










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.34 no.1 Bogotá Jan./June 2012
ALEJANDRA HURTADO
SELENE ESCOBAR
ALBA MARINA TORRES
INGE ARMBRECHT
Departamento de Biología, Universidad del Valle, Cali, Colombia. aleja8803@gmail.com
RESUMEN
La colonización de los potreros por especies vegetales diferentes a los pastos está limitada por la escasez de dispersores primarios y la poca disponibilidad de sitios adecuados para la germinación de las plantas. Las hormigas generalistas de potrero podrían promover la dispersión secundaria de árboles de interés para la rehabilitación ecológica mediante la manipulación y el transporte de semillas hacia sus nidos. En este estudio se examinó la necesidad de la escarificación mecánica para la germinación de semillas de tres especies de árboles de interés para la rehabilitación ecológica: Guazuma ulmifolia, Pithecellobium dulce y Senna spectabilis en condiciones de laboratorio. La escarificación potenció la germinación de G. ulmifolia (5% vs. 37%) y S. spectabilis (0% vs. 98.5%) y disminuyó la germinación de P. dulce (94% vs. 87%). Se concluye que la escarificación mecánica es necesaria para la germinación de S. spectabilis y G. ulmifolia. Con el objeto de explorar si las hormigas inciden en la germinación, en el laboratorio se ofrecieron semillas de S. spectabilis sin escarificar a colonias de la hormiga de fuego Solenopsis geminata. No se obtuvo germinación, lo que sugiere que esta especie de hormiga no realiza escarificación mecánica ni química a las semillas, pero tampoco hubo evidencia de daño físico a las semillas. Finalmente, se comparó la germinación de semillas escarificadas de S. spectabilis en suelo de nidos inactivos de S. geminata y en suelo adyacente al nido, en invernadero y campo. En invernadero la germinación fue mayor en suelo de nidos que en suelo adyacente (98% vs. 86%, respectivamente) pero en el campo sucedió lo contrario (24% vs. 43%, respectivamente). No se encontró evidencia que los cambios provocados al suelo por S. geminata favorezcan la germinación de S. spectabilis. Se recomienda evaluar la contribución del suelo de nidos en etapas post-germinación de establecimiento, supervivencia y crecimiento de plántulas.
Palabras clave. Germinación, hormigueros, Guazuma ulmifolia, Pithecellobium dulce, Senna spectabilis.
ABSTRACT
The colonization of pastures by plant species other than grasses is limited by the scarcity of primary seed dispersers and the reduced availability of suitable sites for seed germination. In pastures, generalist ants could promote secondary dispersal of trees of importance for ecological restoration by manipulating and transporting seeds to their nests. First, we evaluated in the laboratory the importance of mechanical scarification for seed germination of three tree species with potential for ecological restoration: Guazuma ulmifolia, Pithecellobium dulce and Senna spectabilis. Scarification enhanced germination of seeds of G. ulmifolia (from 5 to 37%) and S. spectabilis (from 0 to 98.5%) while decreased the germination of P. dulce (from 94 to 87%). We concluded that mechanical scarification is necessary for germination of S. spectabilis and G. ulmifolia. Then, to explore the effect of ants on seed germination, we offered non-scarified seeds of S. spectabilis to colonies of fire ant Solenopsis geminata in the laboratory. We did not observe germination with this treatment, suggesting that this ant species does not scarify mechanically or chemically the seeds, but there was no evidence of physical damage to the seeds. Finally, germination of scarified seeds of S. spectabilis in soil from S. geminata nests was compared with germination of scarified seeds in soil adjacent to the nests, both under greenhouse and field conditions. In greenhouse, germination was higher in nest soil than in adjacent soil (98% vs. 86%, respectively) whereas the opposite occurred in field (24% vs. 43%, respectively). There was no evidence that the changes in soil induced by nests of S.geminata favor the germination of S. spectabilis. We recommend assessing the contribution of soil from ant nests in post-germination stages of seedlings, their establishment, survival and growth.
Key words. Germination, ant nests, Guazuma ulmifolia, Pithecellobium dulce, Senna spectabilis.
Recibido: 16/06/2011
Aceptado: 12/03/2012
INTRODUCCIÓN
En América tropical, una gran proporción de la tierra se dedica a pasturas y el incremento de esta actividad se ha realizado, en la mayor parte de los casos, a costa de la reducción de ecosistemas naturales, especialmente bosques tropicales (Murgueitio & Ibrahim 2004). En Colombia se ha perdido la tercera parte de los bosques tropicales por debajo de los 1000 metros de elevación, todos caracterizados por altos niveles de riqueza y endemismo de especies, y en la actualidad, se estima que el país cuenta con 40,6 millones de hectáreas de tierra de pastoreo permanente y una tasa de deforestación anual de 300.000 hectáreas (Murgueitio et al. 2011).
