










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.35 no.2 Bogotá July/Dec. 2013
FAVIO GONZÁLEZ
NATALIA PABÓN-MORA
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, D.C., Colombia. fagonzalezg@unal.edu.co
Instituto de Biología, Universidad de Antioquia, Apartado 1226, Medellín, Colombia.
ABSTRACT
Castilleja paramensis F. González & Pabón-Mora, a perennial species of small herbs from páramos of Santander, Boyacá and Cundinamarca, in the Colombian Eastern Cordillera, is described and fully illustrated by photographs of the type collection. The new species is morphologically and ecologically similar to C. nubigena, C. pumila, and C. virgata, three perennial high Andean species from Ecuador, Peru and northern Chile and Argentina. C. paramensis is clearly distinguished from its relatives by the small size of the individuals, the broadly elliptic leaves and bracts (which are entire to slightly trilobed), the short (< 1 mm) pedicel, the small size (0.8- 1.15 cm long) of the flowers, the not inflated calyx with the distal half bright yellow, the corolla often included to slightly exserted from the calyx, and the included style and stigma. The included corolla, stamens, styles and stigmas in C. paramensis strongly suggest the occurrence of cleistogamy. The new species often grows as a root hemiparasite of Plantago rigida.
Key words. High Andean flora, flora of Colombia, hemiparasitic plants, horizontal gene transfer, páramo endemics, root hemiparasites.
RESUMEN
Castilleja paramensis F. González & Pabón-Mora, una especie de hierbas perennes pequeñas propias de páramos de Santander, Boyacá y Cundinamarca, en la Cordillera Oriental Colombiana, es descrita y detalladamente ilustrada con fotografías de la colección tipo. La especie nueva es morfológica- y ecológicamente similar a C. nubigena, C. pumila y C. virgata, tres especies perennes de zonas altoandinas de Ecuador, Perú y norte de Chile y Argentina. Castilleja paramensis se distingue claramente de sus especies afines por el tamaño pequeño de los individuos, las hojas y las brácteas ancho-elípticas (las cuales son enteras a levemente 3-lobadas), el pedicelo extremadamente corto (< 1 mm), el tamaño reducido de las flores (0.8- 1.15 cm largo), el cáliz no inflado, con la mitad distal de color amarillo brillante, la corola generalmente inserta o muy poco exerta del cáliz, y el estilo y estigma insertos. La corola, los estambres y los estigmas insertos en C. paramensis sugieren la ocurrencia de cleistogamia. La nueva especie se encuentra frecuentemente como hemiparásita de raíces de Plantago rigida.
Palabras clave. Endemismos de páramo, flora altoandina, flora de Colombia, hemiparásitos de raíces, plantas hemiparasíticas, transferencia horizontal de genes.
Recibido: 19/02/2013
Aceptado: 05/09/2013
INTRODUCTION
Castilleja Mutis ex L. f. comprises approximately 200 species (Holmgren 1978, Chuang & Heckard 1991, Heide-Jørgensen 2008) most of which are native to western North America. The South American species (estimated between 15 and 20; Egger, pers. comm.) are concentrated in the Andes. During the ongoing study of parasitic and hemiparasitic plants in Colombia by the authors, a number of specimens of Castilleja collected in the páramos of the Eastern Cordillera drew our attention because of the small size of the individuals and the frequent association with Plantago rigida Kunth. A specimen recently collected by FG in the páramo of Siscunsí (Sogamoso, Boyacá) matches all the morphological and ecological traits of the small-sized individuals previously collected in similar páramos of Santander, northern Boyacá, and Cundinamarca, in the Colombian Eastern Cordillera. Until now, these specimens have remained either undeterminated or were tentatively identified as infraspecific taxa of C. fissifolia L. f. or assigned to C. nubigena Kunth, a species known from the high Andes of Ecuador and Peru (cf. Holmgren 1984, Luteyn 1999, Molau, 1999). A detailed examination of these collections reveals, however, that they do not match any of the previously described species of Castilleja from Costa Rica/Panama (cf. Holmgren 1978) Venezuela (cf. Pennell 1953, Egger 2009), Colombia (cf. Fernández-Alonso 1987), Ecuador (cf. Holmgren 1984) or Peru (cf. Weddell 1857, Herrera 1926, Edwin, 1971). Thus, here we describe the new species and compare it with other high Andean species of Castilleja characterized by the small size of the individuals. Besides the disjunct distribution between the new species and the other morphologically similar high Andean species (C. nubigena, C. pumila (Benth.) Wedd. ex Herrera, and C. virgata (Wedd.) Edwin), the newly described species possesses clear-cut differences in the size and overall morphology of leaves, bracts, and flowers (Table 1).
