










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.35 no.1 Bogotá Jan./June 2013
Phytocenoses in the Mogotes from Gran Meseta de Guantánamo, Eastern Cuba
ORLANDO J. REYES
FÉLIX ACOSTA CANTILLO
Centro Oriental de Ecosistemas y Biodiversidad (BIOECO), Ministerio de Ciencia, Tecnología y Medio Ambiente (CITMA), Santiago de Cuba, Cuba.joel@bioeco.ciges.inf.cu
RESUMEN
Los mogotes de la Gran Meseta de Guantánamo están constituidos por calizas duras de la Formación Charco Redondo. La temperatura del aire varía entre 22 y 24°C y llueve de 1 200 a 1 400 mm al año. Mediante la metodología de la Escuela Zurich - Montpellier se describe un nuevo orden, Coccothrinaci leonis - Guapiretalia rufescentis, con su alianza Coccothrinaci leonis - Guapirion rufescentis y tres asociaciones. Bactrio cubensis - Podocarpodetum ekmanii y Phyllantho epiphyllanthi - Tabebuietum myrtifoliae que se establecen en la parte superior de los mogotes y la asociación Pileo fruticulosae - Thelypteridetum alatae en fragmentos verticales, las dos primeras tienen una bien desarrollada estera radical donde se produce el reciclaje de nutrientes de este ecosistema.
Palabras clave: Mogotes, sintaxonomía, asociaciones vegetales, Cuba Oriental.
ABSTRACT
The Mogotes (karstic dome hill) of Gran Meseta de Guantánamo karst are over Charco Redondo formation. The wheather conditions as Air temperature vary from 22 to 24°C and rainfall from 1 200 to 1 400 mm anually. By using the Zurich – Montpellier school methodology one new orders are described Coccothrinaci leonis - Guapiretalia rufescentis with the aliance Coccothrinaci leonis - Guapirion rufescentis and three asociations. Bactrio cubensis - Podocarpodetum ekmanii and Phyllantho epiphyllanthi - Tabebuietum myrtifoliae occupied the top of mogotes with a well-developed rot mat where this ecosystem's nutrient cycle is concentrated. Pileo fruticulosae - Thelypteridetum alatae grow in vertical fragments.
Key words: Mogotes, sintaxonomy, associations, Eastern Cuba.
Recibido: 20/02/2012
Aceptado: 30/04/2013
INTRODUCCIÓN
Los bosques siempreverdes se presentan en todo el archipiélago cubano, sobre todo en los macizos montañosos (Capote & Berazain 1984, Borhidi 1996, Reyes & Acosta 2005, Reyes 2006), también denominados como bosques subperennifolios (Del Risco 1995) o manacales (León 1946), son bosques que mantienen sus hojas durante el período menos lluvioso. En Sagua Baracoa (sensu Núñez & Viña Bayés 1989), donde se encuentra el área estudiada, el bosque siempreverde es escaso, encontrándose generalmente en lugares relativamente húmedos y son más abundantes en zonas premontanas. Su mayor extensión se observa en la Gran Meseta de Guantánamo y en la parte Oeste de las Cuchillas del Toa. Estos bosques han sido muy alterados por la actividad humana para transformarlos en áreas agropecuarias. Actualmente solo quedan relictos fragmentados, aislados, muchas veces secundarios. Se han diferenciado dos tipos principales: bosque siempreverde mesófilo y bosque siempreverde micrófilo (Capote & Berazain 1984), este último en condiciones de menos humedad y/o nutrientes. Con posterioridad Reyes (2006) caracterizó en la Sierra Maestra el bosque siempreverde micrófilo calcífilo (en sitios cársicos) y el bosque siempreverde mesófilo submontano. Reyes & Acosta (2010) describieron de la parte superior de los mogotes del Carso de Baire las fitocenosis del bosque siempreverde micrófilo calcífilo mientras que refirieron de la parte inferior de los mogotes en estudio el bosque siempreverde mesófilo (Reyes & Acosta 2009). Por ello, el objetivo de este trabajo es exponer las fitocenosis presentes en las áreas por encima de dicho bosque.
