










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.34 no.1 Bogotá Jan./June 2012
ALFONSO N. GARCÍA-ALDRETE
Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 70-153, 04510 México, D. F. México. anga@ibiologia.unam.mx
ABSTRACT
A monotypic genus of Epipsocidae is here described and illustrated. It belongs in the clade Phallofractus-Goja-Rogojiella-Ianthorntonia and shows several striking autapomorphic characters. Goja Navás is divided in two genera on basis of the male wing venation. The type of the Colombian new genus is deposited in the Entomological Museum of the Universidad del Valle, Santiago de Cali, Colombia.
Key words. Taxonomy, neotropics, endemic, Epipsocetae.
RESUMEN
Un nuevo género monotípico de Epipsocidae es descrito e ilustrado en este trabajo. Pertenece al clado Phallofractus-Goja-Rogojiella-Ianthorntonia y presenta varios caracteres autapomórficos notables. Goja Navás es dividido en dos géneros con base en la venación de los machos. El tipo del nuevo género colombiano está depositados en el Museo Entomológico de la Universidad del Valle, en Santiago de Cali, Colombia.
Palabras clave. Taxonomía, neotrópico, endémico, Epipsocetae.
Recibido: 11/10/2011
Aceptado: 20/04/2012
INTRODUCTION
Casasola González (2006) studied the phylogenetic relationships of the genera in the psocopteran infraorder Epipsocetae, that includes the families Dolabellopsocidae, Ptiloneuridae, Spurostigmatidae, Cladiopsocidae and Epipsocidae; in the strict consensus tree of 104 most parsimonious trees (Fig. 124), a clade was recognized, for the latter family, including the genera sp4 THAI, Goja, Rogojiella and Ianthorntonia. The former genus was later described as Phallofractus (García Aldrete, 2009). In this clade, the males have either caeciliusid venation, with Rs-M in the hindwing fused or joined by a crossvein, or they have supernumerary venation on both wings; in these genera, the phallosome is complex, in most cases with two pairs of endophallic sclerites. Phallofractus is Oriental; it is known only from one male; the arms of the aedeagal arch are separated and the phallosome has only one pair of endophallic sclerites (García Aldrete 2009), it is believed to constitute the sister group of the other three genera, which are neotropical (Casasola González & García Aldrete 2002, García Aldrete 2004 a and b). The females of Goja and Ianthorntonia (female unknown in Rogojiella), are either brachypterous or apterous, with v1 directed posteriorly from their junction with the clunium. In both sexes, the head is not decidedly elongate, the compound eyes, in most of the species do not protrude beyond the level of the vertex, and the outer cusp of the lacinial apex is diagonal.
Recently, Ranulfo González Obando and Nancy Soraya Carrejo, of the Biology Department, Universidad del Valle, Santiago de Cali, Colombia, showed to me some specimens collected by them near Cali, in an area of cloud forest. These specimens clearly belong in the clade indicated above, but they are not assignable to any of the genera of the clade, thus representing an undescribed genus; the purpose of this work is to describe it and illustrate it, and to assess the relationships with the other genera in the clade; also, the genus Goja Navás is re-evaluated, and a new genus is erected from a fraction of it.
MATERIAL AND METHODS
28 males and 12 females were available for study. Two specimens of each sex were dissected in 80% ethyl alcohol, and their parts (head, right wings and legs and genitalia) were mounted on slides in Canada Balsam. Color was recorded by placing whole specimens in 80% ethyl alcohol under a dissecting microscope, illuminated with white, cold light, at 80X. Measurements, in µm, of parts on the slides, were taken with an AO filar micrometer. Abbreviations of parts measured, or counted, are as follows: FW and HW:
lengths of fore- and hind- wings, F, T, t1 and t2: lengths of femur, tibia and tarsomeres 1 and 2 of right hind leg, ctt1: number of ctenidobothria on t1, Mx4: length of fourth segment of right maxillary palp, IO, D and d respectively: minimum distance between compound eyes, antero-posterior diameter and transverse diameter of right compound eye in dorsal view of head, PO: d/D. The types are deposited in the Entomological Museum of the Universidad del Valle, Santiago de Cali, Colombia (MUSENUV). One paratype of each sex is deposited in the National Collection of Insects (CNIN), Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México, México City.
