tratamiento taxonómico de las familias siguió los planteamientos filogenéticos del Grupo de Filogenia de las Angiospermas (APG 1998, 2003). Las muestras botánicas se encuentran depositadas en el Herbario de la Asociación Chira, Mallares (Sullana, Piura) (HACh), de reciente creación.
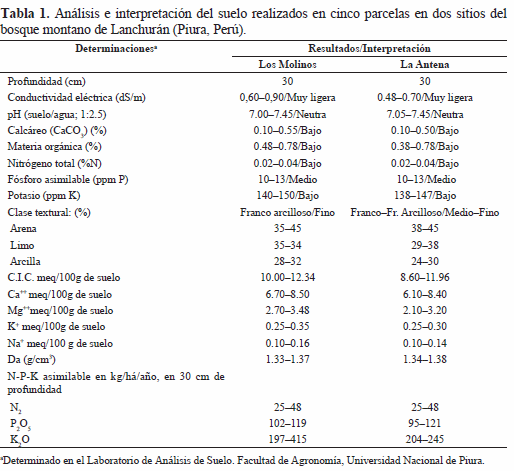
Se tomaron tres muestras de suelo, escogidas al azar, a 30 cm de profundidad en cada parcela muestreada de 500 m2, con un total de quince muestras por sitio de estudio. El análisis del suelo se realizó en el Laboratorio de Análisis de Suelo de la Facultad de Agronomía de la Universidad Nacional de Piura. En la determinación de la conductividad eléctrica, pH, potasio y cationes cambiables se siguieron las metodologías descritas por Jackson (1982) y en la determinación del fósforo, materia orgánica total, nitrógeno disponible, clase textural, carbonatos y la densidad aparente se siguieron las metodologías descritas por López & López (1990).
Procesamiento y análisis de la información. En la determinación de diferencias estadísticas significativas, entre las variables de suelo analizadas: pH, materia orgánica (%), fósforo asimilable (ppm P), capacidad de intercambio catiónico C.I.C (meq/100 g) y N-P-K asmilables (kg/ha/año), se realizó la prueba T de "student". En la tipificación de la abundancia o densidad de las especies presentes en los sitios de muestreo se utilizó la clasificación propuesta por Maleheiros & Rotta (1982) por la cual especies representadas por 1-6 individuos corresponden a especies raras o escasas, aquellas con 7-25 individuos se consideran poco abundantes y especies con más de 25 individuos se clasifican como abundantes. La estructura horizontal del bosque se describió a partir de la distribución del número de árboles por clase diamétrica (Hawley & Smith 1972). La Densidad Relativa (DER) se calculó: Número de individuos de una especie / número total de individuos x 100; la Frecuencia Relativa (FER): Frecuencia de la especie / frecuencia de todas las especies X 100; el Área Basal Relativa o (AB) o Cobertura Relativa (COB): AB de una especie / AB total x 100; el Índice de Valor de Importancia (IVI): Sumatoria de la DER, FER y AB relativa y Valor de Importancia para la Familia (VIF): Sumatoria de la Diversidad Relativa (Número de especies en una familia / número total de especies x 100), DER y Dominancia Relativa o Área Basal Relativa (Rangel & Velásquez 1997). En la medición de la diversidad alfa fue utilizado el índice de riqueza específica (IR) o índice de diversidad de Margalef, en tanto que en la medición de la diversidad beta se utilizó el índice o coeficiente de similitud (IS) de Sørensen. También se ha utilizado el índice de dominancia de Simpson y el índice de equidad de Shannon-Wiener (Magurran 1988, Moreno 2001).
RESULTADOS
Análisis del suelo
El análisis del suelo mostró características similares entre los sitios Los Molinos y La Antena, con excepción del K2O asimilable cuya concentración en el sitio Los Molinos duplicaba al determinado en el sitio La Antena. En ambos sitios de estudio, los suelos tienen un pH neutro (7.0 - 7.45) y las concentraciones de nutrientes disponibles son bajas: nitrógeno total (%N) 0.02 - 0.04, fósforo asimilable (ppm P) 10 - 13 y potasio (ppm K) 140 - 150; asimismo, la materia orgánica también es baja (0.48 - 0.78%) (Tabla 1). Realizada la prueba T de "student", entre las variables de suelo analizadas, suponiendo varianzas iguales, si bien se encontró ligeras diferencias, éstas no fueron estadísticamente significativas (p > 0.05).
Composición florística de Los Molinos
En los 2500 m2 inventariados en el sitio Los Molinos se encontraron 399 individuos, entre árboles y arbustos con un DAP≥ 2.5 cm, pertenecientes a 41 especies, 33 géneros y 25 familias (Tabla 2). A ello se suman 82 especies, 75 géneros y 48 familias de vegetación herbácea con diferentes hábitos de crecimiento (bejuco, hierba, hierba enredadera, hierba epífita, subarbusto, subarbusto trepador y sufrútice), lo que hizo un total de 123 especies, 108 géneros y 73 familias (Anexo 1).
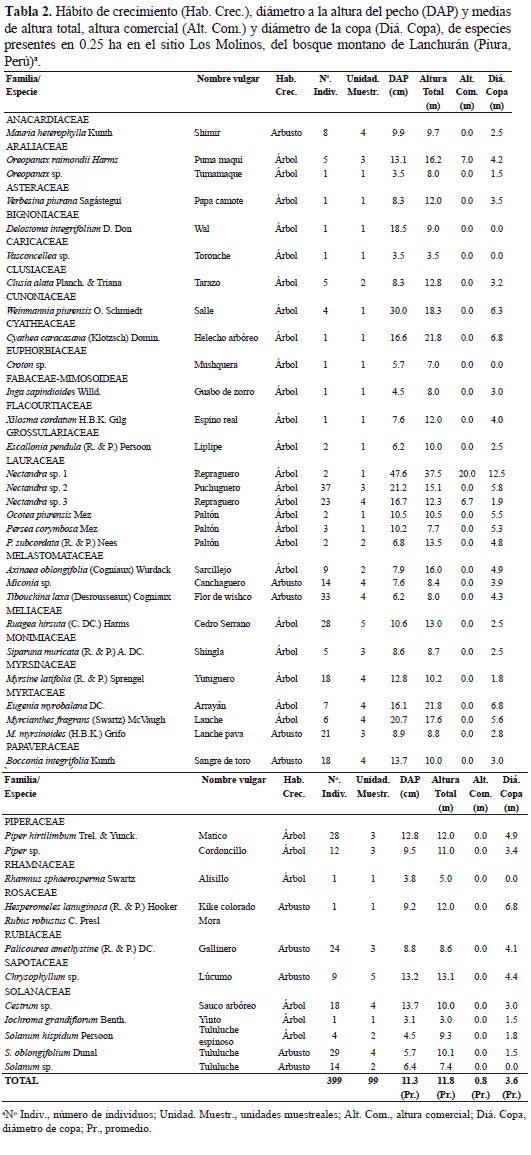

Estructura. De las 41 especies presentes, 30 son árboles (73.2%) y 11 arbustos (26.8%). Resulta importante destacar la presencia de seis especies de Lauraceae, agrupadas en los géneros Nectandra, Ocotea y Persea, haciendo un total de 69 individuos. Algunas especies como Delostoma integrifolia, Hesperomeles lanuginosa y Iochroma grandiflorum muestran un hábito de crecimiento arbustivo y arbóreo.