Descripción de la zona de estudio
Los mogotes de la Gran Meseta de Guantánamo (Núñez & Viña Bayés 1989), constituyen un área irregular y se extienden entre Yambeque y La Tagua principalmente (alrededor de 27° 17' Norte, 75° 20' Oeste y 20° 22' Norte 75° 10' Oeste), en la parte Centro - Oeste de dicha meseta. Forman un área importante de la provincia de Guantánamo (extremo oriental de Cuba), situada al Norte de la ciudad del mismo nombre. Están constituidos por cúpulas cársicas (Núñez et al. 1989), compuestas por calizas eocénicas de la Formación Charco Redondo. A su vez, sobreyace la Formación Santo Domingo, la cual se observa cuando la erosión destruye la cubierta caliza. Los mogotes poseen una pendiente de alrededor de 45 grados conforma valles en forma de "v" y la superficie está cubierta por "lapiez" o "diente de perro", profundos, que forman oquedades y áreas filosas que dificultan caminar. La altitud varía entre 300 y 600 msm. En el área la precipitación varía entre 1 200 y 1 400 mm (Gagua et al. 1989) y la temperatura oscila entre 22 y 24°C (Lapinel 1989). La zona de los mogotes ha sido antropizada en los bordes y en los valles, principalmente para el cultivo del café. Debido a lo difícil de su orografía los mogotes propiamente dichos, están conservados, manteniendo su vegetación característica; solo se ha extraído madera para uso local.
MATERIALES Y MÉTODOS
Como muestras de vegetación (listas, inventarios, levantamientos), se usaron parcelas de 400 m2 con características típicas, recorriendo además los alrededores para observar lo que pudiera quedar fuera de dicha parcela, éstas fueron realizadas siguiendo los principios sigmatistas. Se diferenciaron en la misma: estructura, composición florística, abundancia - dominancia y sociabilidad de las especies. En la estructura se observó la estratificación y sus características. En la composición florística se determinaron los taxones presentes en la muestra y sus alrededores. La abundancia - dominancia de las especies se estimó en cada estrato de la forma siguiente: 5 cuando cubre 75% o más del área de la muestra; 4 cubre 50 a 75%; 3 cubre 25 a 50%; 2 cubre 5 a 25%; 1 cubre menos del 5%; "+" con pocos ejemplares aislados y con poca cobertura y "r" cuando eran dos o tres ejemplares aislados con poca importancia cenológica. La sociabilidad de las especies se separó de la abundancia – dominancia por un punto. Además, se realizaron observaciones del ecótopo en el lugar de la muestra y sus alrededores: geología, forma del "lapiez", inclinación, exposición y altitud. La ordenación de las listas de vegetación y la separación de las fitocenosis se efectuó por métodos fitocenológicos (Scamoni & Passarge 1959, 1963). Luego de organizada la tabla (y unificadas las fitocenosis), se realizó la comprobación de la homogeneidad de acuerdo con la ley de Raunkiaer (1934). Para la combinación característica de la asociación se utilizaron las especies con grados de presencia IV y V, y para las variantes las combinaciones diferenciales (Scamoni 1960, Samek 1973, Mueller-Dombois & Ellemberg 1974). Para la categorización y la nomenclatura de las fitocenosis se siguió el Principio II y el Artículo 11 del Código de Nomenclatura Fitosociológica (Barkman et al. 1988). La denominación de dichas fitocenosis se realizó según el código expuesto. Como esta zona presenta condiciones particulares de evolución florística, en la selección de las especies características territoriales y locales se utilizó la metodología propuesta por Mueller-Dombois & Ellemberg (1974).
RESULTADOS
Se considera el siguiente arreglo fitosociológico:
- Clase: Thrinaceto – Plumerietea obtusae Borhidi 1996.
- Orden: Coccothrinaci leonis - Guapiretalia rufescentis Reyes ord. nov. En esta contribución.
Typus: Coccothrinaci leonis - Guapirion rufescentis.Composición - especies características: Guapira rufescens, Spathelia pinetorum, Tabebuia dubia, Podocarpus ekmanii, Acrosynanthus trachyphyllus, Bactris cubensis, Ossaea verrucosa (características territoriales), Coccothrinax leonis, Talauma minor, Gesneria cubensis, Pilea fruticulosa (características locales), Coccoloba wrighti, Spirotecoma apiculata, Garcinia aristalata, Amyris lineata, Oplonia tetrasticha, Plumeria tuberculata, Phyllanthus epiphyllanthus, Peperomia alata y Pilea obtusangula.
