










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.34 no.1 Bogotá Jan./June 2012
VICTOR H. GONZALEZ
TERRY L. GRISWOLD
Division of Entomology, Natural History Museum, 1501 Crestline Drive – Suite 140, University of Kansas, Lawrence, Kansas 66045, USA. victorgonzab@gmail.com
USDA-ARS, Bee Biology & Systematics Laboratory, Utah State University, Logan, Utah 84322-5310, USA. terry.griswold@ars.usda.gov
ABSTRACT
Three new species of cleptobiotic stingless bees of the genus Lestrimelitta Friese, L. opita sp. n. and L. huilensis sp. n. from Colombia and L. catira sp. n. from Venezuela, are described and figured. The males of the Central American species L. chamelensis, L. danuncia, and L. mourei are also described and figured. New geographical records and an updated key to species from Central America and northern South America are provided.
Key words. Anthophila, Apoidea, Meliponini, Colombia, Venezuela, taxonomy.
RESUMEN
Se describen e ilustran tres especies nuevas de abejas sin aguijón cleptobióticas del género Lestrimelitta Friese, L. opita sp. n. y L. huilensis sp. n. de Colombia y L. catira sp. n. de Venezuela. También se describen e ilustran los machos de las especies Centroamericanas L. chamelensis, L. danuncia y L. mourei. Se presentan registros geográficos nuevos y una clave actualizada para las especies de Centroamérica y norte de Suramérica.
Palabras clave. Anthophila, Apoidea, Meliponini, Colombia, Venezuela, taxonomía.
Recibido: 18/11/2011
Aceptado: 15/12/2011
INTRODUCTION
Unusual and astonishing behaviors are often found in the commonly encountered stingless bees, highly eusocial bees restricted to the tropical areas of the world but more abundant and diverse in the Western Hemisphere (e.g., Michener 2007). Examples of these are the defensive behavior of Oxytrigona Cockerell, whose workers repel intruders by biting and depositing into the skin a mandibular secretion that contains formic acid and causes a long-lasting blister (e.g., Michener 2007); the mutualistic relationship of Plebeia coccidophila (Camargo & Pedro) with a soft-scale insect (Cryptostigma sp.; Coccidae), which is housed inside the bee nest and provides sweet secretions and additional wax to build the nest (Camargo & Pedro 2002); the obligate necrophagy of Trigona hypogea Silvestri, T. crassipes (Fabricius), and T. necrophaga Camargo & Roubik, the only known species to use the flesh of dead animals as their only source of protein (Roubik 1982, Camargo & Roubik 1991); the lacrophilic behavior (tear drinking habits) of minute Asian stingless bees of the genera Lisotrigona Moure and Pariotrigona Moure at mammalian eyes presumably to obtain proteins in addition to, or in lieu of pollen (Bänziger et al. 2009); the pollen or propolis collecting behavior exhibited by males of some Asian species which, while lacking a corbicula, frequently carry these materials on a metatibia that resembles those of the workers (Boongird & Michener 2010); and the obligate robbing or cleptobiotic behavior of the Neotropical Lestrimelitta Friese and the African Cleptotrigona Moure, which despite having nests of their own, steal food and nest materials from other stingless bees and even honey bees (Portugal-Araújo 1958, Sakagami et al., 1993). Trichotrigona Camargo & Moure, a monotypic genus from the Brazilian Amazon, is likely another cleptobiotic bee or possibly the only known meliponine social parasite, given the reduction of pollen-manipulating structures on the hind leg (Camargo & Pedro 2007a, Michener 2007).
Lestrimelitta is widely distributed in the Neotropical region, with apparent center of diversity in Brazil; 14 of the 21 currently recognized species are from there (Table 1). Such a high diversity of Lestrimelitta in Brazil is likely due to the country's large expanse that covers a significant area of South America as well as the result of the effort of numerous Brazilian researchers to document their bee fauna. Other South American countries, such as Colombia, despite having preliminary checklists, are still in early stages of exploration as indicated by the number of taxa discovered in recent years (e.g., Engel 1997, 2009, 2010, Engel & Gonzalez 2009, Gonzalez 2004, 2006, Gonzalez & Engel 2004, 2011, Gonzalez & Ruz 2007, Smith-Pardo & Gonzalez 2007, 2009, Gonzalez et al., 2010, Gonzalez & Florez 2011, Gonzalez & Griswold 2011, Gonzalez & Engel 2012).
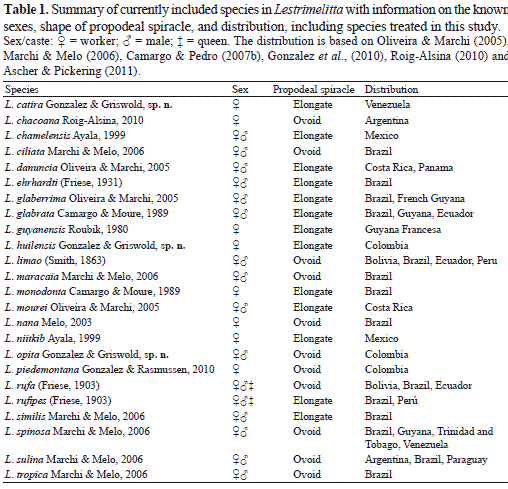
Lestrimelitta is likely to be related to Plebeia Schwarz, as suggested by both morphological and molecular studies (Michener 2007, Rasmussen & Cameron (2010). When compared to other stingless bee genera, Lestrimelitta species seem to be monotonously similar at first glance, but recent works (e.g., Ayala 1999, Oliveira 2002, Melo 2003, Oliveira & Marchi 2005, Marchi & Melo 2006, Gonzalez et al., 2010, Roig-Alsina 2010) have highlighted and illustrated a suitable set of characters that are reliable in species recognition. Those characters include the shape of the propodeal spiracle, length of the mesotibial spur, presence or absence of hairs on the body, and the length, density, and type of pubescence. Two large species groups can be recognized within Lestrimelitta based on the shape of the propodeal spiracle (Table 1): the exclusively South American limao species group, which consists of species with an ovoid propodeal spiracle, 2–3 times longer than broad (c.f. Marchi & Melo 2006; fig. 1, Gonzalez et al. 2010; fig. 2D), and the ehrhardti species group found in both Central and South America distinguished by an elongate propodeal spiracle, at least 4.6 times longer than broad (c.f. Marchi & Melo 2006; fig. 2).

