










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.35 no.2 Bogotá July/Dec. 2013
MARISOL AMAYA MÁRQUEZ
LAURENCE E. SKOG
LARS PETER KVIST
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C., Colombia. mamayam@unal.edu.co
Department of Botany MRC-166, PO Box 37012, Smithsonian Institution, Washington, DC 20013-7012 U.S.A. SKOGL@si.edu
Institute of Biological Sciences, University of Aarhus, Building 540, Ny Munkegade, DK-8000, Aarhus C., Denmark. larskvist@biology.au.dk
ABSTRACT
Two new species of Columnea belonging to section Collandra (Gesneriaceae) from the "cordillera Occidental" in the Colombian Andes are described and illustrated. Columnea caudata is distributed along the Biogeographical Chocó in the Departments of Antioquia, Chocó, Risaralda, and Valle del Cauca, whereas Columnea megafolia is restricted to Antioquia, and probably is an endemic species of the National Natural Park Las Orquídeas.
Key words. Collandra, Columnea , Colombia, Flora of Colombia, Gesneriaceae, Biogeographical Chocó, Plant Taxonomy.
RESUMEN
Se describen e ilustran dos nuevas especies de Columnea pertenecientes a la sección Collandra (Gesneriaceae) en la Cordillera Occidental en Colombia. Columnea caudata se distribuye en el Chocó Biogeográfico Colombiano, en los departamentos de Antioquia, Chocó, Risaralda, y Valle del Cauca, mientras que Columnea megafolia tiene una distribución restringida a los bosques montanos en Antioquia, siendo probablemente endémica del Parque Nacional Natural Las Orquídeas.
Palabras clave. Collandra, Columnea, Colombia, Flora de Colombia, Gesneriaceae, Chocó biogeográfico, Taxonomía Vegetal.
Recibido: 16/05/2013
Aceptado: 06/08/2013
INTRODUCTION
The taxonomic revision of section Collandra (Lem.) Benth. & Hook.f. (Columnea L., Gesneriaceae) has revealed that our knowledge about this lineage of plants is far from complete. Several new species of Columnea, most of them from Colombia, have been published in the last ten years (e.g., Amaya-Márquez et al. 2004; Kriebel 2005; Amaya-Márquez 2010a, b; Amaya-Márquez & Clark 2011; Amaya-Márquez & Marín-Gómez 2012; Amaya-Márquez & Smith 2012; Clark & Clavijo 2012). The species of the genus Columnea play an important role in maintaining biodiversity due to the specialized system of pollination by hummingbirds (Morley 1971; Jones & Rich 1972; Stiles & Freeman 1993; Amaya-Márquez 1996; Kastinger & Weber 2000). Biotic specialization, (e.g., the Neotropical interaction hummingbird-Columnea) is known to promote coexistence and speciation (Thompson 1994; Dyer et al. 2007; Schemske 2009) and therefore these ecological interactions may determine local and global biodiversity patterns (Schemske 2002, Schemske et al. 2009). Colombia is the country with the highest number of species of Columnea (Kvist & Skog 1993; Skog & Boggan 2007) as well as of hummingbird species (Schuchman 1999), which may be the result of selection driven by interactions between these two groups. These facts call for a great responsibility to preserve the biodiversity and the ecological interactions among species (Schemske 2002) and a precondition for this, is to document the species that exist and investigate the ecology including their coevolution with pollinators, dispersers, and other organisms; one limitation to go farther in building this type of knowledge is the lack of taxonomic tools available to identify species. As part of the effort to overcome taxonomic limitations in the knowledge of the genus Columnea, we report in this paper two new species of Columnea from the Biogeographical Chocó, one of the most biodiverse regions in Colombia and in the world (Myers et al. 2000; Rangel-Ch. & Rivera-Diaz 2004).