Additionally, we include preliminary observations on the association of the species with Plantago rigida, as root hemiparasitism and hence physical plant interactions may also have genetic implications, for instance the occurrence of Horizontal Gene Transfer (HGT).
HGT has been reported between members of the Orobanchaceae and Plantaginaceae, preferentially involving mitochondrial and ribosomal over plastid genes (Mower et al. 2004, Davis & Wurdack 2004). For instance, the mitochondrial atp1 has been transferred from Bartsia (Orobanchaceae) to Plantago (Mower et al. 2004). Plastid HGT seemed to have occurred less frequently, like between the parasitic Orobanche and Pelipanche (Park et al. 2007). These are both non-photosynthetic parasites with a shared plastid region including rps2, trnL-F, and rbcL that has probably been transferred via their common host (Park et al. 2007). At this point the genetic consequences of the Castilleja paramensis-Plantago rigida association are still unknown; however, their tight physical link and their exclusive distributions in high altitudes in the Andes, make them an excellent candidate to undertake studies testing explicitly for the occurrence of HGT.
Castilleja paramensis F. González & Pabón-Mora, sp. nov. TYPE: COLOMBIA. Boyacá: Sogamoso, Páramo de Siscunsí, alrededores de la Laguna de Siscunsí, 3875 m, 5º22'N, 72º50'W, 04 Feb 2013 (fl), F. González, G. Plunkett, A. Nicolas, J. Aguirre & A. Herrera 4478 (Holotype: COL; isotypes: HUA, NY, UPTC, to be distributed). Fig. 1, Fig. 2, Fig. 3


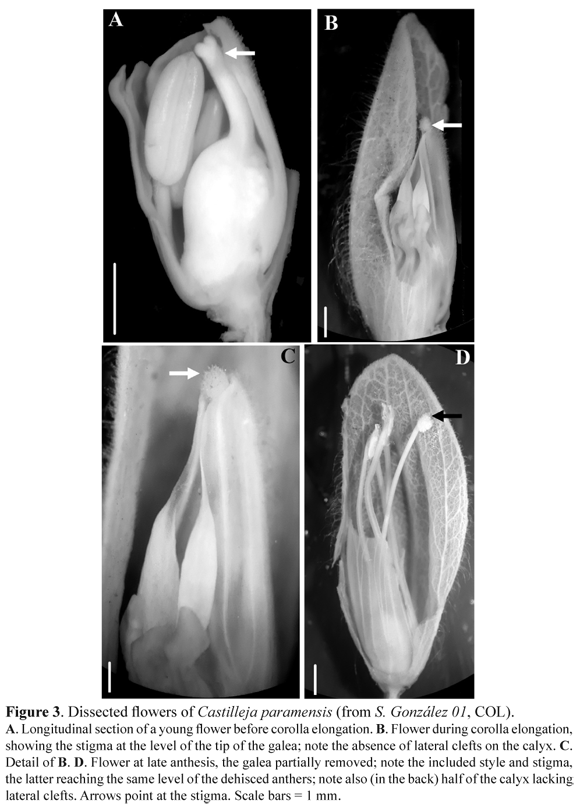
Small, rhizomatous, decumbent, perennial herbs; ascending stems to 5(-8) cm tall, branching from the base. Rhizomes very thin, whitish, with a few scattered achlorophyllous triangular scale-leaves to 2 mm long and 1 mm wide. Internodes of the aerial shoots very short, to 3 mm long. Leaves congested, sub-opposite, cucullate, sessile, broadly elliptic, to 7 mm long and 5 mm wide, entire to slightly bi- or trilobate, light green when young, later with the distal half purplish, lateral lobe(s) triangular, ascending, glabrous above, scarcely villous below, margins villous, especially along the distal half. Bracts broadly elliptic, bi- or trilobate and bright red in their distal half, glabrous above, villous below, with those subtending the mature flowers becoming purplish. Flowers subsessile, pedicel < 1 mm long, axillary to the 3-4 most distal bracts. Calices 8- 11 mm long, green when immature, with bright yellow distal half at maturity, not inflated, villous, tube to 6 mm long, median clefts