En la estratificación vertical se distingue el dosel superior conformado por la copa de los árboles dominantes, en especial individuos de la familia Lauraceae con un promedio de 16.1 m de altura; debajo de éstos árboles crecen arbustos leñosos, ramificados, y el estrato inferior está conformado por diversas especies herbáceas que crecen en los escasos espacios abiertos y otras son tolerantes a la sombra; entre estas especies herbáceas destacan Alternanthera porrigens, Bidens sp., Calceolaria sp., Commelina fasciculata, Salvia sp., Browalia americana, Alonsoa linearis, entre otras (Anexo 1). Los árboles de mayor tamaño son Nectandra sp. 1 con 37.5 m de altura seguido de Eugenia myrobalana y Weinmannia piurensis con 21.8 y 18.3 m, respectivamente, observándose una distribución altimétrica ligeramente en forma de "J" invertida (Tabla 3). La altura promedio de la vegetación es 11.8 m. La altura fustal es 0.8 m y las copas de los árboles no son muy amplias, alcanzando un diámetro promedio de 3.6 m, destacando Nectandra sp. 1, E. myrobalana y W. piurensis con 12.5, 6.8 y 6.3 m, respectivamente. Dos especies, Chrysophyllum sp. y Ruagea hirsuta se inventariaron en las cinco parcelas muestreadas en el sitio Los Molinos, mientras que diez especies en cuatro parcelas, siete en tres parcelas, seis en dos parcelas y 16 especies en solamente una parcela.
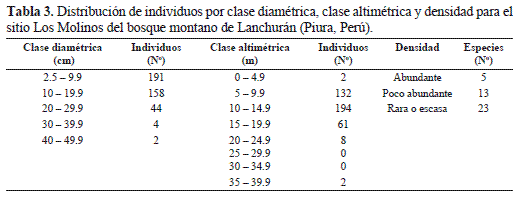
Densidad. Las especies con mayor número de individuos en 0.25 ha fueron: Nectandra sp. 2 con 37 individuos, seguido de Tibouchina laxa con 33, Solanum oblongifolium con 29 y Ruagea hirsuta y Piper hirtilimbum con 28 individuos, respectivamente, haciendo un total de cinco especies clasificadas como abundantes, en tanto que trece especies se consideran poco abundantes y 23 especies como raras o escasas (Tabla 3). Con excepción de T. laxa y S. oblongifolium, que tienen porte arbustivo, el resto son árboles de porte mediano a alto. Debido al crecimiento abundante de arbustos, lianas y epífitas la visibilidad en el bosque no es mayor de diez metros.
Área basal. El mayor DAP correspondió a Nectandra sp. 1 con 47.6 cm seguido de Weinmannia piurensis y Bocconia integrifolia con 30.9 y 22.6 cm, respectivamente, aunque B. integrifolia y Nectandra sp. 1 apenas registraron uno y dos individuos, respectivamente. Se observó una distribución diamétrica en forma de "J" invertida de todas las especies, la que es característica de los bosques tropicales, lo cual indica que la mayoría de individuos en los transectos estuvieron en la clase I (2.5 - 9.9 cm DAP) y clase II (10- 19.9 cm DAP) y a medida que aumentan los diámetros va disminuyendo la cantidad de individuos encontrados (Tabla 3). El valor acumulado de área basal es de 89.23 m2. La mayor cobertura relativa la tuvo Nectandra sp. 1 (19.79), seguido de Nectandra sp. 2 (14.38), Myrsine latifolia (9.11), Chrysophyllum sp. (8.45) y Myrcianthes fragrans (6.09). Las demás especies presentan valores bajos de cobertura relativa. En cuanto a familias destacaron Lauraceae (38.90), Myrtaceae (9.73), Myrsinacea (9.11) y Sapotaceae (8.45), con la mayor cobertura relativa (Tabla 4).
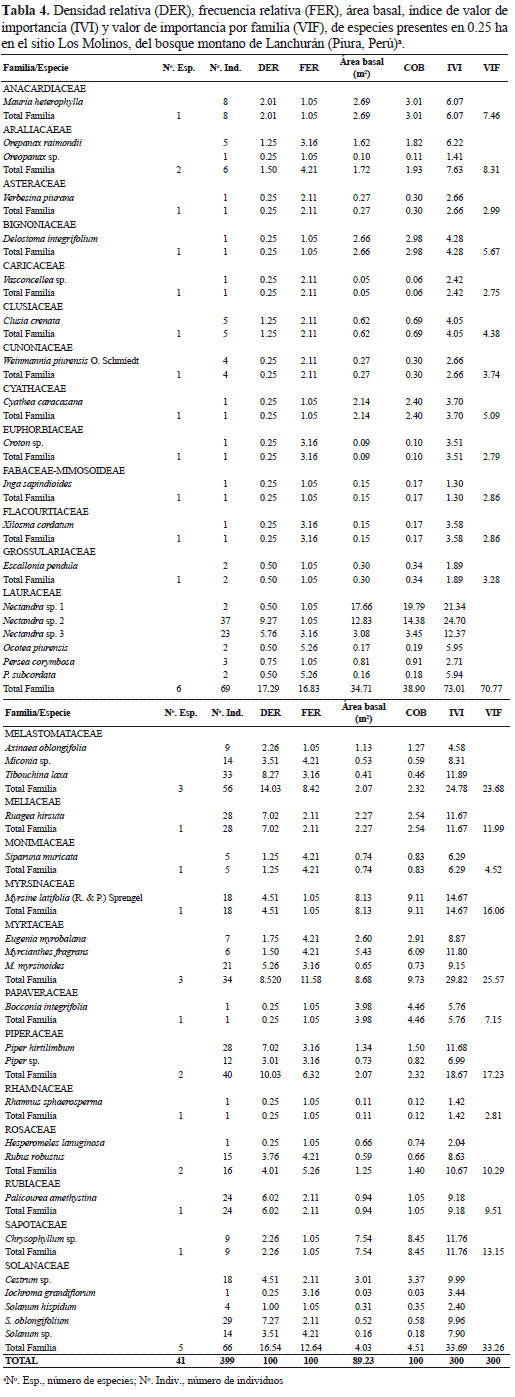
Índice de Valor de Importancia por especie (IVI). De las seis especies que presentan los mayores valores de importancia, cinco son árboles, Nectandra sp. 2 (24.70), Nectandra sp. 1 (21.34), Myrsine latifolia (14.67), Chrysophyllum sp. (11.76) y Piper hirtilimbum (11.68) y únicamente Myrcianthes fragrans (11.80), que es un arbusto. En cuanto a familias destacaron Lauraceae (73.01), Solanaceae (33.69), Myrtaceae (29.02) y Melastomataceae (24.78) con el mayor valor de importancia (Tabla 4).