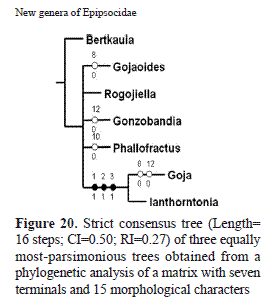
To assess the relationship of the new genera in the clade with the other genera, a matrix was built utilizing the 15 characters shown in Table 1; in the matrix (Table 2), Bertkauia was the outgroup. Goja was split in two groups: Goja, for species with males having supernumerary venation in both wings, and Gojaoides gen. n., for species with males having caeciliusid wing venation (see section on re-appraisal of Goja below). A parsimony analysis using NONA (Goloboff, 1999), was conducted. Three trees were generated utilizing an heuristic search. The characters parsimony informative, were treated initially as unweighted and unordered; the trees were rooted by the outgroup method; the characteristics of the strict consensus tree are the following: L= 16 steps, consistency index (CI)= 0.50, and retention index (RI)= 0.27.


RESULTS
Gonzobandia gen. n.
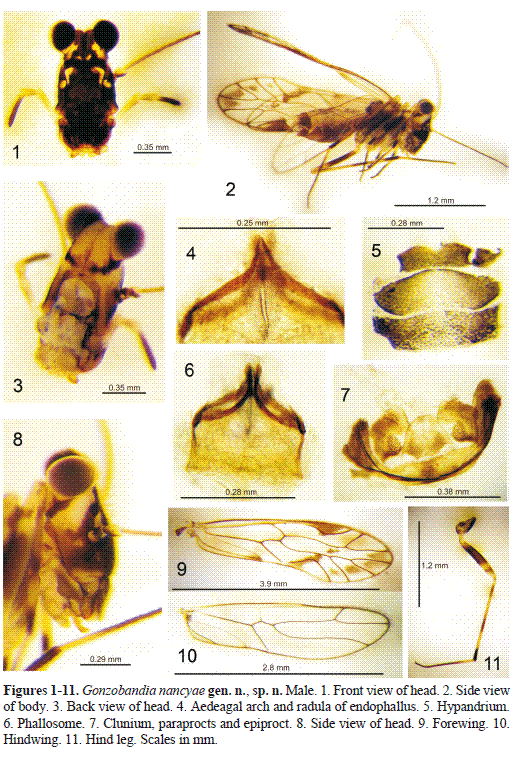
Diagnosis. Belonging in the Epipsocidae. Head long, slender; lower genae protruding laterally, at the level of the subgenal sulcus (Figs. 1, 3, 12).Compound eyes prominent, extending well beyond the level of the vertex (Figs. 1, 3, 8, 12). Outer cusp of lacinial apex straight, with seven denticles. Distal inner labral sensilla: one central placoid, flanked at a distance by a pair trichoid-placoid. Male wing venation caeciliusid (Figs. 9, 10). Phallosome simple, open anteriorly, with aedeagal arch pointed and external parameres slender, incurved, bearing pores distally; lacking endophallic sclerites, but with a radula slit in the middle (Figs. 4, 6). Female brachypterous, only with forewings (Figs. 13, 17); gonapophyses (Fig. 14), with v1 proximally bent at an angle of 90°.

Genus type. Gonzobandia nancyae gen. n., sp. n.
Gonzobandia nancyae sp. n.
Holotype. Male. COLOMBIA. Valle del Cauca. Santiago de Cali, Pueblo Nuevo, Vereda Los Andes, Finca Montserrate, 3°25'57.3"N: 76°37'15.4"W, 1682 m., 18.I.2011. On tree trunks densely covered with mosses, R. González Obando & N. S. Carrejo.
Paratypes. 6 males, 5 females, collected with the holotype. 2 males, 1 female, same locality, Vereda La Candelaria, 3°22'04.6"N: 76°37'58"W, 1740 m., R. González & J. Mendívil; 1 male, 6 females, same locality, Vereda Los Andes, 15.VI.2011, R. González; 7 males, same locality, Vereda Los Andes, 4.VII.2011, R. González & J. Mendívil; 4 males, same locality, 4.IX.2011, R. González.