La ganadería extensiva conlleva al establecimiento de múltiples hectáreas de pastura, frecuentemente desprovistas de cobertura arbórea y dominadas por una especie de gramínea, la cual es introducida en el caso de las pasturas mejoradas. Esta simplificación de la estructura vegetal a escala del paisaje, ocasiona el deterioro del suelo y una drástica reducción en la diversidad de flora y fauna, con la consecuente pérdida de los servicios ambientales asociados a ella (Folgarait 1998). Por ejemplo, en la zona de bosque seco tropical de los valles interandinos, existen especies de árboles endémicos que pueden jugar papeles predominantes en los intentos de mejorar la resiliencia y la sustentabilidad de los sistemas ganaderos. Estas plantas pueden tener varios propósitos adicionales a la actividad pecuaria, como son la producción de madera y frutos, la contribución a un microclima más benigno, la oferta de hábitat para la fauna silvestre (i.e. dispersores, polinizadores, parasitoides), la regulación hídrica en cuencas hidrográficas y una mayor belleza del paisaje (Murgueitio & Ibrahim 2004). Sin embargo, uno de los principales limitantes para la propagación de estas plantas en paisajes ganaderos es la falta de dispersores primarios y la poca disponibilidad de sitios adecuados para la germinación de semillas y el establecimiento de plántulas.
Pese a que los dispersores primarios han sido desplazados de estos escenarios, se ha documentado que algunas especies generalistas de hormigas típicas de potreros, recogen activamente semillas de árboles con potencial en la rehabilitación ecológica de potreros (Escobar-Ramírez et al. 2011). En zonas neotropicales, se ha evidenciado que varias especies de hormigas pueden afectar en forma positiva el destino de semillas de árboles y arbustos que dependen principalmente de vertebrados frugívoros para su dispersión (Handel & Beattie 1990, Levey & Byrne 1993). Es posible entonces que estos insectos sociales altamente ubicuos y abundantes en hábitats fuertemente alterados como las pasturas, tengan una influencia importante sobre la diversidad vegetal.
Las hormigas transportan semillas a sitios que podrían favorecer la germinación y el establecimiento de plántulas (Hanzawa et al. 1988). Las semillas que resultan atractivas para las hormigas y son posteriormente transportadas, pueden ser abandonadas durante el camino y potencialmente germinar y establecerse en nuevos sitios (Horvitz 1981). Aquellas semillas que son llevadas al nido, "manipuladas" por las hormigas y luego descartadas en las galerías, pilas de basura o cerca de las entradas, pueden verse favorecidas por el suelo enriquecido en nutrientes que representan los hormigueros (Leal et al. 2007). Sin embargo, no es claro si las hormigas pueden afectar el proceso mismo de germinación a través de un efecto mecánico (escarificación con las mandíbulas) o químico (rociándolas con secreciones de sus glándulas exocrinas). A nivel ecológico, los factores que afectan la germinación de semillas son fundamentales para el proceso de sucesión vegetal, pues tanto la capacidad de dispersión y colonización como la posibilidad de germinación afectan directamente el éxito de las diásporas. La semilla, por tanto, es vital para el éxito de la planta.
Este estudio examinó cómo incide la escarificación en la germinación de semillas de tres especies de árboles propios del bosque seco tropical de Colombia, cuyas semillas son removidas por hormigas en potreros. Se partió del interrogante: ¿la escarificación es necesaria para la germinación de dichas semillas? Se identificó la especie con mayor porcentaje de germinación en laboratorio y se realizaron pruebas en invernadero y campo con el fin de evaluar cómo la tierra de nidos y la manipulación por parte de las hormigas afectan la germinación de esta especie, potencial en la rehabilitación ecológica de potreros. Se plantearon las siguientes preguntas: ¿la manipulación de semillas de Senna spectabilis (DC) H.S. Irwin & Barneby por Solenopsis geminata (Formicidae: Myrmicinae) favorece su germinación? y, ¿el suelo de nidos de S. geminata favorece la germinación de semillas de S. spectabilis?