Valor de Importancia por Familia (VIF). Las familias con mayor importancia ecológica fueron: Lauraceae (70.77), Solanaceae (33.26), Myrtaceae (25.57) y Melastomataceae, con valores similares al IVI (Tabla 4).
Composición florística de La Antena
En el sitio La Antena, en 2500 m2 muestreados, entre árboles y arbustos, se encontraron 86 especies, 67 géneros y 41 familias, sin inventariarse el número de individuos. Las especies arbóreas son 42 (48.8%) y las especies arbustivas 44 (51.2%). El mayor número de especies correspondió a las familias Asteraceae, Melastomataceae, Solanaceae y Lauraceae con 13 (15.1%), 9 (10.5%), 9 (10.5%) y 5 (5.8%), respectivamente, destacando los géneros Miconia y Solanum con 6 especies, respectivamente. La vegetación de árboles y arbustos se detalla en el Anexo 2.
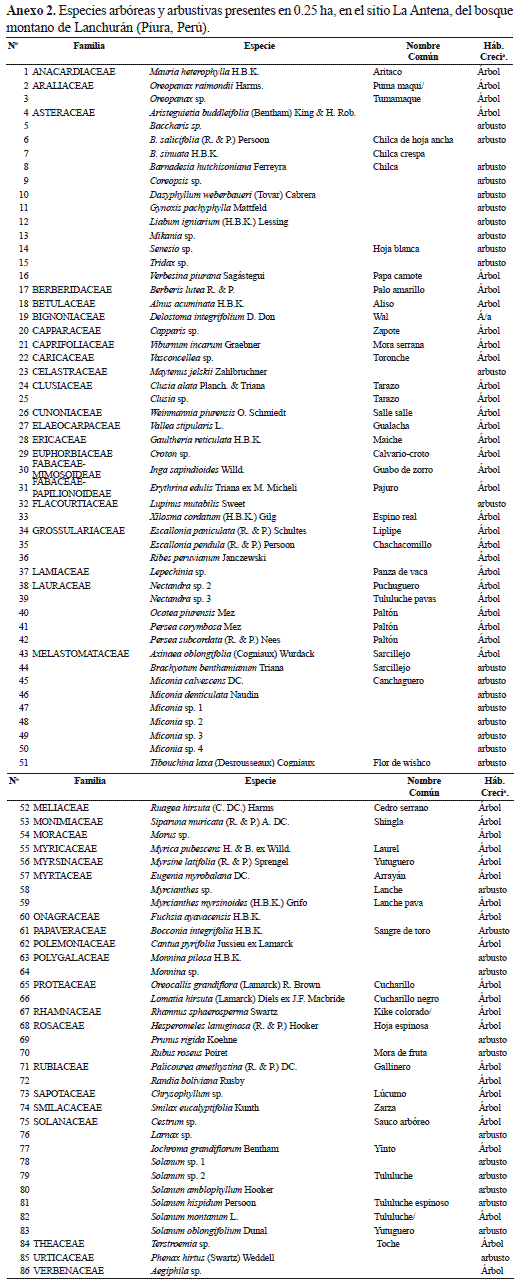
Si se compara el número de especies, géneros y familias, solamente entre árboles y arbustos, entre los sitios Los Molinos y La Antena, se comprueba que La Antena tiene 50 especies (58.1%), 37 géneros (55.2%) y 19 familias (46.3%) que no se encontró en Los Molinos.
Índice de Riqueza (IR) e Índice de similitud (IS)
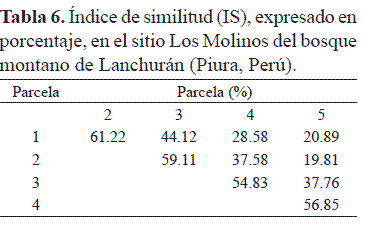
En la tabla 5, en el sitio Los Molinos, el IR específico, que expresa el número total de especies obtenido por un censo de la comunidad, indicó que el IR se incrementó desde la parcela 1 con IR = 3.60 (17 especies) hasta la parcela 4 con IR = 5.88 (26 especies), decayendo significativamente en la parcela 5 con IR = 2.99 (13 especies); el índice de equidad de Shannon-Wiener, que expresa la uniformidad de los valores de importancia a través de todas las especies de la muestra, fue mayor en la parcela 4 con 2.92 y menor en la parcela 5 con 2.16 y el índice de dominancia de Simpson, que considera la representatividad de las especies con mayor valor de importancia sin evaluar la contribución del resto de las especies, fue también mayor en la parcela 5 con 0.94 y menor en la parcela 5 con 0.87. En la tabla 6, el índice de similitud florística, que relaciona el número de especies en común con la media aritmética de las especies en las diferentes parcelas muestreadas en el sitio Los Molinos, indicó que ocurre una mayor similitud o afinidad entre las parcelas P1-P2 y P2-P3 con 61.2 y 59.1%, respectivamente, y entre P3-P4 y P4-P5 con 54.8 y 56.9, respectivamente, en tanto que los valores más bajos de similitud se presentó entre las parcelas P2-P5 y P1-P5 con 19,8 y 20.9%, respectivamente.