Fisionomía. Bosque siempreverde micrófilo, calcífilo, bajo, con un estrato arbóreo entre 7 y 12 m (algunos individuos de las especies dominantes hasta 20 m).Distribución. Se desarrolla en los mogotes de la Gran Meseta de Guantánamo, entre Yambeque y La Tagua.
Ecología. Esta fitocenosis ocupa la parte superior de dichos mogotes, sobre los 50 m del fondo de los valles; se desarrolla sin suelo, en el "lapiez".
- Alianza: Coccothrinaci leonis - Guapirion rufescentis Reyes all. nov. En esta contribución
Typus: Bactrio cubensis - Podocarpodetum ekmanii.
Asociaciones estudiadas:
Bactrio cubensis - Podocarpodetum ekmanii.
Phyllantho epiphyllanthi - Tabebuietum myrtifoliae.
Pileo fruticulosae – Thelypteridetum alatae.
- Bactrio cubensis - Podocarpodetum ekmanii Reyes & Acosta ass. nov. En esta contribución
Typus = 1 Tabla 1
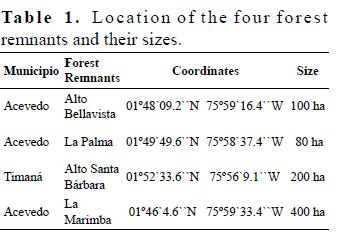
Fisionomía – composición florística. Bosque siempreverde micrófilo, calcífilo, bajo, se presenta en la parte superior de los mogotes, desde cerca de los 50 m sobre el fondo de los valles hasta las cercanías a la cima, ya que entre dicho fondo y la altura expuesta se encuentra un bosque siempreverde mesófilo, constituido por la asociación Ocoteo - Phoebietum elongatae Reyes & Acosta 2009 perteneciente a la clase Swietenio - Calophylletea calabae Borhidi 1996.
El estrato arbóreo cubre entre 70 y 80 % y su altura varía entre 8 y 15 m; cuando tiene dos estratos el superior es de 15 a 16 m, raramente 20 m, y el inferior de 8 a 12 m, ocasionalmente 15 m; promedia 23.8 especies entre las listas. La predominante es Guapira rufescens (Griseb.) Lundell, aunque en menor proporción son localmente abundantes Ocotea coriacea (Sw.) Britt., Buchenavia (tetraphylla) capitata (Vahl) Eichl., Tabebuia dubia (Wr. & Sauv.) Britt. ex Seibert. y Podocarpus ekmanii Urb. El estrato arbustivo tiene entre 40 y 80 % de cobertura, raramente menos y promedia 20.2 especies; es relativamente abundante Spathelia pinetorum M. Vict. y localmente lo son también Ocotea coriacea, Gesneria cubensis (Dcne.) Kuntze, Guapira rufescens, Podocarpus ekmanii y Tabebuia dubia. El estrato herbáceo es el más rico con 33 especies como promedio (Tabla 1), cubre de 60 a 80 % y las especies constantes y relativamenta abundantes son Spathelia pinetorum, Gesneria cubensis, Ocotea coriacea y localmente Guapira rufescens, Oplonia tetrasticha (Wr. ex Griseb.) Stearn., Podocarpus ekmanii, Plumeria tuberculata Lodd. y Guzmania monostachya (L.) Rusby ex Mez. Las lianas presentan diez especies como promedio. Respecto a las epífitas, con ocho especies como promedio, la más abundante y constante es Guzmania monostachya, localmente son también relativamente abundantes Guzmania lingulata (L.) Mez, Hohenbergia penduliflora Mez y Tillandsia fasciculata Sw.
Ecología. Esta fitocenosis tiene una característica peculiar, y es que se desarrolla sin suelo, en el "lapiez", por lo que las plantas solo pueden alcanzar tamaños bajos. El mantillo se compone de una capa L de 2 a 3 cm y una F mayormente de 0.3 a 1 cm, frecuentemente con raicillas. Le sigue una estera radical embebida en una matriz de humus, húmedo, generalmente entre 5 y 7 cm, ocasionalmente más o menos; además hay mucha acumulación de humus en las oquedades del "diente de perro".