Herein we describe three new species of Lestrimelitta: one of the limao species group from Colombia and two of the ehrhardti species group, one from Colombia and one from Venezuela. We also describe the previously unknown males of three Central American species of the ehrhardti species group: L. chamelensis, L. danuncia, and L. mourei. We also provide new geographical records and an updated key to species from Central America and northern South America.
MATERIAL AND METHODS
Morphological terminology and measurements (e.g., interalveolar, alveolocular, ocellocular distances, etc.) follow that of Michener (2007). Measurements were taken using an ocular micrometer on a Leica® MZ12 stereomicroscope. Photomicrographs were taken using a Keyence® VHX- 500F Digital Imaging System. The abbreviations S and T are used for metasomal sterna and terga. The symbol for female and the word itself are used below for worker, not queen. Institutional acronyms used herein are: BBSL, U.S. National Pollinating Insects Collection, Bee Biology and Systematics Laboratory, Utah State University, Logan, UT, USA; ICN, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia; and USNM, National Museum of Natural History, Washington, DC, USA. The novelty of the species described herein were confirmed by comparison with specimens (including types) of other Lestrimelitta species deposited at BBSL, ICN, and the Snow Entomological Collection, Division of Entomology, Natural History Museum, University of Kansas, Lawrence, Kansas, USA.
SYSTEMATICS Tribe Meliponini Lepeletier de Saint Fargeau, 1836 Genus Lestrimelitta Friese, 1903
Diagnosis. Lestrimelitta consists of small (4–7 mm long) species with shiny, sparsely haired bodies that lack the metatibial corbiculae for carrying pollen found in other stingless bees. The genus can also be recognized by the following combination characters: metatibia without penicillum and with a rastellum consisting of tapering hairs; first flagellomere shorter than combined lengths of second and third flagellomeres in workers, shorter than second flagellomere in males; male sixth and seventh sterna with long basolateral apodemes; male sixth sternum medially emarginated on distal margin.
Lestrimelitta catira, new species
Holotype. Venezuela: ♀, Caripito, Edo. Monagas – XII-14-43, R. Maduro / Robbing honey from hive / C.H. Ballou, No BHT / LotNo 44-21773 (USNM).
Paratypes. 2♀, same data as holotype (BBSL, USNM).
Diagnosis. This species is most similar to L. rufipes (Friese) and L. huilensis, which share the following characters: propodeal spiracle elongate; vertex, preoccipital border and anterior margin of mesoscutum with erect hairs; and pronotal lobe and tegula without erect hairs. It can be easily distinguished by body color predominantly reddish-brown, distinctly lighter than in those species, and body pubescence on vertex, preoccipital border, and anterior margin of mesoscutum much shorter and sparser, and on disc of mesoscutellum practically absent (Figs. 1–3). In L. rufipes, the erect hairs of these areas are in general long and abundant; on vertex and preoccipital border at least as long as median ocellar diameter; on mesoscutum present along anterior margin as well as on anterior one-third of disc medially; on disc of mesoscutellum abundant, about as long as (about twice as long as median ocellar diameter) or slightly shorter than those on the distal margin. Refer to diagnosis of L. huilensis for comparative comments with that species.
Description. ♀. Total body length 6.7 mm; forewing length 4.4 mm; head width 2.3 mm. Head 1.3 times broader than long; inner orbits of compound eyes subparallel; malar area 1.5 times median ocellar diameter; clypeus 3.5 times broader than long; interalveolar distance shorter (0.8x) than alveolar diameter; alveolocular distance about twice as long as alveolar diameter, 2.5 times interalveolar distance; interocellar distance 2.8 times median ocellar diameter, shorter (0.8x) than ocellocular distance; scape about 6.3 times longer than wide; pedicel longer than broad; first flagellomere about as long as broad, longer than second flagellomere, remaining flagellomeres broader than long, except apical flagellomere much longer than broad; compound eye 2.5 longer than broad; gena broader than width of compound eye in profile. Propodeal spiracle elongate, about 5.2 times longer than wide; metatibia about three times longer than broad.
Integument smooth and shiny between minute, sparse punctures as in other species of the genus; terga and sterna weakly lineolate-imbricate.
Color predominantly reddish-brown, except yellowish to light reddish-brown on clypeus distally, labrum, outer surface of mandible, anterior surface of scape, legs, propodeal triangle, and sterna. Wing membrane yellowish; veins and pterostigma light brown.
Body largely glabrous, sparsely covered with minute, appressed, simple yellowish to whitish hairs except long, stout, erect hairs as follows: distal margin of labrum and inferior margin of mandible (longest at least twice median ocellar diameter); vertex and preoccipital border with few sparse hairs (at most width of median ocellar diameter); anterior and lateral margins (next to axilla) of mesoscutum, axilla, and mesoscutellum (1–2 times median ocellar diameter), hairs longer on mesoscutellum; mesepisternum ventrally, meso- and, metacoxae, mesotibia, and propodeum laterally (1–1.5 times median ocellar diameter); posterior margin of metatibia, sides of T2–T4, entire T5 and T6 with dark brown hairs (1.5–2 times median ocellar diameter) (Fig. 4).
Etymology. The specific epithet is a word used by Venezuelan people to describe a blonde or red-headed person; this word is used herein in reference to the reddish coloration of this species.
Comments. The mesotibial spurs of the holotype and one of the paratypes are presumably broken-off because in the remaining paratype, which has the head glued to a point and missing left leg, the right mesotibial spur is long, about 1.3 times longer than mesobasitarsal width.
Lestrimelitta huilensis, new species

Holotype. Colombia: ♀, Huila, Rivera, Cementerio, 29.mar.1994, 745 m, G. Nates. Leg. / ICN 037926 (Barcode label) (ICN).