Columnea caudata and C. megafolia are assigned to section Collandra on the basis of having dorsiventral shoots typical for the species of the section, and by having a solitary bilobate or tridentate nectar gland. Additionally, these species have a great similarity to C. ericae Mansf. and C. purpurata Hanst. respectively, two well-known species belonging to section Collandra. Columnea caudata is unique among the species of section Collandra by having a long tubular corolla with a bilateral limb, with two lateral lobes and four appendages inside the corolla, as well as having the larger leaf at each node on the stem present a long caudate apex. C. megafolia has large membranous leaves lacking red spots, but having conspicuous inflorescences crowded with several flowers and bracts. The corolla tube is sigmoid and the limb has a marked bilateral symmetry.
Columnea caudata M. Amaya & L. P. Kvist sp. nov. Figure 1
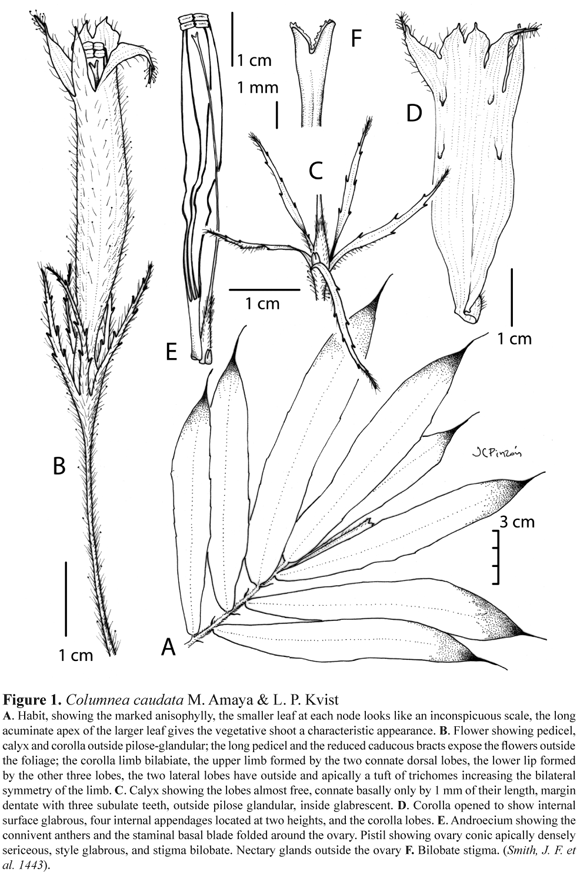
TYPE: COLOMBIA: Valle del Cauca, trail uphill from Río Calima immediately across river from encampment Cuzumbu, 400- 1000 m, 21 Feb. 1989, Smith, J. F. et al. 1443 (Holotype: COL, Isotypes: MO, WIS, US).
Columnea caudata differs from C. ericae by the larger leaf of a pair having a long caudate apex and long narrow corollas with a constricted zygomorphic limb.
Suffrutescent woody climber. Stem subterete, 0.5- 0.7 cm in diam., sericeous; internodes 1- 2 cm long. Leaves opposite, strongly anisophyllous in a pair, papyraceous; larger leaf sessile; blade asymmetric, narrow oblanceolate to falcate, 8.5-23.5 x 1.3- 3.5 cm, base truncate and slightly oblique, apex long caudate, margin serrate; adaxially green and densely sericeous, veins obscure, abaxially green and red-purple apically (ca. 1/5th of the leaf length), sometimes the margins are red-colored, densely sericeous (10- to 14-celled trichomes) and with setulose unicellular trichomes sparsely distributed, 13-17 veins on the larger side of the midrib, veining mixed craspedodromous; smaller leaf sessile, linear, 5-7 x 0.5 mm . Inflorescence fasciculate of 1-2 flowers in the larger leaf axil; 1-2 bracts, lanceolate 2.3 x 0.3 cm or reduced linear 4.0 x 0.5 mm, caduceus. Flowers pedicellate, pedicel 2.5- 3.5 cm, pilose glandular (uniseriate 6- to 10-celled trichomes ending in a glandular head). Calyx green or pale yellow, lobes almost free, connate basally for about 1 mm, subequal, very narrowly elliptic, 2-2.8 x 0.1- 0.3 cm, abaxially glandular (uniseriate 6- to 10-celled trichomes with a glandular head), adaxially glabrescent; margin dentate with 3 subulate teeth per side. Corolla bright orange or yellow, tube cylindrical, dorsally slightly recurved, 4.3- 5.5 cm long, 0.6- 0.8 cm wide along the tube, 2.7- 3.0 cm wide at the