strongly unequal, the adaxial 1- 1.5 mm deep, the abaxial 4.5- 5.5 mm deep, lateral clefts absent. Corollas 8- 11.5 mm long, completely included or very slightly exserted from the calices, hyaline, straight to very slightly incurved, tubes 5- 6 mm long, the galeas 4- 5.5 mm long, villous along the adaxial mid-line, the lower three lips consisting of bright green, extremely reduced, triangular, to 0.5 mm long and 0.2 mm wide, ascending, glabrous. Stamens included, slightly didynamous; filaments 8.5- 10.5 mm long, adnate to the corolla tube for its proximal 1/3, anthers lanceolate, ca. 0.5- 0.8 mm long, thecae slightly asymmetric. Styles as long as the corollas and included during anthesis. Stigmas slightly bilobed, poorly differentiated from the styles, papillose. Capsules oblong, to 10 x 5 mm, villous. Seeds obconical, 1.3-1.5 x 0.8- 1 mm, seed coat papery, translucent, unruptured, with deep and narrowly hexagonal reticulation cells.
Etymology: The specific name refers to the ecosystem where the species has been found.
Distribution and phenology: Castilleja paramensis has so far been collected in páramos of the departments of Santander, Boyacá and Cundinamarca ( Colombia ). It sets flowers in September, November to March and in May, and fruits in February, June and September.
Vernacular name: " Flores de lagarto" (Güicán, Boyacá; FG 1391).
Additional specimens examined (when available, we have included the field notes referring to the association of the new species with Plantago rigida). COLOMBIA. Boyacá: Morrena seca 200 m NW de la Laguna Pintada, con Espeletia colombiana predominante, asociada con Aciachne pulvinata, Agrostis breviculmis, Agrostis trichoides y Acaena cylindrostachya, 3995 m, 25 Sep 1972 (fl, fr), A. M. Cleef & P. A. Florschütz 5508 (COL); Páramo de La Rusia, NW-N de Duitama, Peña Blanca, 2 km al NE de Buenos Aires, pantanos con almohadas de Plantago rigida predominantes, 3720 m, 16 Dic 1972 (fl), A. M. Cleef 7317 (COL); Sierra Nevada del Cocuy, páramo Cóncavo, superpáramo en el límite con el páramo p.d., 3.5 km al NNW del morro Púlpito del Diablo, cabeceras de vallecitos entre morrenas, protegidas a viento con un frailejonal denso y húmedo de Espeletia lopezii, 4335 m, 26 Feb 1973 (fl, fr), A. M. Cleef 8530 (COL); Socotá, páramo de Pisba, sitio La Playita, entre colchones de Plantago rigida (FG 824), c 3600 m, 15 Nov 1986 (fl), F. González et al. 823 (UPTC); Socha, sitio Los Pinos, 3600 m, asociada a Plantago rigida (FG 1118A), 25 Jun 1987 (fl), F. González et al. 1118 (UPTC); Güicán, ascenso al Ritacuba Blanco, por las cabañas