Other records. COLOMBIA. Valle del Cauca. La Cumbre, Chicoral, Finca Loma Linda, 3°34'18.4"N: 76°35'36.7"W, 1920 m., 26.III.2011, 1 male, J. Mendívil. Same locality, 3°34'30.3"N: 76°35'36.2"W, 1722 m., 29.VI.2011, 5 males, R. González & J. Mendívil. Yumbo, Dapa, Finca Palo Alto, 3°34'09.0"N: 76°34'13.6"W, 1866 m., 10.IX.2011, 1 male, J. A. Mendívil.
Description. Male. Color (in 80% alcohol). Body creamy white, with dark brown areas as indicated below. Head (Figs. 1,3, 8), chocolate brown, with creamy areas as illustrated: a large one next each compound eye, two small in front, and a median one bordering epistomal sulcus, next each antennal fossa; a brown band behind, from lower end of each compound eye to upper border of occipital foramen (Fig. 3). Maxillary palpomeres 1 and 2 whitish, 3 pale brown, and 4 dark brown. Antennae: scape and pedicel brown, f1 with two proximal thirds pale brown, f2-f11 dark brown. Legs (Fig. 11): coxa brown, trochanter pale brown, femur brown, with proximal end pale brown and distal third pale brown, with a brown irregular band in the middle. Tibia with three alternating creamy and brown bands, proximally creamy and distally dark brown; t1 proximally creamy, distally pale brown, t2 brown. Forewing with membrane almost hyaline, opaque, with brown spots on lower apex of pterostigma, mesally and distally on cell R-M, distally on cells M, and diagonal from anterior half of areola postica to touch M; confluence of Cu2-A brown; pterostigma brown, distally with a hyaline, transverse band; veins brown (Fig. 9). Hind wing almost hyaline, with a pale brown hue, veins brown (Fig. 10). Thorax deep chocolate brown. Abdomen reddish brown, with a whitish longitudinal band on each side, between tergum and sternum, enclosing the spiracles.
Morphology. As in generic diagnosis, plus the following: Head (Figs. 1, 2), widening in front view from compound eyes to subgenal sulcus. Ocelli absent. Lacinial tip with inner cusp acuminate, with a small tooth next to it. Forewing (Fig. 9) long, slender, narrow, pterostigma narrow anteriorly, wider in the middle. Rs almost straight, about twice as long as R4+ 5. M almost straight, about 1.5 times the length of M3. Areola postica wide, low, triangular. Hindwing (Fig. 10), long, slender, narrow, with Rs-M fused for a distance. Hypandrium (Fig. 5) of two pieces, the posterior one smaller, with a field of setae on surface and a field of five macrosetae on each postero-lateral corner. Phallosome (Figs. 4, 6), side struts short, slender, joined distally to proximal ends of aedeagal arch, this projected posteriorly, apex acute; external parameres arising from junction of side struts and base of aedeagal arch, slightly dilated distally (Fig. 6). Endophallus radular (Fig. 6). Paraprocts (Fig. 7) broad, setose, with setal field apically and a field of microspines along posterior border. Sensory fields mesal, slightly elongate, with 27-30 trichobothria on basal rosettes. Epiproct (Fig. 7) slightly convex anteriorly, almost trapeziform, with three setae mesally, a field of setae on posterior third and a field of microsetae along posterior border.
Measurements. FW: 3944, HW: 2830, F: 1037, T: 1224, t1: 802, t2: 175, ctt1: 49, Mx4: 351, f1: 869, f2: 727, f3: 582, f4: 408, IO: 111, D: 292, d: 235, IO/d: 0.47, PO: 0.80.