DISCUSIÓN
Los resultados en el análisis de suelo, indicaron una ligera diferencia, aunque no estadísticamente significativa, tanto en materia orgánica como en nutrientes, más alta en el sitio Los Molinos respecto del sitio La Antena; se esperaba una mayor similitud debido a su proximidad, formación geológica y estar expuestos a los mismos procesos de erosión y degradación. Aún así, la fuerte meteorización química debido a su naturaleza litológica, la elevada humedad y alta precipitación, en combinación con variaciones significativas de temperatura, lo que provoca un lavado de nutrientes, dio como resultado un suelo pobre en nutrientes, tal como se ha reportado como característico para este tipo de bosques (Loza 2008, Loza et al. 2010). La importancia de la calidad del suelo estaría relacionada con la diversidad vegetal pues algunos estudios han planteado que el número de especies aumenta en proporción inversa con la calidad del suelo (IDEADE 1996), lo que se correlacionó con los resultados obtenidos por León et al. (2009) donde el sitio con mayor contenido de materia orgánica en el suelo y nutrientes presentó menos especies; contrariamente, sitios con alta diversidad vegetal ocurrió en suelos con los más bajos contenidos de materia orgánica y bajas concentraciones de elementos que son importantes para la nutrición vegetal. En este estudio, el sitio La Antena, con menor contenido de materia orgánica en el suelo y nutrientes, presentó una mayor diversidad vegetal en árboles y arbustos, respecto al sitio Los Molinos. Estos resultados, y los obtenidos por León et al. (2009), apoyarían la hipótesis según la cual la riqueza de especies varía inversamente con la calidad de los suelos, por lo tanto, debería ser evaluada puntualmente en futuros estudios, para obtener resultados contundentes con relevancia estadística. Por otro lado, la presencia de árboles de mayor tamaño y diámetro, en el sitio Los Molinos, coincidiría con el mayor nivel de fertilidad, tanto en materia orgánica y los niveles de potasio y fósforo asimilables (Kennard 2001); sin embargo, no es posible conciliar esta observación con los resultados obtenidos en el sitio La Antena puesto que en éste sitio los individuos no eran mayores de 4 m de altura.
Observaciones previas realizadas en el bosque montano de Cuyas (Ayabaca) y otros bosques, próximos a Lanchurán (Farfán 2007, Ambulay 2008), informaciones recogidas de los habitantes del lugar y la presencia de un gran número de tocones y restos de árboles caídos, permiten asegurar que la cobertura vegetal presente en los sitios de muestreo ha sufrido una dramática alteración en su estructura y composición florística originales, lo que fue más significativo en el sitio La Antena que en el sitio Los Molinos; sin embargo, su condición actual aún muestra la composición típica de los bosques montanos del norte y noroeste del Perú (Sagástegui et al. 2003, Llatas-Quiróz & López-Meneses 2005, Arroyo et al. 2008, Sánchez & Grados 2007).
La mayor proporción de árboles de troncos rectos y altos correspondió al sitio Los Molinos, en tanto que árboles pequeños y arbustos correspondió al sitio La Antena, aunque en las áreas abiertas o bordes de caminos y trochas, en el sitio Los Molinos, la vegetación estaba integraba por hierbas, subfrútices, lianas y bejucos. Se ha señalado que los bosques montanos húmedos de las vertientes noroccidentales del Perú fueron en un tiempo continuos, pero a partir de 1940 se inició su intervención y destrucción acelerada por la actividad antrópica (Arroyo et al. 2008), quedando reducidos a no más de 23 relictos de bosques (Weigend et al. 2005), de allí la necesidad de plantear aspectos cruciales para alcanzar la conservación de estos bosques remanentes: mostrar su gran y compleja biodiversidad, los grados de endemismo, el valor económico potencial para la región y su papel como áreas de captación de agua.
La distribución diamétrica y altimétrica de todas las especies en el sitio Los Molinos, fue en forma de "J" invertida. Esta tendencia de distribución podría estar reflejando la característica susecional del bosque debido a las perturbaciones constantes y selectivas que se han realizado por los habitantes cercanos a la zona, manteniéndolo en una sucesión secundaria. La inclusión de individuos con altura de planta < 4.9 m y DAP < 9.9 cm otorga un cabal conocimiento sobre el proceso de regeneración de las especies presentes en las clases diamétricas y altimétricas mayores, lo que es pertinente su ejecución, tal como ha sido recomendado en un estudio reciente sobre estructura y diversidad florística de dos bosques naturales en el Cauca, Colombia (García et al. 2010).
La flora presente puede considerarse baja, para los dos sitios estudiados, donde en el sitio Los Molinos, entre árboles, arbustos y herbáceas se registraron 123 especies, 108 géneros y 73 familias; en tanto que en el sitio La Antena se registraron, entre árboles y arbustos, 86 especies, 67 géneros y 41 familias. En relación al sitio Los Molinos, éstos resultados son muy similares a los reportados para el bosque montano de Canchaque (Huancabamba) donde se registró 118 especies, 103 géneros y 58 familias (Sagástegui et al. 2003); sin embargo, éste bosque es quizás uno de los más depredados de todos los bosques montanos del norte del Perú, razón de su baja composición florística. Para otros bosques montanos como el de Cuyas (Ayabaca) (Farfán 2007) y Mijal (Morropón) (Sánchez & Grados 2007) se reportaron datos similares como 211 y 216 especies, 161 y 150 géneros y 79 y 76 familias, respectivamente. Toda esta información, sin embargo, resulta dramáticamente inferior al ser comparada con lo estimado para el bosque montano La Oscurana (San Miguel de Pallaques) con 258 especies, 169 géneros y 85 familias (Juárez et al. 2005), y el bosque montano de Cachil (Gran Chimú) con 400 especies, 206 géneros y 97 familias (Arroyo et al. 2008), lo que indica que el bosque montano de Lanchurán está en proceso de perder irremediablemente su composición florística si no se toman medidas urgentes de conservación.