Diagnosis. This species is most similar to L. rufipes and L. catira, which share the following characters: propodeal spiracle elongate; vertex, preoccipital border, and anterior margin of mesoscutum with erect hairs; and sides of mesepisternum, pronotal lobe, and tegula without erect hairs. It differs from L. rufipes in the vertex, preoccipital border, and anterior margin of mesoscutum with shorter, sparser erect hairs (Fig. 6); anterior one-third of disc of mesoscutum medially without erect hairs; mesoscutellum with disc covered by short, sparse hairs contrasting with long, abundant hairs on distal margin. It differs from L. catira in the body color predominantly dark reddish-brown (Figs. 5–9), pubescence dense on anterior margin of mesoscutum and mesoscutellum, on metasomal terga much shorter, sparser. In L. catira, the body color is predominantly reddish-brown, distinctly lighter than in L. huilensis; anterior margin of mesoscutum with sparser erect hairs, especially medially; disc of mesoscutellum largely glabrous; metasomal terga with longer, denser hairs (compare Figs. 4 and 7), particularly on sides of T2 and discs of terminal segments.
Description. ♀. Total body length 6.1 mm; forewing length 4.9 mm; head width 2.1 mm. Head 1.3 times broader than long; inner orbits of compound eyes subparallel; malar area 1.4 times median ocellar diameter; clypeus 3.1 times broader than long; interalveolar distance shorter (0.9x) than alveolar diameter; alveolocular distance 1.9 times alveolar diameter, 2.2 times interalveolar distance; interocellar distance 2.3 times median ocellar diameter, shorter (0.8x) than ocellocular distance; scape about 6.4 times longer than wide; pedicel longer than broad; first flagellomere about as long as broad, longer than second flagellomere, remaining flagellomeres broader than long, except apical flagellomere much longer than broad; compound eye 2.5 longer than broad; gena broader than width of compound eye in profile. Propodeal spiracle elongate, about 4.6 times longer than wide; mesotibial spur long, about 1.3 times mesobasitarsal width basally; metatibia about three times longer than broad.
Integument smooth and shiny between minute, sparse punctures as in other species of the genus; terga and sterna weakly lineolate-imbricate.
Color predominantly dark reddish-brown, lighter on labrum, mandible (except basally), clypeal margin, legs, T1 and T2, and sterna. Wing membrane yellowish; veins and pterostigma light brown.
Body largely glabrous, sparsely covered with minute, appressed, simple yellowish hairs except long, stout, erect hairs as follows: distal margin of labrum and inferior margin of mandible (longest at least twice median ocellar diameter); vertex and preoccipital border with few sparse hairs (at most half width of median ocellar diameter); anterior and lateral margins (next to axilla) of mesoscutum (1–2 times median ocellar diameter) (Figs. 6, 9); axilla (about as long as median ocellar diameter); mesoscutellum with sparse hairs on disc (about as long as median ocellar diameter), much longer (2 times median ocellar diameter) on distal margin (Fig. 8); mesepisternum ventrally, meso- and, metacoxae, mesotibia, and propodeum laterally (1–1.5 times median ocellar diameter); posterior margin of metatibia (1.3–2 times median ocellar diameter), sides of T2–T4, entire T5 and T6 with dark brown hairs (0.5–2 times median ocellar diameter) (Fig. 7).
Etymology. The specific epithet refers to Huila, the department in Colombia where the species is currently known to occur.
Comments. Based on the limited material available it appears that L. huilensis and L. catira are allopatric species inhabiting different ecosystems: L. catira occupying deltaic savannas in northeastern Venezuela, L. huilensis inhabiting dry forests along the Magdalena Valley in Colombia. Lestrimelitta huilensis is likely to be sympatric with L. opita, the only other Lestrimelitta species known to occur along the valley of the Magdalena River in Colombia. It can be easily separated from that species by the shape of the propodeal spiracle (ovoid in L. opita) and the presence of erect hairs on the vertex, preoccipital ridge, and anterior margin of mesoscutum (hairs absent in L. opita).
Lestrimelitta opita, new species

Holotype. Colombia: ♀, Tol. [Tolima], Armero, malaise trap, 26–30-I-1977, Peyton & Suarez (USNM).
Paratypes. 2♂, same date as holotype except I-5-II-197, E.L. Peyton (BBSL, USNM).
Diagnosis. Both sexes of this species are most similar to L. limao (Smith) and L. ciliata Marchi & Melo, which share the following characters: propodeal spiracle ovoid; sides of mesepisternum and disc of mesoscutum practically glabrous, without erect hairs; and mesotibial spur reduced. It can be easily distinguished from those species by anterior margin of mesoscutum without erect hairs (Figs. 12, 14).

Description. ♀. Total body length 5.3 mm; forewing length 4.3 mm; head width 2.1 mm.
Head 1.3 times broader than long; inner orbits of compound eyes subparallel; malar area about as long as median ocellar diameter; clypeus about 3.3 times broader than long; interalveolar distance shorter (0.8x) than alveolar diameter; alveolocular distance 1.8 times alveolar diameter, 2.1 times interalveolar distance; interocellar distance 2.4 times median ocellar diameter, shorter (0.9x) than ocellocular distance; scape about six times longer than wide; pedicel longer than broad; first flagellomere about as long as broad, longer than second flagellomere, remaining flagellomeres broader than long, except apical flagellomere much longer than broad; compound eye 2.4 longer than broad; gena broader than width of compound eye in profile. Propodeal spiracle ovoid, about three times longer than wide; metatibia about three times longer than broad.
Integument smooth and shiny between minute, sparse punctures as in other species of the genus; terga and sterna weakly lineolate-imbricate.
Color predominantly dark reddish-brown, lighter on labrum, mandible (except basally), legs (except metafemur and metatibia), and basal terga and sterna. Wing membrane yellowish; veins and pterostigma light brown.
Body largely glabrous, sparsely covered with minute, appressed, simple yellowish hairs except long, stout, erect hairs as follows: distal margin of labrum and inferior margin of mandible (longest at least twice median ocellar diameter); vertex with very few sparse hairs (at most half width of median ocellar diameter); pronotal lobe inferiorly, anterolateral corner and lateral margin (next to axilla) of mesoscutum, axilla, and disc of mesoscutellum with few dark brown hairs (about as long as width of median ocellar diameter or less); posterior margin of mesoscutellum with longer hairs (1.3–1.6 times median ocellar diameter) (Fig. 13); mesepisternum ventrally, meso- and, metacoxae, mesotibia, and propodeum laterally (1–1.5 times median ocellar diameter); posterior margin of metatibia, sides of T1–T4, entire T5 and T6 with dark brown hairs increasing in length towards apical terga (1.5–2 times median ocellar diameter) (Fig. 11).