del Himat, 3800- 4800 m, 1 Mar 1988 (fl), F. González et al. 1391 (UPTC); Güicán, Sierra Nevada del Cocuy, carreteable entre la cabaña del Inderena y las Lagunillas, frente a la Laguna La Atravesada, 4000 m, entremezclada con Plantago rigida (FG 1417), 2 Mar 1988 (fl), F. González et al. 1416 (UPTC); Belén, páramo del Güina, 3500 m, 22 May 1993 (fl), F. González & R. Small 2720 (COL, UPTC); Sierra Nevada del Cocuy, sector SE, valle del río Lagunillas, entre 3650 y 4000 m, 24 May 1993 (fl), F. González & R. Small 2734 (COL, UPTC); Güican, ascenso hacia el Boquerón de Cardenillo, 6º18'50.6''N, 72º12'15.6''W, ca. 4300 m, 17 Ago 2013 (fl), S. González et al. 01 (COL); Güicán, Sierra Nevada del Cocuy, Laguna Grande de los Verdes, en colchones de Distichia sp., 3900- 4100 m, 20 Sep 1978 (fl, fr), O. Rangel & H. Sturm 1534A (COL); below the Playas of Ritacuba in the Cocuy mountains above Güicán, locally frequent in poorly drained damp paramo 3700- 4500 m, particularly in damp flushes and poorly drained grassy hollows, 4200 m, 24 Jun 1984 (fl, fr), J. R. I. Wood 4458 (COL). Cundinamarca: Páramo de Sumapaz, Chisacá, pantano con cojines de Plantago rigida, 3680 m, 11 Dic 1971 (fl), A. M. Cleef 143 (COL); Macizo de Bogotá, páramo de Chisacá, around the lakes at about 3620 m, 16 Sep 1961 (fl), J. Cuatrecasas & R. Jaramillo M. 25972 (COL). Santander: Cordillera Oriental, páramo del Almorzadero, above 3600 m, slope above ciénaga, very wet, with hummocks of mosses, Plantago rigida, etc., 31 Dec 1959-1 Jan 1960 (fl), H. G. Barclay & P. Juajibioy 10369 (COL).
Castilleja paramensis is morphologically similar to C. pumila, C. virgata and especially to C. nubigena (Table 1).
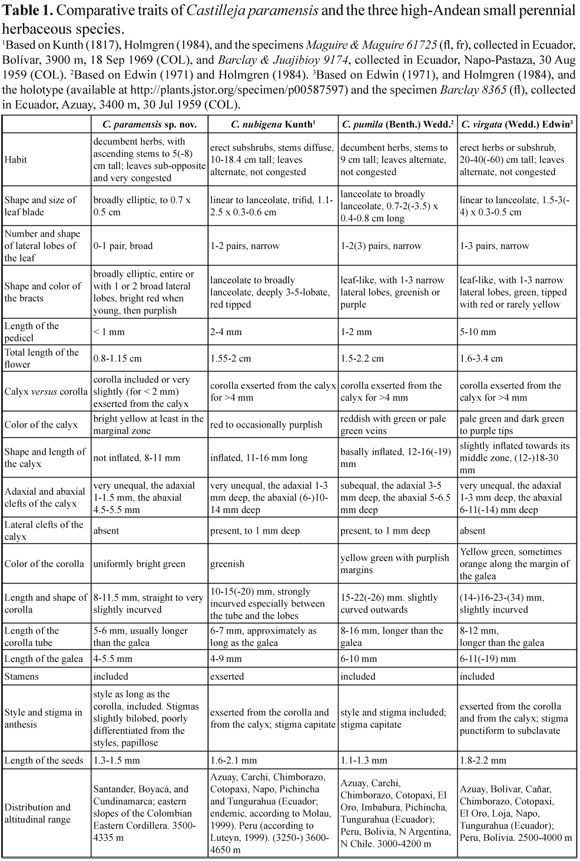
It clearly differs by its smaller size (up to 8 cm), broadly elliptic and entire to slighly trilobed leaves and bracts, reduced pedicel (<1mm) and flower (0.8- 1.15 cm) lenght, bright yellow and not inflated calyx, and corolla included or very slightly (for < 2 mm) exserted from the calyx (Fig. 1). Additional differences between C. paramensis and C. nubigena include the absence of lateral clefts of the calyx (Fig. 3B, D), the straight to very slightly incurved corolla, the included stamens and the bilobed, poorly differentiated stigma in the former, versus the presence of lateral clefts of the calyx, the strongly incurved corolla, the exserted stamens and the capitate stigma in the latter. Finally, the presence of included corolla, stamens, styles and stigmas (Fig. 1, Fig. 3) in C. paramensis strongly suggests the ocurrence of cleistogamy in this species.