MATERIALES Y MÉTODOS
Material vegetal y sitio de recolección
Se recolectaron semillas de tres especies de árboles propios de bosque seco tropical con potencial en la rehabilitación ecológica de potreros, cuyas semillas son transportadas por hormigas en potreros del valle geográfico del río Cauca (Escobar-Ramírez et al. 2011): Pithecellobium dulce (Roxb.) Bent., Guazuma ulmifolia Lam. y S. spectabilis (Tabla 1).
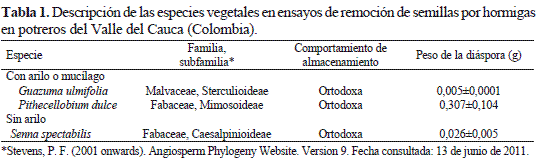
Las semillas de P. dulce y S. spectabilis se recolectaron en dos localidades: 1) Cali (3°27'26"Norte, 76°31'42"Oeste), ciudad que se encuentra a una elevación de 970 m y tiene una temperatura promedio anual de 24 °C (Espinal 1967) y 2) Ecoparque Río Pance (3°20'Norte, 76°34'Oeste), ubicado a 12 km de Cali, sobre las riberas del río Pance a una altura de 1100 msnm (Osorio 1991). Las dos localidades presentan características de una zona de vida de bosque seco tropical (bs-T) según la clasificación de Holdridge (Espinal 1967). Allí se adquirieron frutos maduros que estuvieran sobre las plantas, en aproximadamente seis árboles para cada especie, y las semillas se extrajeron manualmente mediante la impactación mecánica de los frutos. Posteriormente se almacenaron en bolsas plásticas selladas herméticamente a una temperatura de 5°C hasta su utilización. Por otra parte, las semillas de G. ulmifolia se obtuvieron de la casa comercial El Semillero®.
En general, las semillas empleadas en este estudio son ortodoxas, es decir, que aumentan su longevidad cuando se reduce el contenido de humedad de las semillas y se almacenan a bajas temperaturas (Ellis & Roberts 1981, Berjak & Pammenter 2002). En el caso de P. dulce, se establece que sus semillas se asemejan más a semillas ortodoxas que a semillas recalcitrantes (Royal Botanic Gardens Seed Information Databasse -SID- 2008). Además, se ha demostrado que las semillas de estas especies requieren procedimientos pre-germinativos que permitan acelerar el proceso de imbibición y asegurar la germinación (Baskin et al. 1998, Royal Botanic Gardens Seed Information Databasse -SID- 2008). Con tratamientos de escarificación como la abrasión mecánica de la testa y bajo condiciones de temperatura y fotoperiodo de 26 °C y 12/12 horas respectivamente, los porcentajes de germinación de estas especies pueden ser de 87 a 100% (Francis 1991, Royal Botanic Gardens Seed Information Databasse -SID- 2008).
Pruebas de viabilidad y efecto de la escarificación
En mayo de 2010 se realizaron pruebas de germinación con el objetivo de conocer el tiempo de germinación y verificar la viabilidad de las semillas recolectadas. Para ello se efectuó muestreo al azar del lote de semillas usando cuatro repeticiones en el caso de S. spectabilis y P. dulce, y dos para G. ulmifolia. Cada repetición consistió de 50 semillas dispuestas equidistantemente sobre hojas de papel absorbente como sustrato, en cajas de Petri debidamente etiquetadas. Las muestras se pusieron en una germinadora DiEs modelo KU115, con condiciones preestablecidas de temperatura de 25/30 °C durante períodos de 16/8 horas respectivamente, y de fotoperiodo de 12/12 horas. La lectura de germinación fue diaria y el riego con agua destilada se hizo hasta el punto de saturación una sola vez al momento de iniciar la prueba. De acuerdo con Fenner & Thompson (2005), el criterio de germinación fue emergencia de la radícula. La prueba de germinación tuvo una duración de diez días. Para establecer si las especies de estudio requieren escarificación para germinar, se sembraron simultáneamente en las mismas condiciones ambientales, semillas a las que se les practicó escarificación mecánica, que consistió en realizar una incisión de 1 mm o menos sobre la testa de la semilla con un cortaúñas común previamente esterilizado, teniendo cuidado de no afectar la zona embrionaria. En este caso se usó el mismo número de repeticiones modificando a 25 el número de semillas en cada una.