Esta fitocenosis es rica, con 59 especies como promedio, la combinación característica se compone de 41 especies. El levantamiento tipo es el 42 (Tabla 1).
- Phyllantho epiphyllanthi - Tabebuietum myrtifoliae Reyes & Acosta ass. nov. En esta contribución (Tabla 2)
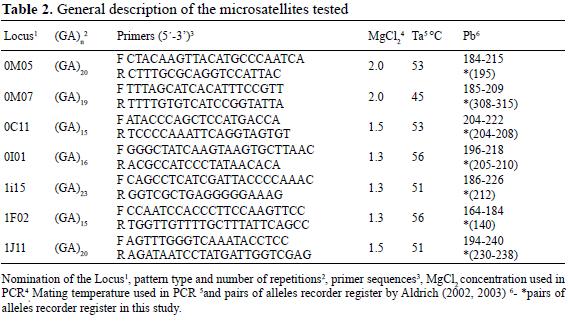
Fisionomía – composición florística. Es un bosque micrófilo calcífilo, claro, bajo. El estrato arbóreo varía entre 6 y 10 m, ocasionalmente 12 m, cubre entre 20 y 50 % y tiene como promedio 13 especies; la predominante es Guapira rufescens, a veces, aunque en menor proporción Coccothrinax leonis Muñiz & Borhidi. El estrato arbustivo es denso, entre 60 y 80 %, excepcionalmente menos; tiene como promedio 15.1 especies y las que más abundan son Spathelia pinetorum, Oplonia tetrasticha y localmente Amyris elemifera L., Coccothrinax leonis y Plumeria tuberculata. El estrato herbáceo, que promedia 30.6 especies, tiene entre 70 y 80 % de cobertura en la variante Neobracea valenzuelana y de 30 a 50 % en la Típica; las especies con mayor cobertura son Phyllanthus epiphyllanthus L., Agave underwoodii Trelease y en ocasiones Oplonia tetrasticha, Guapira rufescens, Spathelia pinetorum, Gesneria cubensis, Guzmania lingulata y Tillandsia fasciculata. Las lianas son relativamente abundantes con 10.8 especies como promedio. En las epífitas, con diez especies como promedio, la más abundante es Tillandsia fasciculata, localmente también Guzmania monostachya.
Ecología. Se desarrolla en la cima de los mogotes, muy batidos por el viento, por lo que las condiciones son más extremas; la vegetación crece sobre el "lapiez", agudo; las epífitas prosperan principalmente sobre la hojarasca y algunas en la parte baja de las plantas. Las capas del mantillo están bien definidas, la L tiene de 1 a 2 cm y la F de 0.5 a 2 cm; la H se constituye en una estera radical bien desarrollada, de más de 10 cm.
Esta fitocenosis es homogenea según la ley de Raunkiaer (1934), tiene 54.8 especies como promedio y 33 en la combinación característica, el levantamiento tipo es el 36.
Se observan dos variantes: Típica y Neobracea valenzuelana.
Variante Típica
Tiene tendencia hacia el Sur; además, se presenta en el extremo Oeste del conjunto de mogotes. Aquí se encuentra la menor cantidad de especies en todos los estratos. Se caracteriza sobre todo negativamente, pues faltan muchas especies, incluso Clusia tetrastigma Vesque, Eupatorium sp. y Tillandsia balbisiana Sch. que forman parte de la combinación característica (Tabla 2).
Variante Neobracea valenzuelana
En esta variante se observa la mayor cantidad de especies en todos los estatos y sinusias. Tiene dos subvariantes Ossaea verrucosa y Guettarda ferruginea, la primera ocupa las exposiciones hacia el Norte, mientras que en la segunda la dirección principal es Este-Oeste. Las combinaciones diferenciales se encuentran en la Tabla 2.