♂. As in female except for yellowish inferior paraocular area (next to inner orbit of compound eye), distal margin of clypeus, labrum, mandible, and legs, and the following: Total body length 6.1–6.8 mm; forewing length 3.9–4.1 mm; head width 1.9–2.0 mm. Inner orbits of compound eyes slightly converging below; malar area short, about one-third width of median ocellar diameter; clypeus about 2.5 times broader than long; alveolocular distance 1.3 times alveolar diameter, 1.5 times interalveolar distance; interocellar distance about twice as long as median ocellar diameter, 1.2 times ocellocular distance; scape about 5.5 times longer than wide; pedicel about as long as broad; first flagellomere slightly broader than long, about as long as second flagellomere, remaining flagellomeres about as long as broad, except apical flagellomere much longer than broad; compound eye 2.3 longer than broad; gena narrower (~0.6x) than width of compound eye in profile. Metatibia 2.7 times longer than broad. T5 apical margin invaginated medially. S5–S7 and genital capsule as in figures 17–21. Pubescence in general longer and denser than in female, especially on mesofemur and mesotibia (0.5–1 times median ocellar diameter), metafemur and metatibia (1–2 times median ocellar diameter), and metasoma (Figs. 14–16).
Etymology. The specific epithet is a self-referential nickname for inhabitants of the department of Tolima, the area where this species is currently known.
Lestrimelitta chamelensis Ayala
Lestrimelitta chamelensis Ayala, 1999: 25 [♀].

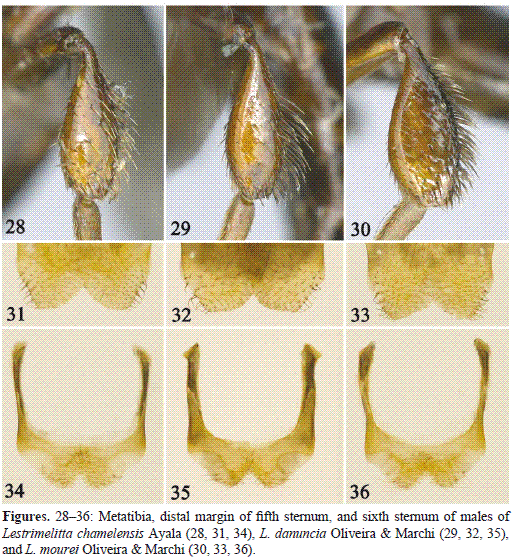
Diagnosis. The female of this species can be separated from other Central American species of Lestrimelitta by the following combination of characters: small body size (length about 5 mm); head and mesosoma practically glabrous, except for scattered hairs on preoccipital border and distal margin of mesoscutellum; mesotibia without erect hairs on outer surface; and mesotibial spur very reduced. The male can be recognized by mesotibial spur reduced, mesoscutellum with few short hairs on distal margin (at most as long as median ocellar diameter), and metatibia with sparse, rather long hairs (about as long as median ocellar diameter or longer) on outer surface (Fig. 28).
Description. ♂. Total body length 7.1 mm; forewing length 3.9 mm; head width 1.9 mm. Head 1.2 times broader than long; inner orbits of compound eyes slightly converging below (Fig. 23); malar area short, about half width of median ocellar diameter; clypeus about twice as broad as long; interalveolar distance shorter (0.8x) than alveolar diameter; alveolocular distance 1.4 times alveolar diameter, 1.8 times interalveolar distance; interocellar distance 2.1 times median ocellar diameter, 1.2 times ocellocular distance; scape about five times longer than wide; pedicel about as long as broad; first flagellomere slightly broader than long, about as long as second flagellomere, remaining flagellomeres about as long as broad, except apical flagellomere much longer than broad; compound eye about twice as long as broad; gena about half width of compound eye in profile. Propodeal spiracle elongate, 6.3 times longer than wide; mesotibial spur reduced, about one-third of mesobasitarsus width basally; metatibia 2.8 times longer than broad. T5 apical margin not invaginated medially; S5 and S6 as in figures 31 and 34, genital capsule and S7 as in L. opita.
Integument smooth and shiny between minute, sparse punctures as in other species of the genus; terga and sterna weakly lineolate-imbricate.
Color predominantly dark reddish-brown, darker on face and mesoscutum, yellowish to light reddish-brown on clypeus distally, labrum, anterior surface of scape, mandible, legs, and sterna. Wing membrane brownish; veins and pterostigma dark brown.
Body largely glabrous, sparsely covered with minute, appressed, simple yellowish to whitish hairs except as follows: mesoscutellum distally, mesepisternum ventrally, and meso- and metacoxae with long (about as long as median ocellar diameter), stout hairs; posterior margin of metatibia, sides of T6, and distal margin of T7 with long (at least as twice as long as median ocellar diameter), stout hairs.
Material examined. Mexico: 1♂, Jalisco, Careyes, 12 Feb-19 Mar 1997, F.D. Parker (BBSL).
Lestrimelitta danuncia Oliveira & Marchi
Lestrimelitta danuncia Oliveira & Marchi, 2005: 1 [♀].
Diagnosis. Among Central American Lestrimelitta, the female of this species can be easily recognized by the combination of: vertex with erect hairs present on preoccipital border and anterior margin of mesoscutum, disc of mesoscutum practically glabrous, and mesotibial spur long. The male can be recognized by the combination of: mesotibial spur long, and mesoscutellum with long (1.5–1.7 times median ocellar diameter), relatively dense, stout hairs on distal margin.
Description. ♂. As in L. chamelensis except as follows: Total body length 6.0 mm; forewing length 4.3 mm; head width 2.1 mm. Head 1.3 times broader than long (Fig. 25); malar area about one-third width of median ocellar diameter; interalveolar distance slightly shorter than alveolar diameter; alveolocular distance 1.5 times interalveolar distance; interocellar distance 1.8 times median ocellar diameter; scape 5.2 times longer than wide; compound eye 2.4 times longer than broad; gena about 0.6 times width of compound eye in profile. Propodeal spiracle elongate, about six times longer than wide; mesotibial spur long, about 1.5 times mesobasitarsal width basally. T5 apical margin not invaginated medially; S5 and S6 as in figures 32 and 35, genital capsule and S7 as in L. opita.