Un análisis comparativo sobre las especies representativas de los relictos de bosques montanos en el norte y noroeste del Perú reveló algunas peculiaridades sobre la ausencia o presencia de determinadas especies en el bosque montano de Lanchurán. Así, se tiene la presencia en Lanchurán del "helecho arbóreo" Cyatea caracasana, especie que no se reportó para el bosque de Cuyas (Ambulay 2008), pero sí para el bosque de Kañaris, que además registró a Nephelia, otro helecho arbóreo del lugar (Llatas-Quiróz & López-Meneses 2005); ambas especies constituyen soportes para varias epífitas, principalmente orquídeas, considerándoseles amenazadas. Entre las gimnospermas, no se encontró en Lanchurán el "olivo", "romerillo" o "pino andino" Podocarpus oleifolius, una de las especies más amenazadas de los relictos de bosques montanos (Vicuña-Miñano 2005), y tampoco se le reportó en Cuyas (Ambulay, 2008), pero sí en Cachil (Arroyo et al. 2008) y Kañaris (Llatas-Quiróz & López-Meneses 2005). Entre las monocotiledóneas, no se encontró en Lanchurán la "palma de cera" Ceroxylon andicola y tampoco en Cuyas (Farfán 2007, Ambulay 2008) aunque todavía se encuentran varios individuos en Kañaris (Llatas-Quiróz & López-Meneses 2005) y en el Parque Nacional de Cutervo (Sagástegui et al. 2003), la primera área natural protegida establecida en el Perú y paradójicamente la más depredada de los bosques montanos. En lo referente a las dicotiledóneas, no se encontró en Lanchurán especies arbóreas tan representativas como la "cascarilla Cinchona ledgeriana, "nogal" Juglands neotropica, "higuerón" Ficus sp. y "cedro" Cedrela odorata, las que tampoco se reportan para Cuyas (Farfán 2007, Ambulay 2008), pero sí para Kañaris (Llatas-Quiróz & López-Meneses 2005). Estos resultados se correlacionan con lo ya afirmado de que el bosque de Lanchurán está perdiendo su composición florística, igual como sucede con el bosque de Cuyas.
El inventario de las especies leñosas y herbáceas en el bosque montano de Lanchurán, no obstante las variaciones significativas en su flora con relación a otros bosques montanos del norte y noroeste del Perú, demuestra poseer aún un gran potencial fitogenético, si se tiene en cuenta la presencia de varios géneros como recursos maderables: Escallonia, Ruagea, Oreocallis, Myrsine, Nectandra, Alnus, Ocotea y Persea; alimenticios: Chrysophyllum, Passiflora y Myrcianthes; medicinales: Croton, Clusia, Piper, Maytenus y Stevia; cosméticos: Cestrum y Iochroma; ornamentales: Bomarea, Nasa, Fuchsia, Palicourea, Calceolaria, Epidendrum, Verbesina, Telipogon, Begonia y Tillandsia e industrial tinctórea: Bocconia. La mayoría de estos géneros han sido considerados importantes económicamente en una evaluación florística realizada en los relictos de bosques de Cachil (Arroyo et al. 2008) y de Cuyas (Ambulay 2008). Asimismo, si se comparan los resultados que se presentan con los reportados por Llatas-Quiróz & López-Meneses (2005), para el relicto de bosque de Kañaris, se observa que algunas especies del bosque húmedo Montano Bajo Tropical (bh-MBT) ( 1750 a 2250 m) como el "lanche" (Myrcianthes), "aliso" (Alnus), "congona" (Peperomia), "matico" (Piper), "zarzamora" (Rubus), "huaba shilimba" (Inga), "añasquero" (Siparuna) y "chotaquiro" (Miconia), así como algunas especies del bosque muy húmedo Montano Bajo Tropical (bmh-MBT) ( 2250 a 3000 m) como Nectandra, Ocotea, Persea, Weinmannia y los helechos arborescentes del género Cyathea, también se encuentran en el bosque montano de Lanchurán, lo cual aún les permite mantener la consistencia en la composición taxonómica propia de los bosques montanos de los Andes Tropicales.
En el sitio La Antena, conformado por un sotobosque muy denso y casi impenetrable de especies arbóreas y arbustivas, no mayor de 4 m de altura, si bien se reporta la mayoría de las especies maderables más representativas registradas en el sito Los Molinos, la competencia con otros árboles y arbustos dificultaría el que puedan prosperar y constituirse en especies dominantes. Llatas-Quiróz & López-Mesones (2005) reportaron que una parte del relicto de bosque montano de Kañaris está formado por un sotobosque impenetrable donde predominan especies de las familias Rubiaceae, Ericaceae, Araceae, entre otras, ocurriendo algo similar con el bosque de Lanchurán, donde aparte de las familias mencionadas se puede señalar a Melastomataceae, Solanaceae y Asteraceae.
Las diferencias encontradas entre los bosques montanos del norte y noroeste del Perú, en términos de diversidad de especies, podrían explicarse por las características particulares del sitio como la historia de uso, el tipo de intensidad de las perturbaciones pasadas, la topografía del lugar y la fauna existente; estos factores podrían resultar claves en el desarrollo de la dinámica forestal, como la formación de claros, regeneración natural, dispersión, entre otros, y verse reflejados en características del bosque como la riqueza y la composición florística, tal como ha sido señalado para explicar la dinámica forestal de robledales de la región norte de la cordillera central de Colombia (León et al. 2009). En efecto, las características geomorfológicas, climáticas (temperatura y pluviosidad) e hidrográficas originales del bosque montano neotropical de Lanchurán, así como la similitud en los análisis de suelo y la poca variación en la latitud, longitud y altitud, entre los sitios Los Molinos y La Antena, deberían determinar la ocurrencia de una diversidad vegetal y estructura bastante parecida; sin embargo, ello no fue así atribuyéndose a que ambos sitios han estado expuestos a diferentes presiones de deforestación, resultando muy intensa en el sitio La Antena, como lo reportaron las observaciones preliminares de Farfán (2007) y Ambulay (2008), para los bosques montanos de los alrededores de Lanchurán. En este sitio, la proximidad a la ciudad de Ayabaca y la presencia de una carretera afirmada que conduce a una estación de televisión, la que bordea gran parte del bosque, han determinado una deforestación intensa, hecho que viene sucediendo recientemente puesto que los nuevos especímenes, por regeneración natural o que han rebrotado, no superan los 4 m de altura, pero en compensación exhiben una intrincada distribución lo que hizo imposible el muestreo cuantitativo. Diferente resultó el bosque en el sitio Los Molinos donde la deforestación ha resultado menor, lo que se atribuye a que se extiende en una quebrada profunda, de difícil acceso, y de alguna manera protegido por sus propietarios.