- Pileo fruticulosae - Thelypteridetum alatae Reyes & Acosta ass. nov. En esta contribución
Fisionomía – composición florística. La vegetación tiene una fisonomía peculiar, ya que constituye un estrato herbáceo de 40 a 70 % de cobertura, predominando entre 50 y 60 %; la altura es alrededor de 30 cm, con arbustos aislados de hasta un metro. Hay abundancia de helechos y suculentas (especies de begonias y pileas). Las especies constantes y más abundantes son: Thelypteris alata (L.) Reed f. tipica, T. scolopendrioides (L.) Proctor y Pilea fruticulosa Morton. Son también constantes (a veces abundantes) Marcgravia rectiflora Tr. & Pl. y Gesneria purpurascens Urb. La combinación característica y las diferenciales se observan en la Tabla 3. El levantamiento tipo es el 24.
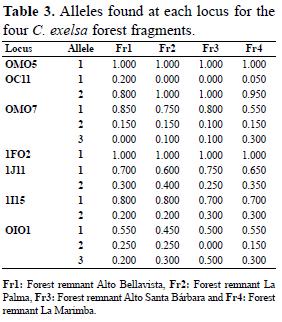
Ecología. Esta fitocenosis se presenta en los paredones verticales (90 grados), que se observan en la parte baja de estos mogotes. Debido a la inclinación, nunca hay acumulación de sustrato, sino las plantas se fijan en las fisuras de las rocas; frecuentemente se advierte una capa delgada, de menos de un centímetro, de restos de musgos, polvo y partículas producidas por la meteorización de la roca, donde se adhieren las raicillas de las pequeñas plantas. Las exposiciones son variadas, predominando las dirigidas al Norte. Por ello el edátopo es muy extremo, pero con humedad ambiental elevada.
DISCUSIÓN
Sintaxonomía
Este bosque siempreverde micrófilo se incluye en la clase Thrinaceto – Plumerietea obtusae Borhidi 1996 por el tipo de formación vegetal y las condiciones geológicas y geomorfológicas donde se desarrolla (Borhidi 1996), ya que difiere considerablemente de los otros sitios donde se describe dicha clase, tanto en las condiciones ecológicas y la fenología como en la composición florística. El orden Coccothrinaci leonis - Guapiretalia rufescentis, aunque cercano a Synapsidi ilicifoliae – Coccothrinacetalia Borhidi 1996 por la geomorfología y la riqueza en pileas y gesnerias, tiene desde el punto de vista de evolución florística condiciones particulares, ya que se desarrolló aislado, en una zona donde predominan las ofiolitas y el complejo basal de Cuba, el cual es sobreyacido por las calizas que conforman estos mogotes, por lo cual comparte muchas especies que son abundantes en los pinares y pluvisilvas nororientales.
Ecología
Al ocupar estas fitocenosis la parte más alta de los mogotes, el edátopo es extremo (ausencia de suelo, afectación por el viento, gran insolación, temperaturas elevadas, alta evaporación y sequedad), por lo que esta vegetación solo puede desarrollarse como un bosque bajo. Se considera, que la diferencia entre este bosque y el que se encuentra debajo, que pertenece incluso a otra clase (Reyes & Acosta 2009), fue explicado por Hager & Zanoni (1993) como debido a la acumulación de niebla nocturna, y que la sombra que proyectan los mogotes disminuye la temperatura y la evaporación, aumentando la humedad en el fondo de los valles, fenómeno conocido como altura relativa. En este caso, se concuerda con Herrera & Rodríguez (1988) y Pérez (2007) que exponen que 50 m (de dicha altura relativa) es el efecto de la linea de drenaje sobre el humedecimiento (cañada, fondo de los valles). Las asociaciones Bactrio cubensis - Podocarpodetum ekmanii y Phyllantho epiphyllanthi - Tabebuietum myrtifoliae constituyen un complejo de comunidades, que presentan numerosas especies en común, que se convierten en características de la alianza y el orden. No obstante, las diferencias en las condiciones ecológicas producen una separación en asociaciones, con sus correspondientes disimilitudes florísticas (Tabla 4). Como se desarrollan totalmente en el "lapiez" (sin suelo), las condiciones de humedad ambiental existentes permiten el desarrollo de una estera radical, constituida por raíces y raicillas vitales (Herrera & Rodríguez 1988, Fornaris 2007, Matos 2007). Los nutrientes se absorben del humus y parcialmente de las hojas en descomposición (Reyes 2005); constituyendo dicha estera un subsistema tropical donde se realiza la recirculación de nutrientes del ecosistema, lo que ya fue observado por otros autores (Reyes & Acosta 2005, Reyes & Fornaris 2011). Al faltar el suelo y el carso ser un sustrato muy percolante, el agua se acumula en el mantillo, que lo cede posteriormente a las raicillas y a los demás usuarios. Debido a que la vegetación de la asociación Pileo fruticulosae - Thelypteridetum alatae se desarrolla en los paredones húmedos de estos mogotes, aunque estructuralmente es diferente, debido a su composición florística se incluye en este orden y alianza, hasta que estudios posteriores aclaren su situación.