Color predominantly dark reddish-brown to black, except yellowish on clypeus distally, inferior paraocular area, labrum, anterior surface of scape, mandible, pronotum, legs, and sterna.
Body largely glabrous, sparsely covered with minute, appressed, simple yellowish to whitish hairs except as follows: inferior margin of mandible with long (at most as long as median ocellar diameter), stout hairs; distal margin of clypeus with longer hairs than on mandible (longest hairs about 1.4 times median ocellar diameter); mesoscutellum distally with long (1.8 times median ocellar diameter), stout, black hairs; mesepisternum ventrally and meso- and metacoxae with long (slightly longer than median ocellar diameter), stout hairs; posterior margin of metatibia, sides of T6, and distal margin of T7 with long (two times or more median ocellar diameter), stout, dark brown to black hairs.
Material examined. Costa Rica: 3♂4♀, Guanacaste, Finca Montezuma, 3 km SE Rio Naranjo, March, Apr, Jun, Jul, Oct 1992, Jul 1993, F. Parker (BBSL); 5♀, Guanacaste, Santa Rosa National Park, Oct 1983, 300 m, D.H. Janzen & W. Hallwachs (USNM); 1♀, Heredia, Chilamate, 75 m, Sep-Oct 1989, Hanson & Godoy (BBSL); 1♀, Heredia, LaSelva Res. Sta., 11–17 Jun 1986, W. Hanson, G. Bohart (BBSL); 1♂, Limon, 4 km NE Bribri, 50 m, Sep-Nov 1989, P. Hanson (BBSL); 1♀, Puntarenas, Quepos, 13 Feb 1987, G. E. Bohart (BBSL).
Comments. This species was previously known from eastern Panama ( Panama Province) and central Costa Rica (Turribalba). The specimens from the Costa Rican province of Guanacaste represent the northwestern most distribution of this species.
Lestrimelitta mourei Oliveira & Marchi
Lestrimelitta mourei Oliveira & Marchi, 2005: 2 [♀].
Diagnosis. The female of L. mourei is most similar to that of L. niitkib from Mexico, which shares the following characters: vertex, preoccipital border, and anterior margin of mesoscutum with erect hairs; disc of mesoscutum densely covered by short, fine hairs giving a tomentose appearance; and mesotibial spur reduced. It differs from that species in anterior margin of mesoscutum with longer erect hairs, disc of mesoscutum without erect hairs, disc of mesoscutellum with shorter, sparser erect hairs. The male is distinctive for its metatibia, which is about 2.6 times longer than broad, with outer surface distinctly flattened and the anterior margin strongly convex (Fig. 30).
Description. ♂. As in L. chamelensis except as follows: Total body length 7.2 mm; forewing length 4.4 mm; head width 2.2 mm. Clypeus 2.3 times broader than long; interalveolar distance slightly shorter than alveolar diameter; alveolocular distance about 1.6 times alveolar diameter and interalveolar distance individually; interocellar distance about twice as long as median ocellar diameter; compound eye 2.4 times longer than broad; gena about 0.8 times width of compound eye in profile. Propodeal spiracle elongate, 4.6 times longer than wide; mesotibial spur very reduced, with apex barely visible (c.f. Gonzalez et al. 2010; fig. 3E); metatibia 2.6 times longer than broad, with outer surface distinctly flattened, anterior margin strongly convex (Fig. 30). T5 apical margin not invaginated medially; S5 and S6 as in figures 33 and 36, genital capsule and S7 in general as in L. opita but with longer hairs on gonostylus.
Body with minute, appressed, simple yellowish to whitish hairs denser and longer than in L. chamelensis; distal margin of mesoscutellum with darkened hairs; posterior margin of metatibia, sides of T6, and distal margin of T7 with stout, black hairs about 1.5 times median ocellar diameter.
Material examined. Costa Rica: 2♂, 3♀, Guanacaste, Finca Montezuma, 3 km SE Rio Naranjo, Apr, May 1992, Jun, Aug 1993, F. Parker (BBSL); 4♀, Guanacaste, 14 km S Cañas, Jan 1989, Mar, Apr 1990, F. D. Parker (BBSL); 1♀, Guanacaste, LaTaboga For.Res., 9 km SW Cañas, 23–30 Apr 1987, W. L Rubink (BBSL); 1♀, Limon, Guacimo, 4–7 Mar 1988, F. D. Parker (BBSL); 1♀, Puntarenas, Rincon de Osa, 31 Jul 1966, D. Veirs (BBSL); 1♂, San Jose, Escazu, 18 Jun–3 Jul 1988, F. D. Parker (BBSL); 1♀, San Jose, Escazu, 8 Feb 1987, G. E. Bohart (BBSL).
Comments. The material examined from northwestern Costa Rica ( Guanacaste Province) expands the distribution of this species from its previously known range in central Costa Rica ( San Jose Province).
Lestrimelitta glabatra Camargo & Moure
Lestrimelitta glabatra Camargo & Moure, 1989: 199 [♀].
New Record. Ecuador: 1♀, Napo Province, Huahua Sumaco, km 45 on Hollin-Loreto road, XII-20-1989, Malaise trap, M & J. Wasbauer, collrs. (BBSL); 1♀, idem; Res. Ethnica Waorani, 1 km S. Onkone Gare Camp, Trans. Ent. 6 Oct. 1995, 220m, 00°38'S, 076°36'W, T. L. Erwin, et al / Insecticidal fogging of mostly bare green leaves, some with covering of lichenous or brypothytic plants Project MAXUS At s-trans 6...5, Lot 1225 (USNM); 1♀, idem; 2 July, 1995 / At s-trans 7...9, Lot 1069 (USNM).
Comments. This new record expands the distribution of this species from its previously known range: Brazil and Guyana.
Lestrimelitta spinosa Marchi & Melo
Lestrimelitta spinosa Marchi & Melo, 2006: 25 [♀].
New Record. Venezuela: 1♀, T.F. Amaz. [Territorio Federal, Amazonas] Cerro de la Neblina basecamp, 140 m, 0°50'N, 66°10'W1, 10-20 February 1985 / Malaise trap in rainforest; P.J. & P.M. Spangler, R.A. Faitoute & W.E. Steiner colrs (USNM).