No fue posible realizar comparaciones sobre el AB, IVI y VIF, entre los resultados que se presentan con los de otros bosques montanos del Perú, puesto que los escasos trabajos realizados al respecto fueron solamente de levantamiento florístico (Sagástegui et al. 2003, Juárez et al. 2005, Farfán 2007, Sánchéz & Grados 2007), distribución altitudinal (Llatas-Quiróz & López-Mesones 2005), etnobotánicos (Ambulay 2008) o con fines de conservación (Weigend et al. 2005, Sánchez & Grados 2007, Arroyo et al. 2008) y en ningún caso se evaluó la estructura de las poblaciones que aún persisten; sin embargo, si se considera el AB obtenido en el sitio Los Molinos, que fue 89.23 m2, con otros bosques montanos de países vecinos, se comprueba que resultó significativamente alto respecto al AB = 37.5 m2 en el bosque montano boliviano Madidi, donde se evaluó 15 sitios de 0.1 ha (20 x 50 m) (Loza et al. 2010) y el AB = 29.4 m2 en el bosque montano de la Cordillera Mosetenes en Bolivia, donde se evaluó cuatro parcelas de 0.1 ha, tres de 50 x 20 y una de 100 x 10 (Macía & Fuertes 2008).
Para entender los resultados obtenidos con el índice de riqueza (IR), el índice de equidad (IE), el índice de dominancia (ID) y el índice de similitud (IS) conviene recalcar que el transecto de muestreo, con 40% de pendiente, fue establecido de manera perpendicular a la base de la montaña, con parcelas de 50 x 10 m e intervalos de 20 m, por lo tanto, entre la parcela 1 y la parcela 5 se estableció una distancia de 330 m, con la parcela 3 a 2400 m y la parcela 5 a 2500 m de elevación. En lo referente al índice de riqueza, en el sitio Los Molinos, los resultados expresaron que ésta se incrementó conforme se ascendió en la posición de la parcela hasta alcanzar el mayor valor con IR = 5.88, en la parcela P4, para luego decaer dramáticamente en la parcela P5 con IR = 2.99. Algunos autores han señalado que la riqueza de especies tiende a disminuir conforme se asciende en altitud, tal como fue comprobado que la diversidad florística de los bosques montanos a altitudes superiores a los 1500 m disminuye linealmente (Gentry 1995). Asimismo, en el bosque montano de la Cordillera Mosetenes, en Bolivia, las parcelas ubicadas entre 1240 - 1290 m de altitud mostraron una mayor riqueza de especies en comparación con la parcela ubicada a 1560 m de altura que mostró la menor riqueza de especies (Macía & Fuertes 2008). Una tendencia similar se observó tanto para el índice de equidad como para el índice de dominancia.
En lo referente al índice de similitud, en el sitio Los Molinos, los resultados expresaron más de 50% de similitud florística entre las parcelas ubicadas en la parte inferior del transecto (P1-P2 y P2-P3) y entre las parcelas de la parte superior del transecto (P3-P4 y P4-P5), en cambio la similitud florística es alrededor de 20% entre las parcelas extremas del transecto (P1-P5 y P2-P5), por lo tanto, la distancia de separación entre las parcelas puede resultar determinante para explicar la escasa similitud florística entre las parcelas ubicadas en los extremos del transecto. Loza et al. (2010), estudiando la variación de la diversidad y composición florística en relación a la elevación en el bosque montano boliviano Madidi ( 1000 a 1600 m), concluyeron que la diversidad florística, calculada por los índices de diversidad alfa y beta, no fue explicada por la elevación, en tanto que la composición florística, determinada mediante el análisis de ordenación NMS (non-metric multidimensional scaling), sí cambió en relación a la elevación. Un estudio similar, aunque más complejo puesto que involucra condiciones climáticas, análisis de suelo y características topográficas, ha sido realizado recientemente por Homeier et al. (2010) en los bosques montanos del Ecuador. Los resultados que se presentan abren una puerta para la investigación sobre la composición florística de los bosques montanos del norte y noroeste del Perú en relación con otros factores como, por ejemplo, la elevación del terreno.
CONCLUSIONES
Las características del suelo fueron similares entre los sitios Los Molinos y La Antena con excepción del K2O asimilable con una concentración en el sitio Los Molinos duplicando al determinado en el sitio La Antena.
En el sitio Los Molinos se encontraron 399 individuos, entre árboles y arbustos con un DAP ≥ 2.5 cm, correspondiendo a 41 especies, 33 géneros y 25 familias, a lo que se sumaron 82 especies, 75 géneros y 48 familias de vegetación herbácea con diferentes hábitos de crecimiento.
En el sitio La Antena, se registraron, entre árboles y arbustos, 86 especies, 67 géneros y 41 familias.
Los dos sitios estudiados, Los Molinos y La Antena, difieren en su composición florística y estructura, no obstante presentar características biofísicas similares y que en ambos casos se encuentran depredados. Las diferencias encontradas se atribuyen, a los diferentes factores que interactúan en cada sitio como la historia de uso, la intensidad y duración de las perturbaciones antrópicas pasadas y presentes, la topografía, calidad del suelo, la fauna y el clima local, que se ven reflejados en los parámetros estructurales de la vegetación.
AGRADECIMIENTOS
A la Xunta de Galicia (España) y al Proyecto Binacional Catamayo - Chira, por el financiamiento de la investigación, a Jorge Chanamé Céspedes, profesor de la Universidad Nacional Pedro Ruiz Gallo de Lambayeque, por su contribución en el análisis estadístico, a José Ayasta, por su colaboración en la identificación de algunas muestras botánicas y a Alexander Huamán y Jeannine Córdova, por el apoyo técnico en la preparación del manuscrito.