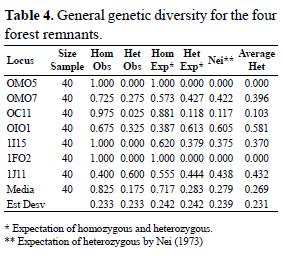
Riqueza de especies
Ambas asociaciones boscosas son ricas en especies. Con mayor riqueza se observa a Phyllantho epiphyllanthi - Tabebuietum myrtifoliae, con con un total de 131 especies y 54.8 de promedio entre las listas, tiene además 33 en la combinación característica, 23 diferenciales y 51 especies que no se repiten entre las listas. Le sigue a su vez, Bactrio cubensis - Podocarpodetum ekmanii con un total de 106 especies y 59 de promedio entre las listas; además, aquí se observa mayor riqueza en los estratos arbóreo, arbustivo y herbáceo que la anterior fitocenosis; presenta también 41 especies en la combinación característica y 43 que se encuentran solo una vez. Pileo fruticulosae –Thelypteridetum alatae, debido a sus condiciones fisiográficas particulares tiene en total 47 especies, con un promedio estre las listas de 13.7, tiene a su vez siete en la combinación característica y nueve diferenciales.
Relación con otros bosques del Caribe
Bosques parecidos, conocidos como Vegetación de los Mogotes, se observan en los mogotes de Los Haitises, República Dominicana, aunque con una composición diferente (Hager & Zanoni 1993).
AGRADECIMIENTOS
Al financista del proyecto nacional "Diversidad biológica de los macizos montañosos Sierra Maestra y Nipe Sagua Baracoa", en cuyo marco se realizó este estudio. Al Dr. Ángel Motito Marín por la revisión del manuscrito y al Ing. Pedro Bergues Garrido por la traducción del resumen al inglés.
LITERATURA CITADA
1. Barkman, J.J., J. Moravec & S. Rauschert. 1988. Código de Nomenclatura fitosociológica. Opusc. Bot. Pharm. Complutensis 4: 9-61. [ Links ]
2. Borhidi, A. 1996. Geography and Vegetation Ecology of Cuba. 2a Ed. Akademiai Kiadó, Budapest. 923 pp. [ Links ]
3. Braun-Blanquet, J. 1979. Fitosociología. Bases para el estudio de las comunidades vegetales. H. Blume ediciones, 820 pp. Madrid. [ Links ]
4. Capote, R. & R. Berazaín. 1984. Clasificación de las formaciones vegetales de Cuba. Revista Jard. Bot. Nac. Univ. Habana 5(2): 27-75. [ Links ]
5. Del Risco, E. 1995. Los Bosques de Cuba: Su historia y características. Ed. Científico Técnica. La Habana. 94 pp. [ Links ]
6. Fornaris, E. 2007. Estudio funcional de algunas formaciones boscosas de la Sierra Maestra. Tesis en Opción al Título Académico de Master en Ecología y Sistemática Aplicada, Mención Ecología. IES-BIOECO. 80 pp. Santiago de Cuba. [ Links ]
7. Gagua, G., S. Zarembo & A. Izquierdo. 1989. Precipitación media anual. 1931-72. En: Nuevo Atlas Nacional de Cuba. Mapa 24. pp. VI.3.1. [ Links ]
8. Lapinel, B. 1989. Temperatura media anual del aire en julio. En: Nuevo Atlas Nacional de Cuba. Mapa 17, pp. VI.2.4. [ Links ]
9. Hager, J. & T.A. Zanoni. 1993. La vegetación natural de la República Dominicana: una nueva clasificación. Moscosoa 7: 39-81. [ Links ]