Female. Color (in 80% alcohol). Body same as the male. Head pattern: a broad brown band on vertex, between compound eyes, creamy areas below as illustrated (Fig. 12). Brown bands next to lower ends of compound eyes, directed backwards but not reaching the occipital foramen. Maxillary palpomeres: 1 to proximal end of 4 creamy white, rest of 4 dark brown. Antennae as in the male, slightly less pigmented. Legs (Fig. 19), as in the male, only with coxa and trochanter creamy white, and outer apex of coxa dark brown. Genital segments brown. Thorax pale brown. Abdomen reddish brown; a longitudinal whitish band on each side, as in the male (Fig. 13).
Morphology. Brachypterous, only with forewings. Ocelli absent. Compound eyes and lacinial tip as in the males. Forewing (Fig. 17) broad, margin setose, venation reduced. Subgenital plate (Fig. 16 ) broad, obtusely convex posteriorly, pigmented area anteriorly concave. Gonapophyses (Fig. 14): v1 short, slender, v2+3 with slender anterior heel, with a field of seven setae longitudinally, v2 lobe not well defined, distal projection pointed, with field of microsetae distally on inner edge. Ninth sternum broadly elliptic, with a sclerotized peripheral rim; spermapore small, central, with elongate, ochre pigmented area surrounding it (Figs. 14-15). Paraprocts (Fig. 18) long, broadly conical, setose, sensory fields with 4-5 trichobothria, one in a large rosette, and 3-4 issuing from small rosettes. Epiproct (Fig. 18), long, convex anteriorly, distally blunt, setose, with three mesal setae anteriorly.
Measurements. FW: 741, F: 952, T: 1611, t1: 988, t2: 250, Mx4: 335, f1: 599, f2: 496, f3: 408, f4: 279, IO: 110, D: 272, d: 235, IO/d: 0.46, PO: 0.86.
Etymology. The genus name is a compound noun, of female gender, formed with the two last names of Ranulfo González Obando, of the Universidad del Valle, Santiago de Cali, Colombia. The species name derives from the first name of Nancy Soraya Carrejo, of the same institution. Gonzobandia nancyae is dedicated to them, in recognition to their strong and energetic impulse to survey and study the Psocoptera of Colombia, mostly of Valle del Cauca, Cauca and Caldas.
The position of Gonzobandia in the clade
The strict consensus tree is presented in Fig. 20, the relationships of the genera in the clade are unresolved, except for Goja and Ianthorntonia; the clade is monophyletic, supported by two synapomorphies: endophallus radular or with sclerites (5-1), and external parameres membranous or well defined projections (7-1).
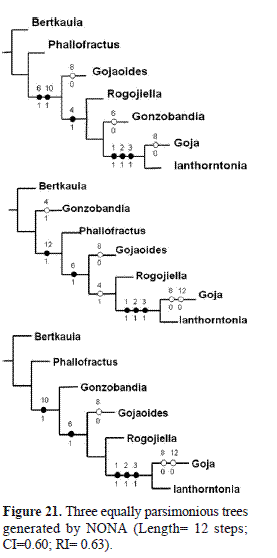
On basis of the three most parsimonious trees found (Fig. 21), two hypotheses are presented, as to whether Phallofractus or Gonzobandia constitute the sister group to the other genera in the clade; on geographical grounds, it would seem sensible to consider the former as the sister group to the rest of the assemblage, and it so appears in two of the three most parsimonious trees. In one of three trees, Gonzobandia is the sister group to the rest of the assemblage. The position of Gojaoides as sister group to Rogojiella-Goja –Ianthorntonia, in two of the trees is also sensible, (Fig. 21), as the latter pair shares having supernumerary venation in the male forewing, and Rogojiella, as well as Gojaoides, have caeciliusid wing venation. It would appear, to resume the above, that the third most parsimonious tree found, best represents the relationships of the genera in the clade.
A re-appraisal of Goja Navás, 1927
The species presently recognized in Goja Navás, 1927 (see Casasola González & García Aldrete, 2002), are assignable in two well defined groups, based on male wing venation characters, as follows:
Group I. Male forewing venation supernumerary, in the branches of Rs, M, or both. The hindwing venation may be either caeciliusid, or with Rs 2-branched and M supernumerary, or with both Rs and M with supernumerary veins. In all cases, hindwing veins Rs and M are joined by a crossvein.