LITERATURA CITADA
1. AMBULAY, I.M. 2008. Etnobotánica en las comunidades campesinas Cuyas - Cuchayo, Joras y Suyupampa del bosque de Cuyas, Ayabaca - Piura, 2006. Tesis Lic. Biología. Universidad Nacional de Piura, Perú. [ Links ]
2. APG (ANGIOSPERM PHYLOGENY GROUP). 1998. An ordinal classification for the families of flowering plants. Annals of Missouri Botanical Garden 85:531-553. [ Links ]
3. APG (ANGIOSPERM PHYLOGENY GROUP). 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141:399-436. [ Links ]
4. ARROYO, S.J., E.F. RODRÍGUEZ, S. LEIVA, M. ZAPATA & M. MORA. 2008. El bosque relicto de Cachil (Provincia Gran Chimú, Departamento La Libertad, Perú), un ecosistema que necesita planes de conservación urgente. Arnaldoa 15: 289-296. [ Links ]
5. BRACK, A. & C. MENDIOLA. 2004. Ecología del Perú. PNUD. Asociación Editorial Bruño. Lima. 495 p. [ Links ]
6. BRUIJNZEEL, L.A. 2001. Hydrology of tropical montane cloud forest: a reassessment. Land Use and Water Resources Research 1: 1.1-1.18. [ Links ]
7. CHURCHIL, I.S.P., H. BALSLEV, E. FORERO & J. LUTEYN (eds.). 1995. Biodiversity and Conservation of Neotropical Montane Forest. The New York Botanical Garden, Nueva York. 702 p. [ Links ]
8. CONDIT, R., P.S. ASHTON, P. BAKER, S. BUNYAVEJCHEWIN, S. GUNATILLEKE, N. GUNATILLEKE, S.P. HUBBELL, R.B. FOSTER, A. ITOH, J.V. LA FRANKIE, H-S. LEE, E. LOSOS, N. MANOKARAN, R. SUKUMAR & T. YAMAKURA. 2000. Spatial patterns in the distribution of tropical tree species. Science 288: 1414-1418 [ Links ]
9. CUESTA, F., M. PERALVO & N. VALAREZO. 2009. Los bosques montanos de los Andes Tropicales. Una evaluación regional de su estado de conservación y de su vulnerabilidad al efecto del cambio climático. Serie Investigación y Sistematización # 5. Programa Regional ECOBONA - INTERCOOPERATION, Quito.Regional [ Links ]
10. DÍAZ, A. 2003. Orquídeas del bosque de Cuyas. Tesis Lic. Biología. Universidad Nacional de Piura, Perú. [ Links ]
11. DILLON, M.O. 1994. Bosques húmedos del norte del Perú. Arnaldoa 2: 29-42. [ Links ]
12. DILLON, M.O., A. SAGÁSTEGUI, I. SÁNCHEZ, S. LLATAS & N. HENSOLD. 1995. Floristic inventory and biogeographic analysis of montane forests in north western Peru. En: S.P. Churchill, H. Balslev, E. Forero & J.L. Luteyn (eds.), Biodiversity and Conservation of Neotropical Montane Forests: 251-269. The New York Botanical Garden. Nueva York. [ Links ]
13. FARFÁN, M.S. 2007. Fanerógamas del bosque de Cuyas, Ayabaca, Enero - Julio 2006. Tesis Lic. Biología. Universidad Nacional de Piura, Perú. [ Links ]
14. GARCÍA, C., C. SUÁREZ & M. DAZA. 2010. Estructura y diversidad florística de dos bosques naturales (Buenos Aires, Dpto Cauca, Colombia). Revista Biotecnología en el Sector Agropecuario y Agroindustrial (Universidad del Cauca) 8: 74-82. [ Links ]
15. GENTRY, A.H. 1982. Patterns of neotropical diversity. Evolutionary Biology 15: 1-84. [ Links ]
16. GENTRY, A.H. 1989. Northwest South America (Colombia, Ecuador and Peru. Págs. En: D.G Campbell & H.D. Hammond (eds.), Floristic Inventory of Tropical Countries: 392-400. The New York Botanical Garden, Nueva York. [ Links ]
17. GENTRY, A.H. 1995. Patterns of diversity and floristic composition in Neotropical Montane Forests. En: S.P. Churchill, H. Balslev, E. Forero & J.L. Luteyn (eds.), Biodiversity and Conservation of Neotropical Montane Forests: 103-126. The New York Botanical Garden, Nueva York. [ Links ]
18. HAMILTON, L.S. 2001. Una campaña por los bosques nublados. Ecosistemas únicos y valiosos en peligro. En: M. Kapelle & A.D. Brown (eds.), Bosques Nublados del Neotrópico. Instituto Nacional de Biodiversidad (INBio). Santo Domingo de Heredia. 698 p. [ Links ]
19. HAWLEY, R.C. & D.M. SMITH. 1972. Silvicultura Práctica. Omega, Barcelona. 544 p. [ Links ]
20. HENSOLD, N. 1999. Las angiospermas endémicas del Dpto. de Cajamarca, Perú. Arnaldoa 6: 141-184. [ Links ]
21. HOLDRIDGE, L. 1996. Ecología basada en Zonas de Vida. Instituto Interamericano de Cooperación para la Agricultura, San José, Costa Rica. [ Links ]
22. HOMEIER, J., S.W. BRECKLE, S. GÜNTER, R.T. ROLLENBECK & C. LEUSCHNER. 2010. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich ecuadorian montane rain forest. Biotropica 42: 140-148. [ Links ]
23. IDEADE. 1996. Evaluación de estatus ecosistémico y manejo de los bosques de Fagaceas (Quercus humboldtii Bonpl. y Trigonobalanus excelsus) en el norte de la cordillera oriental (Cundinamarca, Santander y Boyacá). Informe Final. Universidad Pontificia Bolivariana, Santa Fe de Bogotá, Colombia. [ Links ]
24. INRENA (INSTITUTO NACIONAL DE RECURSOS NATURALES). 1995. Mapa Ecológico del Perú. Guía Explicativa. Lima. [ Links ]
25. JACKSON, M.L. 1982. Análisis químico de suelos. Cuarta Edición. Ed. Omega, S.A., Barcelona. 662 pp. [ Links ]
26. JØRGENSEN, P.M. & C. ULLOA ULLOA, R. VALENCIA R. & J.E. MADSEN. 1995. A floristic analysis of the high Andes of Ecuador. En: S.P. Churchill, H. Balslev, E. Forero & J. L. Luteyn (eds.), Biodiversity and Conservation of the Neotropical Montane Forests: 221-237. The New York Botanical Garden, Nueva York. [ Links ]
27. JUÁREZ, A.M., J.E. AYASTA, R.P. AGUIRRE & E.F. RODRÍGUEZ. 2005. La Oscurana (Cajamarca), un bosque relicto más para conservar en las vertientes occidentales del norte del Perú. Revista Peruana de Biología 12: 289-298. [ Links ]