On the systematics and life history of Castilleja: The systematics of Castilleja has been considered difficult, mainly due to polyploidy, lack of crossing barriers among species, and effective natural hybridization, all of which can drastically change the morphological traits used to distinguish species (Heckard 1964, Heckard & Chuang 1977, Chuang & Heckard 1991, Mathews & Lavin 1998, Hersch-Green & Cronn 2009, Clay et al. 2012, Hersch-Green 2012). At a suprageneric level, polyploidy has also led to differential sampling of paralogs (e.g. PHYA) in phylogenetic analyses, resulting in a polyphyletic Castilleja (Bennett & Mathews 2006).
The South American species of Castilleja fall into two different subgenera, Castilleja and Colacus. All the Colombian species belong to the subgenus Castilleja (sensu Chuang & Heckard 1991), as the flowers are narrow and tubular, the associated bracts, the calices and/or corollas are red, and the galea is at least three times longer than the lower three lips. All these traits have been associated with hummingbird, pollination in the genus (Pennell 1948, Chuang & Heckard 1991, Heide-Jørgensen 2008). Hummingbird pollination, in turn, appears to be correlated with the evolution of perenniality in the genus (Tank & Olmstead 2008, 2009). This perennial habit has evolved once from an annual condition in Castilleja, and it seems to be tightly associated with polyploidy at least in the North American species (Tank & Olmstead 2008). The new species can be assigned unequivocally to subgenus Castilleja, and it is morphologically and ecologically similar to the high-Andean species C. nubigena, C. pumila and C. virgata (Table 1). The latter three species form a clade of perennial but primarily diploid species (Tank & Olmstead 2009). We anticipate that the perennial C. paramensis belongs to this clade; however, chromosome counts and molecular data are needed to confirm this placement. In addition, studies on pollination biology of C. paramensis are critical, as its colorful flowers are near the ground and appear to be too small and too low for hummingbird pollination. The included style and stigma of C. paramensis (Fig. 3) strongly suggest that the species might be cleistogamous, despite the fact that it can co-occur with C. integrifolia, a species pollinated by the hummingbird Oxypogon guerinii (Salamanca-Reyes 2011).
Castilleja species as facultative root hemiparasites. Species of Castilleja are facultative root hemiparasites, which means they can complete their life cycle either as parasites or singly without a host (Heckard 1962, 1964, Matthies 1997, Heide-Jørgensen 2008, Fay et al. 2010). Castilleja host association is broad and can infect species across a diverse range of flowering plant families (Heckard 1962, Musselman & Press 1995), though some species might have more exclusive associations. This is the case for C. paramensis, frequently found associated belowground with Plantago rigida (Fig. 2). In fact, Cleef (1981:127) included C. paramensis (referred there as "C. fissifolia ssp. pygmaea, ssp. nom. herb.; in ed.?") and P. rigida as two primary components of the Wernerietea syntaxonomy that conforms the "flush vegetation and cushion bogs".
Seed germination in Castilleja can occur in the absence of a host (Heckard 1962); however, species from high elevations may require moist and cold conditions that can slow down germination rates (Kuijt 1969). Heckard (1962) examined hemiparasitic Castilleja species growing with and without hosts and noticed that generally plants growing as parasites are more vigorous, branch more profusely, and exhibit early flowering. Haustoria in Castilleja are small (less than 1 mm in diameter) and are not as invasive or penetrating as haustoria of other parasitic flowering plants. They initiate as elongated epidermal cells similar to root hairs (Kuijt 1969). The first responses during haustorial initiation are cortical enlargement and increasing cell division, resulting in lateral or apical protuberances in the parasite root similar to other Orobanchaceae, such as Bartsia, Lathraea, Melampyrum, Pedicularis and Rhinanthus (Heckard 1962, Dobbins & Kuijt 1973a, Weber & Weberling 1975, Riopel & Timko 1995). A mature haustorium of Castilleja is composed of a plate xylem zone surrounded by parenchyma, a centrally located core of collenchyma (with vessel members scattered), and the haustorial surface (Dobbins & Kuijt 1973a). Penetration is likely due to cell dissolution in the host, accompanied by crushing of cells (Dobbins & Kuijt 1973b). The haustoria of C. paramensis are spherical to cup-shaped and usually terminal in secondary roots.