Manipulación de semillas en colonias artificiales de S. geminata
En julio de 2010, en una zona de potrero abierto de la hacienda Sachamate, ubicada en la ciudad de Jamundí (3°16'34"Norte, 76°33'24"Oeste), se seleccionaron aleatoriamente 10 nidos de S. geminata para realizar su extracción y la del suelo adyacente (separado de los nidos por una distancia entre uno y dos metros). Esta especie de hormiga fue seleccionada para el estudio debido a que presenta alta abundancia en hábitats perturbados (Ness & Bronstein 2004), especialmente en potreros del valle geográfico del río Cauca (Armbrecht & Ulloa-Chacón 1999, Achury et al. 2008). Solenopsis geminata transporta semillas hacia sus nidos (Holway et al. 2002) y puede dispersarse rápidamente en sistemas en los que la vegetación del suelo ha sido reducida (Perfecto 1991). A partir de los nidos, se recolectaron obreras y crías de S. geminata, con las cuales se establecieron colonias artificiales siguiendo el método sugerido por Herrera & Armbrecht (2007) y Ulloa-Chacón (2003): las hormigas y las crías obtenidas de cada nido se transfirieron a cajas plásticas de 24 cm de ancho y 10 cm de profundidad. Los bordes internos se impregnaron con Tanglefoot® ( Grand Rapids Michigan; sustancia pegajosa que no se seca) para evitar el escape de hormigas. Cada caja contenía un nido artificial en su interior, elaborado de pasta-yeso y protegido de la luz con un recipiente oscuro. Las cajas se cubrieron con una tapa plástica con dos agujeros sellados con malla metálica muy fina (o muselina) para permitir el intercambio de gases. Posteriormente, a cada nido se le ofreció una semilla de S. spectabilis, especie que presentó el mayor éxito germinativo en las pruebas de viabilidad en comparación con P. dulce y G. ulmifolia. A partir de allí se realizó seguimiento semanal y al finalizar el experimento, se hicieron pruebas de germinación a las semillas ofrecidas a las hormigas con el objetivo de observar si la capacidad de germinación había sido alterada por la manipulación de las hormigas.
Efectos de la transformación del suelo sobre las dinámicas de germinación
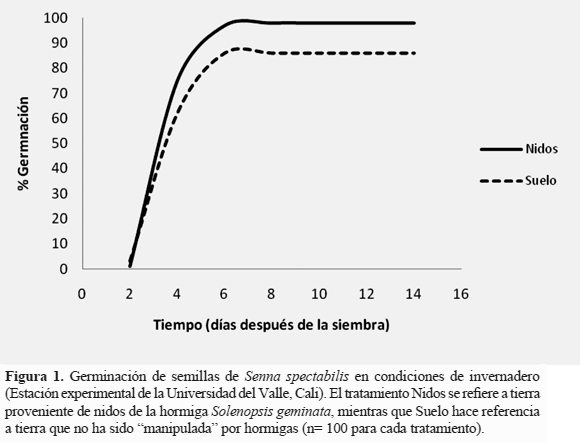
Pruebas de germinación en invernadero. Las muestras de suelo de nidos y de suelo adyacente extraídos de la hacienda Sachamante se almacenaron en recipientes con dos agujeros en la parte superior, cubiertos con malla metálica o tela para permitir el intercambio de gases mientras eran llevadas a la Estación Experimental de Biología de la Universidad del Valle. El suelo se transfirió a recipientes plásticos de 13,5x9,0x13,5 cm dividiéndolo en dos tratamientos: el primero fue suelo proveniente de nidos de S. geminata del que se extrajeron las hormigas y el segundo fue suelo adyacente a dichos nidos. Para cada tratamiento se emplearon diez recipientes, a los que se les realizaron agujeros en la parte inferior para permitir el drenaje de agua. En cada recipiente se sembraron diez semillas de S. spectabilis escarificadas, separadas una de otra por dos centímetros y enterradas a una profundidad de un centímetro en el substrato. Los recipientes se ubicaron en el invernadero en dos filas de diez espacialmente intercalando los dos tratamientos. A partir de la fecha de siembra se realizó monitoreo y riego cada dos días. El criterio de germinación fue emergencia de la plúmula. La prueba tuvo una duración de diez días. A lo largo de este tiempo se registró el número de semillas en las que se observó germinación.