Group II. Both male wings with venation caeciliusid (forewing Rs 2-branched, M 3-branched; hindwing Rs 2-branched, M simple). In all cases, hindwing veins Rs and M are joined by a crossvein.
In both groups the females are neotenic, lacking ocelli, or these at most represented by light spots, the coxal organ is little developed or absent, and t1 of hind legs lacks ctenidobothria. They are mostly brachypterous or apterous, lacking hindwings. The subgenital plate is simple, setose, and the ovipositor valvulae are complete in all but one species; the ninth sternum is distinct, almost circular, with the spermapore often distal, surrounded or not by a pigmented ring. The epiproct and paraprocts are elongate, with long setae distally; the paraprocts have a small sensory area not well defined, bearing 2-10 trichobothria issuing from basal rosettes, one or two longer than the others; only one species lacks basal rosettes (Casasola González & García Aldrete, 2002).
Also in both groups the males are similar, other than the differences in wing venation, and can be succinctly diagnosed as follows:
Compound eyes normal, occasionally elevated beyond the level of vertex. Ocelli generally conspicuous. Distal inner labral sensilla: one central placoid, flanked at a distance by a close pair trichoid-placoid. Outer cusp of lacinial apex generally broad, with 3-8 denticles; t1 of hind legs with well developed ctenidobothria. Pretarsal claws with one preapical denticle, pulvillus filiform, acuminate. Macropterous, as indicated above. Hypandrium simple, of two parts, the posterior one generally with a median posterior concavity, and side fields of long setae on postero-lateral corners. Phallosome open or close anteriorly; side struts straight or curved, often fused to the aedeagal arch, this with apex variable; external parameres as pore bearing lobes, membranous or slightly sclerotized, partially immersed in the endophallus. One or two pairs of endophallic sclerites, occasionally with one central sclerite. Epiproct trapeziform, with a field of spicules along posterior border, a field of setae on each side, and 1-3 mesal macrosetae, towards anterior border. Paraprocts broad, almost rounded, each with a large sensory field, with 20-42 trichobothria issuing from basal rosettes, occasionally 1-2 central trichobothria without basal rosettes (Casasola González & García Aldrete, 2002).
For Group I, I propose to retain the name Goja Navás, since the type species of the genus (male), Goja ditata Navás, 1927, has both wings with supernumerary venation (see Fig. 9.a in Navás, 1927). The species included are the following:
G. acuphallica, G. brailovskyana, G. circumvittata, G. chilensis, G. curvata, G. eertmoedi, G. guatemalensis, G. herrerae, G. longispinosa, G. martinezi, G. medellini, G. mockfordi, G. montieli, G. moyai, G. obrienorum, G. ochoterenae, G. ocotensis, G. oculatissima, G. omiltemensis, G. poblana, G. reticulata, G. reticulatoides, G. spinosissima, G. ticana, G. venezuelensis, and G. vittata (all by Casasola González & García Aldrete).
Type species: Goja ditata Navás.
The species Goja bimaculata, G. pallida and G. trinidadensis, all by Casasola González & García Aldrete, are only known from females; on the basis of similarities with females of Goja, they are tentatively assigned in that genus. The finding of males of these species will confirm, or reject, this placement.
For Group II, I propose to create the genus Gojaoides gen. n., as diagnosed above, and including the following species:
Gojaoides aviceps (Badonnel), n. comb., G. bogotana (Roesler), n. comb., G. cubitalis (Mockford), n. comb., G. pechi (Williner), n. comb., G. molinai (Williner), n. comb., G. nebulosa (Roesler), n. comb., G. plaumanni (Roesler), n. comb., G. semiaptera (Mockford), n. comb., G. cornuta, G. erwini, G. latiphallica, G. longiareola, G. robusta and G. spiculata (all by Casasola González & García Aldrete).
Type species: Gojaoides picta (Banks), n.comb.