limb, slightly constricted basally 0.5- 0.6 cm wide; base dorsally gibbous 0.3 x 0.4 cm ; limb bilabiate, upper lip formed by the two dorsal lobes, lower lip formed by the other three; lobes unequal, two dorsal connate, erect, acuminate, 0.5 x 0.3 cm ; two lateral spreading, acute, 1.2 x 0.3 cm ; the ventral lobe slightly reflexed, acute, 1.0 x 0.3 cm ; corolla outside pilose glandular (uniseriate 6- to 10-celled trichomes ending in a glandular head), with a tuft of hairs outside apically crowded in the lateral lobes, inside glabrous and with 4 appendages. Androecium of 4 stamens, filament 4.0- 4.8 cm long, with short trichomes at the base of the filaments, basally connate for 1.0- 1.2 cm of their length forming a staminal blade folded and open dorsally, anther rectangular 2.5 x 1.5 mm . Gynoecium with ovary conical, 0.5 x 0.3 cm, setulose, apically densely sericeous (uniseriate 3-to 6-celled trichomes); style 4.4- 5.0 cm long, glabrous; stigma bilobate. Nectary of one bilobulate dorsal gland, 3 x 3 mm . Fruit a green, ovoid berry, 1.2-1.5 x 0.7- 1 cm . Seeds brown, elliptic, obliquely striated, 1.5 x 0.5 mm.
Etymology. The species name makes reference to the long caudate apex of the larger leaf at each stem node.
Phenology: Flowers have been recorded on the specimens collected in February, March and November. Fruits were recorded from two specimens collected in February and March.
Distribution: Colombia (Antioquia, Chocó, Risaralda, Valle del Cauca), 400- 1900 m alt. This species is distributed along a large part of the Colombian Biogeographical Chocó region. The species seems to be distributed through a corridor of wet and cloud forest on the mountains in the "Cordillera Occidental" of Colombia in the Departments of Antioquia, Chocó, Risaralda, and Valle del Cauca.
Representative specimens: COLOMBIA. Antioquia: Parque Nacional Natural Las Orquídeas, trail from Venados to Carauta on the margin of the Río Venados, 1300- 1350 m, Feb. 15, 1989, Cogollo, A. et al. 4031 (JAUM). Area around community called La Madrona, 6-8 hrs walk from La Clara which is 3 hrs From El Sireno, which is 2 hrs on bus north of Urrao, 1050- 1200 m, Nov. 10, 1982, Folsom, J. P. 10582 (US). Chocó: Municipio San José del Palmar, river basin of river Torito, tributary of the river Hábita, 850- 950 m, Mar. 16, 1980, Forero, E. et al. 7398 (COL). Municipio of Nóvita, north slope of Cerro Torrá, western edge of the river Surama, trail to the Alto del Oso, 600- 900 m, Feb. 22, 1977, Forero, E. et al. 3128 (COL). Risaralda: Municipio of Mistrató, 12 Km NE from the municipality headwaters, 1800-1900m, Mar. 7, 1991, Galeano, G. et al. 2575 (COL). On the road San Antonio de Chami and Mistrató, Quebrada Sutu, 1750 m, Apr. 27, 1992, Fernández, A. et al. 10214 (COL).