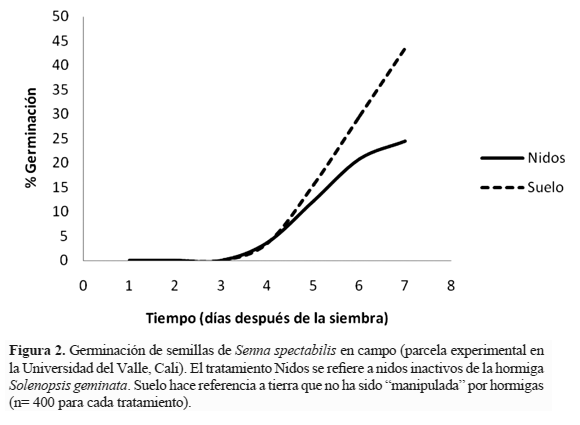
Pruebas de germinación en campo. En septiembre de 2010 se demarcaron sobre una parcela de aproximadamente 0,5 ha desprovista de árboles en el campus la Universidad del Valle (Cali) 16 bloques que consistían cada uno en dos tratamientos: nido inactivo de S. geminata y control, es decir, suelo que no había sido alterado por las hormigas. En cada tratamiento se sembraron 25 semillas de S. spectabilis previamente escarificadas, enterradas a una profundidad menor a un centímetro en el sustrato. Los tratamientos se protegieron con una malla de alambre sobre cada área sembrada con el objeto evitar la actividad de vertebrados. A partir de la fecha de siembra se realizó monitoreo y riego diariamente con dos litros de agua, excepto en días de lluvia cuando no fue necesario. El criterio de germinación fue emergencia de la plúmula. La prueba tuvo una duración de siete días. A lo largo de este tiempo se registró el número de semillas en las que se observó germinación.
Análisis de datos
En todos los experimentos la variable de respuesta fue porcentaje de semillas germinadas, la cual fue transformada mediante la función arcoseno √% de germinación. Se evaluaron los supuestos de normalidad y homogeneidad de varianzas por medio de las pruebas de Kolmogorov-Smirnov y Levene, respectivamente. Posteriormente se utilizó una prueba pareada de t-student para comparar las medias de la variable de respuesta en invernadero y campo (semillas germinadas dentro y fuera de los nidos).
RESULTADOS
Pruebas de viabilidad y efecto de la escarificación
Las semillas de S. spectabilis presentaron alta viabilidad y germinación rápida cuando se les realizó escarificación, pasando de un porcentaje de germinación de 0% en diez días a 98,5% en tres días. Se observó lo contrario en las semillas de P. dulce, que a pesar de presentar alta viabilidad, disminuyeron el porcentaje de germinación de 94 a 87% cuando se realizó escarificación. Las semillas de G. ulmifolia, por otra parte, tuvieron una respuesta positiva a la escarificación incrementando la germinación de 5 a 37%, sin embargo, el porcentaje máximo de germinación es indicador de baja viabilidad.
Manipulación de semillas en colonias artificiales de S. geminata
Las hormigas no efectuaron ningún cambio visible a las semillas de S. spectabilis ofrecidas. Tampoco hubo germinación en ninguna de estas semillas en la germinadora después de 30 días. Por lo tanto, no se encontró evidencia de algún tipo de escarificación física (por mandíbulas u otras estructuras que pudieran afectar la testa de la semilla), ni química (a través de glándulas o aguijón).
Efectos de la transformación del suelo sobre las dinámicas de germinación
Pruebas de germinación en invernadero. La velocidad de germinación en S. spectabilis fue similar en nidos y suelo adyacente. El mayor número de plántulas, independiente del tratamiento al que pertenecían, emergieron el cuarto día después de la siembra (Figura 1). Sin embargo, el porcentaje de germinación sí fue diferente. En el tratamiento "nidos" germinaron en total 98 semillas, un número significativamente mayor al de tratamiento "suelo" (t = 2,42, p= 0,02), que presentó 86 semillas germinadas.
Pruebas de germinación en campo. Los resultados obtenidos en esta etapa contrastaron con aquellos en invernadero. La velocidad de germinación fue mayor en el tratamiento "suelo", superando al tratamiento "nidos" a partir del quinto día después de la siembra (Figura 2). También fue significativamente mayor el porcentaje de germinación total de las semillas sembradas en suelo adyacente (t = 3,4, p < 0,001), con 174 versus 98 semillas germinadas en los nidos de S. geminata.
DISCUSIÓN
Los resultados de este estudio confirman que la escarificación favorece la germinación de las semillas de S. spectabilis y G. ulmifolia. En cuanto a S. spectabilis, este trabajo constituye uno de los primeros intentos en el conocimiento de las características germinativas de la especie en el país. Debido a su comportamiento en los tratamientos semillas escarificadas vs. no escarificadas, se establece que las semillas presentan latencia física. En estos casos, el nivel de hidratación de la semilla juega un papel en el rompimiento de la latencia (Baskin & Baskin 2001). Así, la ausencia de germinación cuando no se realiza escarificación es consecuencia de la impermeabilidad de la semilla al agua.