ACKNOWLEDGEMENTS
Thanks to J. Aguirre, A. Nicolas, and G. Plunkett (The New York Botanical Garden), and A. Herrera and S. González (National University of Colombia), for accurate observations and valuable help during fieldwork while collecting the type specimen and some of the paratypes. Thanks also to J. M. Egger (Herbarium, Burke Museum of Natural History and Culture, University of Washington, Seattle), O. Rangel ( Institute of Natural Sciences, National University of Colombia), and three anonymous reviewers, for valuable comments on the manuscript.
LITERATURE CITED
1. Bennett, J.R. & S. Mathews. 2006. Phylogeny of the parasitic plant family Orobanchaceae inferred from Phytochrome A. American Journal of Botany 93: 1039-1051. [ Links ]
2. Chuang, T.I. & L.R. Heckard. 1991. Generic realignment and synopsis of subtribe Castillejinae (Scrophulariaceae–tribe Pediculareae). Systematic Botany 16: 644-666. [ Links ]
3. Clay, D.L., S.J. Novak, M.D. Serpe, D.C. Tank & J.F. Smith. 2012. Homoploid hybrid speciation in a rare endemic Castilleja from Idaho (Castilleja christii, Orobanchaceae). American Journal of Botany 99: 1976-1990. [ Links ]
4. Cleef, A. M. 1981. The vegetation of the páramos of the Colombian Cordillera Oriental. Dissertationes Botanicæ 61. J. Cramer, Vaduz. [ Links ]
5. Davis, C.C. & K. Wurdack. 2004. Host-to-parasite gene transfer in flowering plants: phylogenetic evidence from Malpighiales. Science 305: 676-678. [ Links ]
6. Dobbins, D. & J. Kuijt. 1973a. Studies on the haustorium of Castilleja (Scrophulariaceae). I. The upper haustorium. Canadian Journal of Botany 51: 917-922. [ Links ]
7. Dobbins, D. & J. Kuijt. 1973b. Studies on the haustorium of Castilleja (Scrophulariaceae). II. The endophyte. Canadian Journal of Botany 51: 923-931. [ Links ]
8. Edwin, G. 1971. Scrophulariaceae. Flora of Peru. Field Museum of Natural History, Bot. Ser. 13, part 5B (3): 461-717. [ Links ]
9. Egger, J.M. 2009. A new species of Castilleja (Orobanchaceae) from Trujillo. Venezuela. Brittonia 61:41-45. [ Links ]
10. Fay, M.F., J.R. Bennett, K.W. Dixon & M.J.M. Christenhusz. 2010. Parasites, their relationships and the disintegration of Scrophulariaceae sensu lato. Curtis's Botanical Magazine 26: 286–313. [ Links ]
11. Fernández-Alonso, J.L. 1987. Castilleja. Flora de la Real Expedición Botánica del Nuevo Reyno de Granada, Vol. 39: 77-82, T. LVIII-LXIV. Ediciones Cultura Hispánica, Madrid. [ Links ]
12. Fernández-Alonso, J.L. 2000. Scrophulariaceae. Pp. 349- 359 in O. Rangel-Ch. (ed.) Colombia Diversidad Biótica III. La región de vida paramuna. Universidad Nacional de Colombia, Bogotá [ Links ].