Distinctive features: Columnea caudata is easily recognized by having a dorsiventral vegetative shoot, the pair of leaves at each node are strongly anisophyllous, the smaller leaf being an inconspicuous linear scale leaf of 0.5-0.7 x 0.1 cm is the smallest leaf known in any species of Columnea section Collandra. The apex of the larger leaf is long acuminate and 1.4- 2.8 cm long giving to the plant a characteristic appearance making it easy to recognize the species. The bracts are reduced, lanceolate to linear, most of the time absent, suggesting that they are caducous. Unlike other species in section Collandra in which the flowers are covered and protected by the bracts and are kept very close to the vegetative shoot, in this species the reduced bracts and long pedicels allow the flowers to be exserted from the vegetative shoot. The corolla limb is bilabiate but narrow, which points out a specialization to restrict the nectar only to hummingbirds with long bills. It is interesting that a narrow bilabiate limb neither allows hummingbirds with short bills (e.g. C. ericae) nor bees to access the nectar. Besides, the flowers of C. caudata present trichomes at the edges of the lateral limbs, adding to the zygomorphic symmetry of the limb; the long and narrow corolla suggests pollination exclusively by a long-billed hummingbird, and the limb design might indicate a visual signal directed to a specific species of hummingbird. This conjecture might be found to be true if the species is a plentiful nectar producer and has other ornithophilous plants competing for the same bird pollinator in the same area. A reliable nectar producing plant might increase its fitness (in, for example, pollinator visits, temporal flower fidelity, fertilization, and outcrossing) by being distinctive from other congenerics which may be less competitive. Another morphological novelty reported in C. caudata is the four internal appendages, of unknown function, located inside the corolla at two heights (Fig. 1). All of these floral traits broaden the diversity of floral architectures known for the genus Columnea and pose the question about their functional value.
Columnea megafolia L. E. Skog & M. Amaya, sp. nov. Figure 2
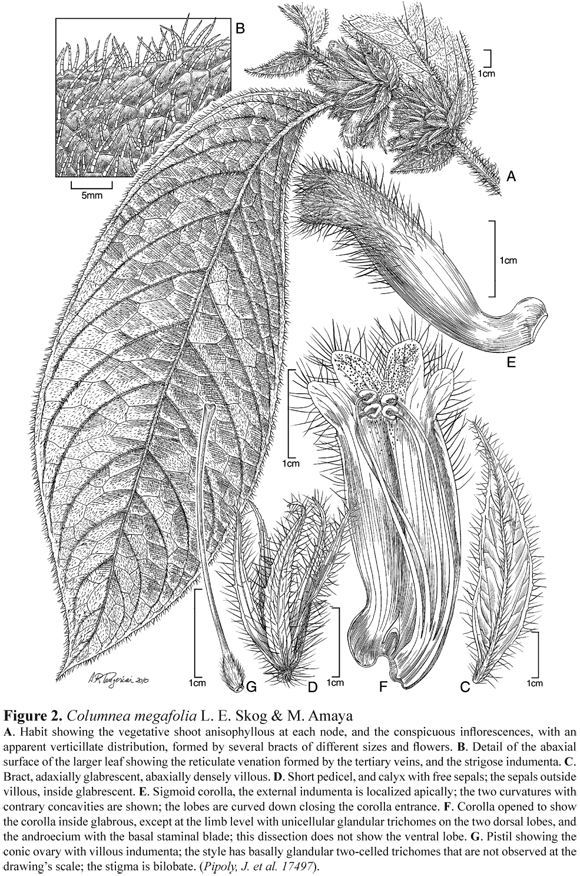
TYPE: COLOMBIA: Antioquia, municipio Urrao: Vereda Calles, Alto de Palmitas 1 km from the Inderena`s cabin at Calles, 76º19' W, 6º32' N, 1300- 1400 m, Pipoly, J. et al. 17497 (Holotype: US, Isotype: MO).
Columnea megafolia differs from C. purpurata by having larger leaves and flowers, corolla red or apically red and yellow, with a marked zygomorphic limb.