Comments. This species was previously known from Trinidad and Tobago and the Brazilian Amazon; this new record is from a locality intermediate between these two areas.
Keys to workers of Lestrimelitta from Central America and Northern South America
Note: Some characters used in the keys have been recently illustrated elsewhere and are not duplicated here. When necessary, a reference to those works and their corresponding figures are indicated in the couplets.
1. Propodeal spiracle ovoid, 2–3 times longer than broad (c.f. Marchi & Melo 2006; fig. 1)............................................................ 2
-. Propodeal spiracle elongate, at least 4.6 times longer than broad (c.f. Marchi & Melo 2006; fig. 2) ........................... 6
2(1). Lateral surface of mesepisternum practically glabrous, without dense, stout, erect simple hairs ........................ 3
-. Lateral surface of mesepisternum distinctly covered by stout, erect, simple hairs (c.f. Marchi & Melo 2006; fig. 14)............................................................. 4
3(2). Pronotal lobe and anterior margin of mesoscutum practically glabrous, without dense, stout, erect simple hairs (Fig. 12) ............................... L. opita sp. n.
-. Pronotal lobe and anterior margin of mesoscutum distinctly covered by stout, long (1–1.4 times median ocellar diameter), erect, simple hairs.......................................................L. limao (Smith)
4(3). Vertex with few, sparse, short erect hairs restricted to interocellar area; disc of mesoscutum practically glabrous, not densely covered by short, fine hairs, without a tomentose appearance; Trinidad and Tobago, Venezuela, Brazil................................ L. spinosa Marchi & Melo
-. Vertex and preoccipital border with longer, denser hairs throughout; disc of mesoscutum densely covered by short, fine hairs giving a tomentose appearance from which sparse, long, stout hairs arise............................................................. 5
5(4). Mesotibial spur very reduced, apex barely visible; Bolivia, Brazil, Ecuador......................................... L. rufa (Friese)
-. Mesotibial spur normal, about as long as or slightly shorter than mesobasitarsus width basally; Colombia........................ L. piedemontana Gonzalez & Rasmussen
6(1). Vertex, preoccipital border, and anterior margin of mesoscutum practically glabrous, without erect hairs, at most with sparse, short hairs restricted to interocellar area and anterolateral corner of mesoscutum; disc of mesoscutum practically glabrous, with minute, appressed hairs barely visible............. 7
-. Vertex, preoccipital border and anterior margin of mesoscutum with long, erect hairs; disc of mesoscutum with variable pubescence, practically glabrous or densely covered by short, fine hairs giving a tomentose appearance from which sparse, long, stout hairs usually arise.... 10
7(6). Small bees (body length 5 mm; head width 2.2 mm); mesoscutellum distally with short (about twice as long as median ocellar diameter), erect hairs; mesotibia with few long, stout, semierect or erect hairs along anterior and posterior margins............................................................. 8
-. Larger bees (body length 6–7.4 mm; head width at least 2.3 mm); mesoscutellum distally with long (at least three times as long as median ocellar diameter), erect hairs; mesotibia distinctly covered by long (about as long as median ocellar diameter or longer), erect, stout hairs............... 9
8(7). Lateral surface of propodeum covered by relatively long and dense pubescence, integument visible among hairs; mesotibial spur very reduced; Mexico............ L. chamelensis Ayala
-. Lateral surface of propodeum sparsely covered by fine, short hairs (hairs barely visible); mesotibial spur long, normal; Brazil, French Guyana..................................... L. glaberrima Oliveira & Marchi
9(7). Anterolateral corner of mesoscutum with abundant, long, erect hairs; pronotal lobe with long, erect hairs inferiorly, with branched hairs relatively long and dense on posterior margin; mesotibial spur very reduced; T2–T5 laterally with long hairs (at least 1.3 times median ocellar diameter); body length ~ 6 mm; head width 2.3 mm; French Guyana.......................................... L. guyanensis Roubik
-. Anterolateral corner of mesoscutum and pronotal lobe distinctly glabrous, at most with very short, sparse hairs; mesotibial spur normal; T2–T6 with shorter hairs (about as long as median ocellar diameter), progressively increasing in length towards apical terga; body length ~ 7.3 mm; head width 2.6 mm; Brazil, Guyana, Ecuador............ L. glabrata Camargo & Moure
10(6). Mandible with a single, small basal tooth; mesepisternum laterally with erect hairs, hairs longer near omaulus, below pronotal lobe; propodeum laterally with long, branched hairs; South America ........ L. monodonta Camargo & Moure
-. Mandible with two small basal teeth; mesepisternum glabrous; propodeum laterally with short, fine hairs............. 11
11(10). Metasoma with erect hairs on all terga, at least laterally, except on T1; South America................................ 12
-. Metasoma with erect hairs on apical terga only; Central America .................. 13
12(11). Head and mesosoma predominantly dark ferruginous (Figs. 1–3); disc of mesoscutellum largely glabrous (Fig. 3); sides of T2 with long hairs (about as long as median ocellar diameter) (Fig. 4); Venezuela ................ L. catira sp. n.
. Head and mesosoma much darker, predominantly dark brown to black (Figs. 5, 6); disc of mesoscutellum sparsely covered with erect, short (about as long as median ocellar diameter) hairs (Fig. 8); sides of T2 with minute erect hairs (at most half width of median ocellar diameter) (Fig. 7); Colombia........................................ L. huilensis sp. n.
13(11). Mesotibial spur normal, long; disc of mesoscutum practically glabrous, with fine, minute hairs barely visible; erect hairs on anterolateral corner of mesoscutum distinctly longer and denser than those along anteromedial margin.......... L. danuncia Oliveira & Marchi
-. Mesotibial spur reduced; disc of mesoscutum more densely covered by short, fine hairs giving a tomentose appearance; erect hairs on anterolateral corner of mesoscutum about as dense as and about as long as those along anteromedial margin......................... 14
14(13). Discs of mesoscutum and mesoscutellum with erect hairs arising from short, fine pubescence, longer and denser on scutellum; Mexico................................................... L. niitkib Ayala
-. Discs of mesoscutum and mesoscutellum practically without erect hairs arising from short, fine pubescence, at most with few, short, erect hairs on scutellum; Costa Rica................................................... L. mourei Oliveira & Marchi
Key to males of Lestrimelitta from Central America and Northern South America
Note: Males of L. catira, L. guyanensis, L. monodonta, L. huilensis, L. niitkib, and L. piedemontana are unknown.