La respuesta positiva de G. ulmifolia a la escarificación, coincide con lo hallado por Hernández et al. (2001) y Muñoz et al. (2004). Esta especie, como S. spectabilis, presenta una fuerte impermeabilidad de la cubierta que se elimina en gran medida con tratamientos pregerminativos. Sin embargo, aún cuando la escarificación es realizada con ácido, imitando lo que ocurriría cuando las frutas y las semillas son masticadas y pasadas a través del sistema digestivo de un animal (Francis 1991), la germinación puede presentar limitaciones que dependen de las características intrínsecas de la semilla. Por otro lado, aunque la mayoría de géneros de la familia Fabaceae tiene semillas que presentan latencia física, el género Pithecellobium posee semillas permeables (Baskin & Baskin 2001). Debido a esto, las semillas de P. dulce estuvieron afectadas negativamente por la escarificación, estas semillas presentan una alta capacidad germinativa y a diferencia de S. spectabilis y G. ulmifolia, no requieren tratamientos especiales para su emergencia.
Los resultados de los experimentos de germinación en invernadero fueron consistentes con los de Leal et al. (2007) al haber incremento en la germinación de las semillas cuando se presenta en nidos de hormigas comparado con sitios adyacentes. Esto puede deberse a que la construcción de los nidos de hormigas modifica positivamente algunos aspectos de las propiedades del suelo (Green et al. 1999, Frouz & Jilková 2008), mejorando así la calidad de los suelos impactados. Dichos cambios podrían tener una influencia significativa en la infiltración de agua desde la superficie del suelo. Los bioporos construidos por las hormigas son considerados particularmente importantes en situaciones donde el suelo desarrolla capas duras, como los cultivos degradados, sitios dedicados al pastoreo y áreas mineras en rehabilitación. La estabilidad de los bioporos probablemente incrementa su papel en la infiltración y distribución de agua y gases en el perfil, aún después de que el nido ha sido abandonado (Nkem et al. 2000). De esta manera, una adecuada entrada de agua hacia las semillas previamente escarificadas habría proporcionado las condiciones ideales para la germinación de S. spectabilis.
Sin embargo, en campo la germinación presentó un comportamiento opuesto al observado en invernadero. Este resultado permitió descubrir que los factores ambientales interactúan entre sí para determinar el éxito de la germinación. Los nidos de S. geminata en condiciones estrictamente controladas fueron más favorables para la germinación por las razones arriba expuestas, pero precisamente estos factores se convirtieron en "trampas" en condiciones de alta precipitación y radiación solar. Las lluvias fuertes incidieron directamente sobre las semillas que estaban en suelo removido (nidos de hormigas), porque la energía cinética de las gotas durante los aguaceros entierra más profundamente algunas semillas, mientras que la posterior radiación solar las atrapa al secarse la tierra. La interacción semilla-ambiente es un elemento muy relevante, y como lo observaron Levey & Byrne (1993), si los nidos no habían sido completamente desalojados, las hormigas pudieron haber dañado algunas semillas antes del inicio de la germinación. Es de anotar que al momento de estos experimentos se produjo el efecto de La Niña (IDEAM 2010), por lo cual lo observado coincide con condiciones más o menos extremas de precipitación fuerte.
Aunque los experimentos con colonias artificiales de S. geminata sugieren que éstas no efectúan ningún cambio positivo en la testa de las semillas de S. spectabilis, tampoco se observaron cambios negativos, por lo que se recomienda realizar estudios que involucren un mayor número de semillas. Es probable que las semillas de estas leguminosas, de gran valor para los ganaderos, deban pasar por el tracto digestivo del ganado (que apetece mucho sus frutos) o estar expuestas a lluvias y fluctuación en la radiación solar, aspectos que deterioran la testa haciendo que las semillas rompan el estado de latencia (Doucette et al. 2001, Miceli-Méndez et al. 2008), luego de lo cual queden aptas para germinar. En este caso, las hormigas actuarían como dispersores secundarios y no como primarios.