28. KAPELLE, M. & D. BROWN. 2001. Introducción a los bosques neotropicales. Una síntesis regional. En: M. Kappelle & D. Brown (eds.), Patrones de Diversidad y Composición Florística en los Bosques de las Montañas Neotropicales: 26-40. Bosques Nublados del Neotropico, 1ra Edición. INBIO-FUA-UICN, Santo Domingo de Heredia. [ Links ]
29. KENNARD, K.K. 2001. Secondary forest succession in a tropical dry forest: patterns of development across a 50-year chronosequence in lowland Bolivia. Journal of Tropical Ecology 18: 53-66. [ Links ]
30. LLATAS-QUIROZ, S. & M. LÓPEZ-MENESES. 2005. Bosques montanos-relictos en Kañaris (Lambayeque, Perú). Revista Peruana de Biología 12: 299-308. [ Links ]
31. LÓPEZ, J. & J. LÓPEZ. 1990. El diagnóstico de suelos y plantas: Métodos de campo y laboratorio. Tercera Edición. Ed. Mundi-Prensa, Madrid. 363 pp. [ Links ]
32. LEÓN, J.D., G. VÉLEZ & A.P. YEPES. 2009. Estructura y composición florística de tres robledales en la región norte de la cordillera central de Colombia. Revista Biología Tropical 57: 1165-1182. [ Links ]
33. LÓPEZ, W. & A. DUQUE. 2010. Patrones de diversidad alfa en tres fragmentos de bosques montanos en la región norte de los Andes, Colombia. Revista de Biología Tropical 58: 483-498. [ Links ]
34. LOZA, I. 2008. Relación entre la composición florística con factores edáficos en un bosque pluviestacional montano húmedo (Parque Nacional Madidi La Paz Bolivia). Tesis de Licenciatura en Biología, Universidad Mayor de San Andrés, La Paz. 90 p. [ Links ]
35. LOZA, I., M. MORAES & P.M. JØRGENSEN. 2010. Variación de la diversidad y composición florística en relación a la elevación en un bosque montano boliviano (PNANMI Madidi). Ecología en Bolivia 45: 87-100. [ Links ]
36. MACÍA, M.J. & J. FUERTES. 2008. Composición florística y estructura de los árboles en un bosque tropical montano de la Cordillera Mosetenes, Bolivia. Revista Boliviana de Ecología y Conservación Ambiental 23: 1-14. [ Links ]
37. MAGURRAN, A.E. 1988. Ecological diversity and its measurement. Princeton University Press, Nueva Jersey, 179 p. [ Links ]
38. MALEHEIROS, D.E.O. & E. ROTTA. 1982. Levantamiento horizontal de uma mata de araucaria do primeiro planalto paranaense. Boletim de Pesquisa 4: 1-39 (Florianópolis). [ Links ]
39. MATTEUCCI, S.D. & A. COLMA. 1982. Metodología para el estudio de la vegetación. Monografías Científicas, Serie Biología No 22. Organización de los Estados Americanos, Washington D.C. 168 p. [ Links ]
40. MOSTACERO, J., F. MEJÍA & W. ZELADA. 2006. Dinámica poblacional del olivo, Podocarpus oleifolius en el bosque de Cachil (La Libertad-Perú), durante 2003-2005. Sciéndo 9(2): 51-56. [ Links ]
41. MORENO, C.E. 2001. Métodos para medir la biodiversidad. M&T - Manuales y Tesis SEA, vol. 1. CYTED, ORCYT/UNESCO & SEA, Zaragoza. [ Links ]
42. MYERS, N., R.A. MITTERMEIER, C.G. MITTERMEIER, G.A.B. DAFONSECA & J. KENT. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. [ Links ]
43. PRO NATURALEZA. 2004. Las áreas de conservación privada en el Perú. Un camino para involucrarse en la conservación de la naturaleza. Lima. [ Links ]
44. PROYECTO BINACIONAL CATAMAYO-CHIRA. 2006. Atlas Temático de la Cuenca Binacional Catamayo Chira. Edit. Proyecto Binacional de Ordenamiento, Manejo y Desarrollo de la Cuenca Catamayo Chira. 51 p. [ Links ]
45. RANGEL-CH., J.O. & A. VELÁSQUEZ. 1997. Métodos de estudio de la vegetación. En: J.O. Rangel, P. Lowy & M. Aguilar (eds). Colombia Diversidad Biótica II: 59-82. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. Bogotá D.C. [ Links ]
46. RECHARTE, J., L. ALBÁN, R. ARÉVALO, E. FLORES, L. HUERTA, M. ORELLANA, L. OSCANOA & P. SÁNCHEZ. 2003. El Grupo Páramos/Jalcas y Punas del Perú. Instituciones y Acciones en Beneficio de Comunidades y Ecosistemas Altoandinos. En: Memorias del Congreso Mundial de Páramos (Mayo de 2002): 785-811. Tomo II. Ministerio de Medio Ambiente, CAR, IDEAM & Conservación Internacional. Bogotá D.C. [ Links ]
47. SAGÁSTEGUI, A., I. SÁNCHEZ, M. ZAPATA & M.O. DILLON. 2003. Diversidad Florística del Norte del Perú. Tomo II. Bosques Montanos. Universidad Privada Antenor Orrego, Trujillo. Edit. Graficart. 305 p. [ Links ]
48. SÁNCHEZ, I. & N. GRADOS. 2007. Estudio florístico y ambiental del bosque releicto Mijal en la provincia de Morropón, Piura. Arnaldoa 14: 259-268. [ Links ]
49. TOBÓN, C. & F. ARROYAVE. 2007. Inputs by fog and horizontal precipitation to the paramo ecosystems and their contribution to the water balance. En: Proceedings Fourth International Conference on Frog Collection and Dew: 233-236. La Serena, Chile. [ Links ]
50. VICUÑA-MIÑANO, E.E. 2005. Las Podocarpáceas de los bosques montanos del noroccidente. Revista Peruana de Biología 12: 283-288. [ Links ]
51. WEIGEND, M. 2002. Observations on the biogeography of the Amotape-Huancabamba Zone in Northern Peru. En: K. Young, C. Ulloa, J.L. Luyeyn & S. Knapp (eds), Plant Evolution and Endemism in Andean South America: 38-54. The Botanical Review 68: 38-54. [ Links ]
52. WEIGEND, M., E. RODRÍGUEZ & C. ARANA. 2005. The relict forests of Northwest Peru and Southwest Ecuador. Revista Peruana de Biología 12: 185-194. [ Links ]
53. WHITTAKER, R.H. 1975. Communities and Ecosystems. Mac Millan Publishing. Nueva York. 385 p. [ Links ]