1. Propodeal spiracle ovoid, 2–3 times longer than broad (c.f. Marchi & Melo 2006; fig. 1); T5 apical margin distinctly invaginated medially.......................... 2
-. Propodeal spiracle elongate, at least 5 times longer than broad (c.f. Marchi & Melo 2006; fig. 2); T5 apical margin not invaginated medially.............................. 5
2(1). Vertex and preoccipital border with relatively dense erect hairs; disc of mesoscutum densely covered by short, fine hairs giving a tomentose appearance from which sparse, long, stout hairs arise; Bolivia, Brazil, Ecuador............................................................ L. rufa (Friese)
-. Vertex practically glabrous, at most with few erect hairs restricted to interocellar area; disc of mesoscutum practically glabrous, with minute, appressed hairs barely visible...................................... 3
3(2). Lateral surface of mesepisternum distinctly covered by stout, erect, simple hairs (c.f. Marchi & Melo 2006; fig. 14); Trinidad and Tobago, Venezuela, Brazil................... L. spinosa Marchi & Melo
-. Lateral surface of mesepisternum practically glabrous, without dense, stout, erect simple hairs................................ 4
4(3). Metafemur and metatibia covered by long (1–2 times median ocellar diameter), dense, stout erect hairs (Fig. 15)................................................. L. opita sp. n.
-. Metafemur and metatibia covered by much shorter (at most as long as median ocellar diameter) erect hairs............................................... L. limao (Smith)
5(1). Mesotibia with long and dense erect hairs; metasoma with erect hairs on all terga, at least laterally, except on T1........................... L. glabrata Camargo & Moure
-. Mesotibia without long and dense erect hairs, practically glabrous or with barely visible minute, fine appressed hairs; metasoma with erect hairs on apical terga only..................................................... 6
6(5). Metatibia 2.6 times longer than broad, with outer surface distinctly flattened and anterior margin strongly convex (Fig. 30).............. L. mourei Oliveira & Marchi
-. Metatibia 2.7–3.0 times longer than broad, with outer surface gently convex, not distinctly flattened, its anterior margin gently convex (Figs. 28, 29)............... 7
7(6). Mesotibial spur long, normal; mesoscutellum with long (1.5–1.7 times median ocellar diameter), relatively dense, stout hairs on distal margin................... L. danuncia Oliveira & Marchi
-. Mesotibial spur reduced; mesoscutellum practically glabrous or with few, relatively sparse, short hairs (at most as long as median ocellar diameter) on distal margin........................................................... 8
8(7). Mesoscutellum with few, sparse, short hairs (at most as long as median ocellar diameter) on distal margin; metatibia with sparse, rather long hairs (about as long as median ocellar diameter or longer) on outer surface (Fig. 28); Mexico....................... L. chamelensis Ayala
-. Mesoscutellum practically glabrous; metatibia with few, short hairs (at most 0.6 times median ocellar diameter) on outer surface, near apex; Brazil, French Guyana............................................. L. glaberrima Oliveira & Marchi
ACKNOWLEDGMENTS
We are grateful to B. Harris and S. Brady (USNM) and C. Sarmiento (ICN) for bringing this material to our attention and permitting its study; Orlando Rangel for the invitation to contribute to this volume; Michael S. Engel, Amy Comfort de Gonzalez, Molly Rightmyer, and anonymous reviewers for their comments and suggestions that improved this manuscript. Support to V.H.G. was provided by US National Science Foundation grant DBI-1057366 (to Michael S. Engel). This is a contribution of the Division of Entomology, University of Kansas Natural History Museum.
LITERATURE CITED
1. ASCHER, J. & J. PICKERING. 2011. Bee Species Guide (Hymenoptera: Apoidea: Anthophila). http://www.discoverlife.org/mp/20q?guide=Apoidea_species. last accessed 11September 2011. [ Links ]
2. AYALA, R. 1999. Revisión de las abejas sin aguijón de México (Hymenoptera: Apidae: Meliponini). Folia Entomológica Mexicana 106: 1–123. [ Links ]
3. BÄNZIGER, H., BOONGIRD, S., SUKUMALANAND, P. & BÄNZIGER, S. 2009. Bees (Hymenoptera: Apidae) that drink human tears. Journal of the Kansas Entomological Society 82(2): 135–150. [ Links ]
4. BOONGIRD, S. & C. D. MICHENER. 2010. Pollen and propolis collecting by male stingless bees (Hymenoptera: Apidae). Journal of the Kansas Entomological Society 83(1): 47–50. [ Links ]
5. CAMARGO, J. M. F. & J. S. MOURE. 1989. Duas espécies novas de Lestrimelitta Friese (Meliponinae, Apidae, Hymenoptera) da Região Amazônica. Boletim do Museu Paraense Emílio Goeldi, Série Zoologia 5(2): 185–212. [ Links ]
6. CAMARGO, J. M. F. & D. W. ROUBIK. 1991. Systematics and bionomics of the apoid obligate necrophages: the Trigona hypogea group (Hymenoptera: Apidae; Meliponinae). Biological Journal of the Linnean Society 44: 13–39. [ Links ]
7. CAMARGO, J. M. F. & S. R. M. PEDRO. 2002. Mutualistic association between a tiny Amazonian stingless bee and a wax-producing scale insect. Biotropica 34(3): 446–451. [ Links ]