10. Herrera, R. & M. Rodríguez. 1988. Clasificación funcional de los bosques tropicales. pp. 574-626. En: R.A. Herrera, L. Menéndez, M.A. Rodríguez, E.E García (eds). Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. ROSTLAC. Montevideo. [ Links ]
11. León (Hno.). 1946. Flora de Cuba. Vol. 1. Gimnospermas. Monocotiledóneas. Contribución del Museo de Hist. Nat. del Colegio de la Salle. No. 8. Cultural SA. La Habana. 441 pp. [ Links ]
12. Matos, R. 2007. Funcionamiento Ecológico de la Pluvisilva submontana sobre suelos de mal drenaje de Monte Iberia. Tesis en opción al título de Ingeniero Forestal. Ministerio de Educación Superior. Centro Universitario de Guantánamo. Facultad Agroforestal de Montaña. [ Links ]
13. Mueller - Dombois, D. & H. Ellemberg. 1974. Aims and Methods of Vegetation Ecology. John Wiley and Sons. Londres. 547 pp. [ Links ]
14. Núñez Jiménez, A. & N. Viña Bayés. 1989. Regiones Naturales Antrópicas. En: Nuevo Atlas Nacional de Cuba. Inst. Geografía e ICGC. Mapa 6. pp. XII.2.1. [ Links ]
15. Nuñez Jiménez, A., N. Viña Bayés & A. Graña. 1989. Carsología. En: Nuevo Atlas Nacional de Cuba. Mapa 1. pp. V.1.2-3. [ Links ]
16. Pérez, E. 2007. Reforestación sucesional en la Sierra de Cubitas, Camaguey, Cuba. Tesis en opción a Dr. Ciencias Biológicas. Instituto Ecología y Sistemática. CITMA. La Habana. 101 pp. + Bibliografía y Anexos. [ Links ]
17. Raunkiaer, C. 1934. The life forms of plants an statistical plant geography. Oxford Univ. Press. 632 pp. [ Links ]
18. Reyes, O.J. 2005. Estudio sinecológico de las Pluvisilvas submontanas sobre rocas del complejo metamórfico. Foresta Veracruzana 7(2): 15-22. [ Links ]
19. Reyes, O.J. 2006. Clasificación de la vegetación de la Sierra Maestra. Biodiversidad de Cuba Oriental 8: 23–41. [ Links ]
20. Reyes, O.J. & F. Acosta Cantillo. 2005. Vegetation. pp. 113–119. D. Maceira, A. Fong, W. S. Alverson & T. Wachter, Eds. Cuba: National Park La Bayamesa. En: Rapid Biological Inventories: 13. The Field Museum, Chicago. [ Links ]
21. Reyes, O.J. & F. Acosta Cantillo. 2009. Fitocenosis en los bosques siempreverdes de Cuba Oriental. I. Ocoteo-Phoebietum elongatae en los mogotes de la Gran Meseta de Guantánamo. Foresta Veracruzana 11(1): 1-8. [ Links ]
22. Reyes, O.J. & F. Acosta Cantillo. 2010. Nuevas fitocenosis del Carso de Baire. Rodriguésia 61 (3): 519-530. [ Links ]
23. Reyes, O.J. & E. Fornaris. 2011. Características funcionales de los principales bosques de Cuba Oriental. Polibotánica 32: 83-99. [ Links ]
24. Samek, V. 1973. Pinares de la Sierra de Nipe; Estudio Sinecológico. Academia de Ciencias de Cuba, Serie Forestal 14. La Habana. 58 pp. [ Links ]
25. Scamoni, A. 1960. Waldgesellschaften und Waldstandorte. Akademie – Verlag, Berlín. 326 pp. [ Links ]
26. Scamoni, A. & H. Passarge. 1959. Gedanken zu einer natürlichen Ordnung der Waldgesellschaften. Arch. Forstw. 8: 382-426. [ Links ]
27. Scamoni, A. & H. Passarge. 1963. Einführung in die praktische Vegetationskunde. 2 Aufl. Jena. 236 pp. [ Links ]