A key to males of the genera in the clade Phallofractus-Ianthorntonia
(females unknown in Phallofractus, Rogojiella and Ianthorntonia)1. Forewing venation caeciliusid................2
-. Forewing venation with supernumerary veins............................................................5
2. Hindwing veins Rs and M joined by a crossvein..........................Gojaoides gen. n.
-. Hindwing veins Rs and M fused.............3
3. External parameres of phallosome membranous lobes; aedeagal arch of two arms, leaving a space at the apex; endophallus with one pair of sclerites.........................................................Phallofractus García Aldrete
-. External parameres of phallosome well sclerotized; aedeagal arch of one piece; endophallus radular or with more than one pair of sclerites...........................................4
4. External parameres slender, bow-shaped, distally rounded; endophallus radular, with a longitudinal slit..........Gonzobandia gen. n.
-. External parameres stout, acuminate, distally curved; endophallus with three pairs of acuminate sclerites..............................................................Rogojiella García Aldrete
5. Hindwing veins Rs and M fused; external parameres of phallosome wide based, narrowing distally, sclerotized.............................................Ianthorntonia García Aldrete
-. Hindwing veins Rs and M joined by a crossvein; external parameres of phallosome membranous lobes or absent.......Goja Navás
DISCUSSION
Several autapomorphies are presented by Gonzobandia (characters 10, 12, 14 and 15 in Table 2, plus the simple phallosome, the endophallic "radula", v1 bent proximally at 90°, and the distally straight outer cusp of the lacinial tip). It differs from the other genera in the clade by the autapomorphies listed above, keeping, in one of three most parsimonious trees found, a sister group relationship with the assemblage Gojaoides-Rogojiella-Goja and Ianthorntonia. Gonzobandia nancyae is a montane species, with distribution probably restricted to a small area in Valle del Cauca, Colombia, it lives on tree trunks densely covered with mosses and runs rapidly when disturbed. The small area of distribution and the peculiar biotope it inhabits, makes it vulnerable with respect to anthropogenic changes of the habitat.
ACKNOWLEDGMENTS
I am most grateful to Ranulfo González Obando and Nancy Soraya Carrejo (Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle, Santiago de Cali, Colombia ), for making available for study this interesting species. I also thank Felipe Villegas Márquez (Instituto de Biología, Universidad Nacional Autónoma de México) for support with the plates of illustrations. Especial thanks are extended to Martín Leonel Zurita García, of the same institution, for advice in cladistics and for running in NONA the matrix of the genera in the clade studied.
LITERATURE CITED
1. CASASOLA GONZÁLEZ, J. A. 2006. Phylogenetic relationships of the genera of Epipsocetae (Psocoptera: Psocomorpha). Zootaxa 1194: 1-32. [ Links ]
2. CASASOLA GONZÁLEZ, J. A. & A.N. GARCÍA ALDRETE. 2002. A taxonomic revision of the genus Goja (Psocoptera: Epipsocidae). Publicaciones Especiales del Instituto de Biología 18. Universidad Nacional Autónoma de México, México, D. F. 143p. [ Links ]
3. GARCÍA ALDRETE, A.N. 2004 a. A new genus of Bolivian Epipsocidae |(Psocoptera). Journal of the New York Entomological Society 112 (4): 217-220. [ Links ]
4. GARCÍA ALDRETE, A.N. 2004 b. A new epipsocid genus (Psocoptera: Epipsocidae) from Bolivia. In: García Aldrete, A. N., C. Lienhard & E. L. Mockford (eds.). Thorntoniana. A commemorative volume for Ian W. B. Thornton. Publicaciones Especiales del Instituto de Biología 20. Universidad Nacional Autónoma de México, México, D. F. 205p. [ Links ]
5. GARCÍA ALDRETE, A.N. 2009. New monotypic genera of Epipsocidae (Psocoptera: Epipsocidae) from Belize and Thailand. Zootaxa 1978: 63-68. [ Links ]
6. GOLOBOFF, P. 1999. NONA (NO NAME), ver. 2. Published by the autor. Tucumán, Argentina. [ Links ]
7. NAVÁS, L. 1927. Comunicaciones Entomológicas. [ Links ]
8. Socópteros del Museo de Hamburgo. Revista de la Academia de Ciencias Exactas, Físico-químicas y Naturales de Zaragoza 11: 37-52. [ Links ]