8. CAMARGO, J. M. F. & S. R. M. PEDRO. 2007a. Notas sobre e bionomía de Trichotrigona extranea Camargo & Moure (Hymenoptera, Apidae, Meliponini). Revista Brasileira de Entomologia 51(1): 72-81. [ Links ]
9. CAMARGO, J. M. F. & S. R. M. PEDRO. 2007b. Meliponini Lepeletier, 1836. In: Moure, J. S., D. Urban., & G. A. R. Melo (Orgs). Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Available at http://www.moure.cria.org.br/catalogue [last accessed 11September 2011 [ Links ]
10. ENGEL, M. S. & V. H. GONZALEZ. 2009. A new species of Chlerogas from the Andes of central Colombia (Hymenoptera: Halictidae). Caldasia 31(2): 441–447. [ Links ]
11. ENGEL, M. S. 1997. Ischnomelissa, a new augoclorine bee genus (Halictidae) from Colombia. Studies on Neotropical Fauna and Environment 32(1): 41–46. [ Links ]
12. ENGEL, M. S. 2009. Notes on the augochlorine bee genus Chlerogas (Hymenoptera: Halictidae). Caldasia 31(2): 449–457. [ Links ]
13. ENGEL, M. S. 2010. Revision of the bee genus Chlerogella (Hymenoptera, Halictidae), part II: South American species and generic diagnosis. ZooKeys 47: 1–100. [ Links ]
14. FRIESE, H. 1903. Neue Meliponiden II. Zeitschrift fü r Systematische Hymenopterologie und Dipterologie 3: 359–361. [ Links ]
15. GONZALEZ, V. H. 2004. A new species of Acamptopoeum from Colombia (Hymenoptera: Andrenidae: Panurginae). Caldasia 26(1): 239–243. [ Links ]
16. GONZALEZ, V. H. 2006. Dos especies nuevas de abejas de la ciudad de Bogotá ( Colombia ). Revista Colombiana de Entomología 32(1): 93–96. [ Links ]
17. GONZALEZ, V. H. & M. S. ENGEL. 2004. The tropical Andean bee fauna (Insecta: Hymenoptera: Apoidea), with examples from Colombia. Entomologische Abhandlungen 62(1): 65–75. [ Links ]
18. GONZALEZ, V. H. & L. RUZ. 2007. New enigmatic Andean bee species of Protandrena (Hymenoptera, Andrenidae, Panurginae). Revista Brasileira de Entomologia 51(4): 397–403. [ Links ]
19. GONZALEZ, V. H. & M. S. ENGEL. 2011. Andinopanurgus, a new Andean subgenus of Protandrena (Hymenoptera, Andrenidae). ZooKeys 126: 57–76. [ Links ]
20. GONZALEZ, V. H. & M. S. ENGEL. 2012. A new species of Geotrigona Moure from the Caribbean coast of Colombia (Hymenoptera, Apidae). ZooKeys 172: 77–87. [ Links ]
21. GONZALEZ, V. H. & J. FLOREZ. 2011. Leioproctus rosellae sp. n., the first record of the genus from northern South America (Hymenoptera, Colletidae). ZooKeys 141: 71–77. [ Links ]
22. GONZALEZ, V. H. & T. GRISWOLD. 2011. Heriades tayrona n. sp., the first Osmiine bee from South America (Hymenoptera: Megachilidae). Journal of the Kansas Entomological Society 84(4): 255–259. [ Links ]
23. GONZALEZ, V.H., C. RASMUSSEN & A. VELÁZQUEZ. 2010. Una especie nueva de Lestrimelitta y un cambio de nombre en Lasioglossum (Hymenoptera: Apidae, Halictidae). Revista Colombiana de Entomología 36(2): 319–324. [ Links ]
24. LEPELETIER DE SAINT FARGEAU, A. L. M. 1836. Histoire Naturelle des InsectesHyménoptères {vol. 1}. Librairie Encyclopédique de Roret, Paris, 547 pp. [ Links ]
25. MARCHI, P. & G. A. R. MELO. 2006. Revisão taxonômica das espécies Brazileiras de abelhas do gênero Lestrimelitta Friese (Hymenoptera, Apidae, Meliponina). Revista Brasileira de Entomologia 50(1): 6–30. [ Links ]
26. MELO, GAR. 2003. Notas sobre meliponíneos neotropicais, com a descrição de três novas espécies (Hymenoptera, Apidae). In: Melo, G.A.R. & I. Alves-dos-Santos (eds.). Apoidea Neotropica: Homenagem aos 90 Anos de Jesus Santiago Moure. Criciúma, Editora UNESC; pp. 84–91. [ Links ]
27. MICHENER, C.D. 2007. The Bees of the World [2nd Edition]. Johns Hopkins University Press, Baltimore. [ Links ]
28. OLIVEIRA, F.F. 2002. The mesotibial spur in stingless bees: a new character for the systematics of Meliponini (Hymenoptera: Apidae). Journal of the Kansas Entomological Society 75: 194–202. [ Links ]
29. OLIVEIRA, F. F. & MARCHI, P. 2005. Três espécies novas de Lestrimelitta Friese (Hymenoptera, Apidae) da Costa Rica, Panamá e Guiana Francesa. Revista Brasileira de Entomologia 49(1): 1–6. [ Links ]
30. PORTUGAL-ARAÚJO, V. 1958. A contribution to the bionomics of Lestrimelitta cubiceps. Journal of the Kansas Entomological Society 31(3): 203–211. [ Links ]
31. RASMUSSEN, C. & S.A. CAMERON. 2010. Global stingless bee phylogeny supports ancient divergence, vicariance, and long distance dispersal. Biological Journal of the Linnean Society 99: 206–232. [ Links ]
32. ROIG-ALSINA, A. 2010. Notas sistemáticas sobre abejas Meliponini del Chaco (Hymenoptera, Apidae). Revista del Museo Argentino de Ciencias Naturales 12(1): 99–106. [ Links ]
33. ROUBIK, D. W. 1982. Obligate necrophagy in a social bee. Science 217: 1059–1060. [ Links ]
34. SAKAGAMI, S. F., ROUBIK, D. W., & R. ZUCCHI. 1993. Ethology of the robber stingless bee, Lestrimelitta limao. Sociobiology 21(2): 237–277. [ Links ]
35. SMITH-PARDO, A. & V. H. GONZALEZ. 2007. Abejas Chilicola (Hymenoptera, Colletidae, Xeromelissinae) del grupo megalostigma: una nueva especie de Colombia y clave para las especies. Neotropical Entomology 36(6): 910–913. [ Links ]
36. SMITH-PARDO, A. H. & V. H. GONZALEZ. 2009. A revision of Neocorynura bees of the joannisi group, with new geographical records for other rare Andean species (Hymenoptera: Halictidae, Augochlorini). Studies on Neotropical Fauna and Environment 44(2): 115–129. [ Links ]