En conclusión, este estudio demuestra la necesidad de realizar tratamientos pregerminativos como escarificación mecánica a semillas de S. spectabilis, especie que tiene un uso potencial en la rehabilitación ecológica de sistemas degradados. No se encontraron resultados que apoyen la suposición que los nidos de hormigas en campo favorecen de alguna manera la germinación de semillas de S. spectabilis. Sin embargo, el suelo de nidos podría ser útil en la elaboración de viveros de esta especie que se piensen usar en rehabilitación ecológica. Debido a que el proceso de germinación es diferente del proceso de establecimiento, para evaluar el papel potencial de las hormigas en la rehabilitación ecológica de sistemas degradados, se recomienda evaluar el aporte de los nidos en el establecimiento, el desarrollo y la supervivencia de las plántulas emergidas de las semillas que logran germinar.
AGRADECIMIENTOS
A Zoraida Calle, James Montoya por su asesoría y valiosas sugerencias en campo. A Aymer Andrés Vásquez y Dagoberto Sinisterra por la colaboración en campo. Al Grupo de Investigación en Ecología de Agroecosistemas y Hábitats Naturales (GEAHNA) de la Universidad del Valle. También a los evaluadores anónimos que realizaron importantes sugerencias a la versión final del manuscrito. Este estudio fue financiado por Colciencias mediante el proyecto "Las Hormigas como agentes funcionales para la recuperación ecológica de sistemas ganaderos" (Cód. 110645221048) programa de Ciencias Básicas y por la Universidad del Valle ( Cali, Colombia ).
LIERATURA CITADA
1. ACHURY MORALES, R.A., CHACON DE ULLOA, P. & A.M. ARCILA. 2008. Interacciones competitivas entre Wasmannia auropunctata y otras especies de hormigas en bosque seco tropical. Revista Colombiana de Entomología 34(2): 209-216. [ Links ]
2. ARMBRECHT, I. & P. ULLOA-CHACÓN. 1999. Rareza y diversidad de hormigas en fragmentos de Bosque Seco Colombianos y sus matrices. Biotropica 31 (4): 646-653. [ Links ]
3. BASKIN, C.C. & J.M. BASKIN. 2001. Seeds: Ecology, biogeography and evolution of Dormancy and Germination. Academic Press. Londres. 666p. [ Links ]
4. BASKIN, J.M., NAN, X. & C.C. BASKIN. 1998. A comparative study of seed dormancy and germination in an annual and a perennial species of Senna (Fabaceae). Seed Science Research 8: 501-512. [ Links ]
5. BERJAK, P.N. & N.W. PAMMENTER. 2002 Orthodox and Recalcitrant Seeds. Cap. 4, págs. 137-147. En: J.A. Vozzo (ed.). Tropical Tree Seed Manual. USDA Forest Service, Washington D.C. [ Links ]
6. DOUCETTE, K.M., WITTENBERG, K.M. & W.P. MCCAUGHEY. 2001. Seed recovery and germination of reseeded Species fed to Cattle. Journal of Range Management 54(5): 575-581. [ Links ]
7. ELLIS, R.H. & H. ROBERTS. 1981. The quantification of ageing and survival in orthodox seeds. Seed Science and Technology 9: 373-409. [ Links ]
8. ESCOBAR-RAMÍREZ, S., DUQUE, S., HENAO, N., HURTADO-GIRALDO, A. & I. ARMBRECHT. 2011. Removal of Nonmyrmecochorous Seeds by Ants: Role of Ants in Cattle Grasslands. Psyche, 2012: 1-8. [ Links ]
9. ESPINAL, L.S. 1967. Apuntes sobre Ecología Colombiana. Universidad del Valle, Cali. 29p. [ Links ]
10. FENNER, M. & K. THOMPSON. 2005. The Ecology of Seeds. Cambridge University Press. Cambridge. 250pp. [ Links ]
11. FOLGARAIT, P. 1998. Ant biodiversity and its relationship to ecosystem functioning: a review. Biodiversity and Conservation 7: 1221-1244. [ Links ]
12. FRANCIS, J.K. 1991. Guazuma ulmifolia Lam. Guácima. SO-ITF-SM-47. LA: U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station. New Orleans. 5p. [ Links ]
13. FROUZ, J. & V. JILKOVÁ. 2008. The effect of ants in soil properties and processes (Hymenoptera: Formicidae). Myrmecological News 11: 191-199. [ Links ]