13. Heckard, L.R. 1962. Root parasitism in Castilleja. Botanical Gazette 124: 21-29. [ Links ]
14. Heckard, L.R. 1964. Causes of taxonomic complexity in Castilleja. American Journal of Botany 51: 686 (abstract). [ Links ]
15. Heckard, L.R. & T.-I. Chuang. 1977. Chromosome numbers, polyploidy, and hybridization in Castilleja (Scrophulariaceae) of the Great Basin and Rocky Mountains. Brittonia 29: 159-172. [ Links ]
16. Heide-Jørgensen, H. 2008. Parasitic Flowering Plants. Brill, Leiden, 438 pp. [ Links ]
17. Herrera, F.L. 1926. Chloris Cuzcoensis. Cuzco, Perú, 222 pp. [ Links ]
18. Hersch-Green, E.I. 2012. Polyploidy in Indian paintbrush (Castilleja; Orobanchaceae) species shapes but does not prevent gene flow across species boundaries. American Journal of Botany 99: 1680-1690. [ Links ]
19. Hersch-Green, E.I. & R. Cronn. 2009. Tangled trios?: characterizing a hybrid zone in Castilleja (Orobanchaceae). American Journal of Botany 96: 1519–1531. [ Links ]
20. Holmgren, N.H. 1978. Castilleja (Scrophulariaceae) of Costa Rica and Panama. Brittonia 30: 182–194. [ Links ]
21. Holmgren, N.H. 1984. Castilleja. Pp. 174–184. In: G. Harling & B. Sparre (eds.), Flora of Ecuador, Vol. 21. Swedish Research Councils, Stockholm, Sweden. [ Links ]
22. Kuijt, J. 1969. The biology of parasitic flowering plants. University of California Press, Berkeley, 246 pp. [ Links ]
23. Kunth, K. S. 1817. Castilleja. Pp. 329-332. S. In F.H. Humboldt (eds.), Nova Genera et Species Plantarum Vol. 2. Paris. [ Links ]
24. Luteyn, J.L. 1999. Páramos. A checklist of plant diversity, geographic distribution and botanical literature. Memoirs of The New York Botanical Garden 84: 1- 278. [ Links ]
25. Mathews, S. & M. Lavin. 1998. A biosystematic study of Castilleja crista-galli (Scrophulariaceae): An allopolyploid origin reexamined. Systematic Botany 23: 213-230. [ Links ]
26. Matthies, D. 1997. Parasite-host interactions in Castilleja and Orthocarpus. Canadian Journal of Botany 75:1252-1260. [ Links ]
27. Molau, U. 1999. Scrophulariaceae. Pp. 891-899 in: P.M. jørgensen & s. león-yanez (eds.), Catalogue of the Vascular Plants of Ecuador. Monographs in Systematic Botany from the Missouri Botanical Garden Press, Vol. 75, St. Louis, Missouri. [ Links ]
28. Mower, J.P., S. Stefanovic, G.J. Young & J.D. Palmer. 2004. Gene transfer from parasitic to host plants. Nature 432 (11 Nov): 165-166. [ Links ]
29. Musselman, L.J. & M.C. Press. 1995. Introduction to parasitic plants. Pp. 1- 13 in M.C. Press & J. D. Graves (eds.), Parasitic plants. Chapman & Hall, London. [ Links ]
30. Park, J.-M., J.-F. Manen & G.M. Schneeweiss. 2007. Horizontal gene transfer of a plastid gene in the non-photosynthetic flowering plants Orobanche and Phelipanche (Orobanchaceae). Molecular Phylogenetics and Evolution 43: 974-985. [ Links ]
31. Pennell, F.W. 1948. The taxonomic significance of an understanding of floral evolution. Brittonia 6:301-308. [ Links ]
32. Pennell, F.W. 1953. Castilleja. In: J. A. Steyermark et al. (eds.), Botanical explorations in Venezuela, III. Fieldiana: Botany 28: 516–519. [ Links ]
33. Riopel, J.L. & M.P. Timko. 1995. Haustorial initiation and differentiation. Pp. 39- 79 in M.C. Press & J.D. Graves (eds.), Parasitic plants. Chapman & Hall, London . [ Links ]
34. Salamanca-Reyes, J.R. 2011. Ecología del barbudito de páramo (Oxypogon guerinii, Trochilidae) en el páramo de Siscunsí, Boyacá, Colombia. Ornitología Colombiana 11: 58-75. [ Links ]
35. Tank, D.C. & R.G. Olmstead. 2008. From annuals to perennials: phylogeny of subtribe Castillejinae (Orobanchaceae). American Journal of Botany 95: 608–625. [ Links ]
36. Tank, D.C. & R.G. Olmstead. 2009. The evolutionary origin of a second radiation of annual Castilleja (Orobanchaceae) species in south America: the role of long distance dispersal and allopolyploidy. American Journal of Botany 96: 1907–1921. [ Links ]
37. Webber, H.C. & F. Weberling. 1975. Zur Morphologie der Haustorien von Rhinanthoideen (Scroph.). Berichte der Deutschen Botanischen Gesellschaft 88: 329-345. [ Links ]
38. Weddell, H.A. 1857. Chloris Andina, Vol. 2. P. Bertrand, Paris, 364 pp. [ Links ]