14. GREEN, W.P., PETTRY, D.E. & R.E. SWITZER. 1999. Structure and hydrology of mounds of the imported fire ants in the southeastern United States. Geoderma 93: 1-17. [ Links ]
15. HANDEL, S.N. & A.J. BEATTIE. 1990. Semillas dispersadas por hormigas. Investigación y Ciencia 263: 76-83. [ Links ]
16. HANZAWA, F.M., BEATTIE, A.J. & D.C. CULVER. 1988. Directed dispersal: Demographic analysis of an ant-seed mutualism. The American Naturalist 13(1): 1-13. [ Links ]
17. HERNÁNDEZ, G., SÁNCHEZ, L.R. & F. ARAGÓN. 2001. Tratamientos pregerminativos en cuatro especies arbóreas de uso forrajero de la selva baja caducifolia de la Sierra de Manatlán. Foresta Veracruzana 3(1): 9-15. [ Links ]
18. HERRERA, J. & I. ARMBRECHT. 2007. Ciclo de vida y potencial reproductivo de la hormiga depredadora Solenopsis cf. picea (Hymenoptera: Formicidae). Revista Colombiana de Entomología 33(1): 64-69. [ Links ]
19. HOLWAY, D.A., LACH, L. SUÁREZ, A.V., TSUTSUI, N.D. & T.J. CASE. 2002. The causes and consequences of ant invasions. Annual Review of Ecology and Systematics 33:181–233. [ Links ]
20. HORVITZ, C.C. 1981. Analysis of How Ant Behaviors Affect Germination in a Tropical Myrmecochore Calathea microcephala (P. & E.) Koernicke (Marantaceae): Microsite Selection and Aril Removal by Neotropical Ants, Odontomachus, Pachycondyla, and Solenopsis (Formicidae). Oecologia 51(1): 47-52. [ Links ]
21. INSTITUTO DE HIDROLOGÍA, METEOROLOGÍA Y ESTUDIOS AMBIENTALES DE COLOMBIA-IDEAM. 2010. Análisis de las condiciones meteorológicas presentadas durante el mes de septiembre de 2010. Boletín Climatológico Mensual. Bogotá, D.C. [ Links ]
22. LEAL, I.R., WIRTH, R. & M. TABARELLI. 2007. Seed dispersal by ants in the semi-arid catinga of North-east Brazil. Annals of Botany 99: 885-894 [ Links ]
23. LEVEY, D.J. & M.M. BYRNE. 1993. Complex ant-plant interactions: Rain-forest ants as secondary dispersers and post-dispersal seed predators. Ecology 74: 1802-1812. [ Links ]
24. MICELI-MÉNDEZ, C.L., FERGUSON, B.G. & N. RAMÍREZ-MARCIAL. 2008. Seed Dispersal by Cattle: Natural History and Applications to Neotropical Forest Restoration and Agroforestry. Cap. 7, págs. 165-191. En: R.W. Myster (ed.). Post-Agricultural Succession in the Neotropics. Springer. Nueva York. [ Links ]
25. MUÑOZ, B.C, SÁNCHEZ, J.A. & W. ALMAGUER. 2004. Germinación, dormancia y longevidad potencial de las semillas de Guazuma ulmifolia. Pastos y Forrajes 27(1): 25-33. [ Links ]
26. MURGUEITIO, E. & M. IBRAHIM. 2004. Ganadería y medio ambiente en América Latina en: XII Congreso Venezolano de Producción e Industria Animal. Noviembre de 2004. [ Links ]
27. MURGUEITIO, E., CALLE, Z, URIBE, F., CALLE, A. & S. BALDOMERO. 2011. Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest Ecology and Management 261: 1654-1663. [ Links ]
28. NESS, J.H. & J.L. BRONSTEIN. 2004. The effects of invasive ants on prospective ant mutualists. Biological Invasions 6: 445-461. [ Links ]
29. NKEM, J.N., LOBRY DE BRUYN, L.A., GRANT, C.D. & N. R. HULUGALLE. 2000. The impact of ant bioturbation and foraging activities on surrounding soil properties. Pedobiologia 44: 609-621. [ Links ]
30. OSORIO, O. 1991. Proyecto arqueológico sobre la cuenca del río Pance. Informe de divulgación No. 1230. INCIVA. Calima, Colombia. 2pp. [ Links ]
31. PERFECTO, I. 1991. Dynamics of Solenopsis geminata in a tropical fallow field after ploughing. Oikos 62(2): 139-144. [ Links ]
32. ROYAL BOTANIC GARDENS KEW. 2008. Seed Information Database (SID). Version 7.1. URL: http://data.kew.org/sid/ (Mayo de 2008) [ Links ]
33. ULLOA-CHACÓN, P. 2003. Biología reproductiva de Wasmannia auropunctata (R.) (Hymenoptera: Formicidae). Revista de la Academia Colombiana de Ciencias Exactas Físicas y Naturales 27(104): 441-447. [ Links ]