Suffrutescent terrestrial, 1 m tall or woody climber≥ 1.5 m . Stem subterete, 1 cm diam., indument lanate (trichomes 10-celled), internodes 1- 7 cm long. Leaves opposite, strongly anisophyllous in a pair, membranous, large leaf petiolate, petioles 0.2- 1.8 cm, villous (9- to 11-celled trichomes); blade asymmetrical, oblanceolate to narrowly oblong, 31-51 x 10- 16 cm, base oblique, shorter side cuneate, longer side rounded, apex acuminate, margin serrate; adaxially green, sparsely strigose (trichomes 5- to 12-celled trichomes); abaxially pale green, sparsely strigose (5- to 12-celled trichomes), 13-16 secondary veins, the tertiary veins forming a reticulate pattern; smaller leaf in a pair sessile, blade notably asymmetric, lanceolate, 2-3 x 0.4- 1.2 cm, base oblique, shorter side obtuse, longer side auriculate, apex acuminate, margin dentate, adaxially green, sparsely strigose (9- to 12-celled trichomes), abaxially pale green, sparsely strigose (10- to 12-celled trichomes). Inflorescence fasciculate, of 4-5 flowers in the axil of larger leaf; bracts 8-12, lanceolate, 1.8-5 x 0.3- 1.1 cm, abaxially red, villous (8- to 12-celled trichomes), adaxially glabrous, margin dentate. Flower shortly pedicellate, pedicels 0.1- 0.8 cm long, villous, glandular near the calyx. Calyx red, sepals free, subequal, lanceolate, 1.5-2 x 0.1- 0.4 cm, abaxially villous (8- to 12-celled trichomes), adaxially glabrous, margin dentate (sometimes with three subulate teeth per side). Corolla red, or red and apically yellow, corolla tube sigmoid, 3.5- 3.8 cm long, 0.7- 0.8 cm wide at the middle, constricted at base 0.4- 0.5 cm, base dorsally gibbous, gibbosity 0.3-0.5 x 0.5- 0.7 cm ; zygomorphic, curved downward, lobes subequal, the two dorsal obtuse, slightly hooded, imbricate and curved down, 3 x 2 mm ; the two lateral obtuse, 3 x 2 mm ; and the ventral rounded 2 x 2 mm ; corolla outside villous more dense apically (trichomes 10- to 12-celled), inside glabrous, except on the two dorsal lobes that are densely glandular (with unicellular trichomes). Androecium of 4 stamens, filament 3- 3.4 cm long, glabrous basally connate by 0.5- 0.7 cm of their length and forming a staminal blade; anther rectangular 2.5 x 1.5 mm, connective rectangular. Gynoecium with ovary conical, 0.4-0.5 x 0.1- 0.2 cm, villous (trichomes 10-celled) and basally setulose (unicellular trichomes); style 3.3 cm long, basally glandular (2-celled trichomes); stigma bilobate. Nectary of one tridentate dorsal gland, 1.5 x 3 mm . Fruit and Seeds not seen.
Etymology: The species name refers to the large size of the larger leaf at each node.
Phenology: Flowers have been recorded from specimens collected in February, March, May, June, July and December.
Distribution: Colombia (Antioquia) at 1240- 1460 m alt. in Andean Premontane Wet Forest on the western slopes of the" Cordillera Occidental ". This species seems to be endemic to the National Natural Park Las Orquídeas.
Representative specimens: COLOMBIA. Antioquia: Municipio de Urrao Parque Nacional Natural Las Orquídeas, sector Calles, 1240 m, 29 May 1988, Cogollo, A. & J. G. Ramírez 3037 (COL, JAUM); margin of river Calles, 1 km ahead from the confluence of the rivers Polo and Calles, 76º19` W, 6º32' N: 1460 m, Feb. 11 1989, Cogollo, A. et al. 3896 (JAUM); 1380 m, Mar. 26 1988, Cogollo, A. et al. 2622 (COL, JAUM); 1350- 1450 m, Jul. 6 1991, Cogollo, A. et al. 5009 (MO); 76º19` W, 6º31' N: 1300- 1320 m, Mar. 27 1991, Ramírez, J. & E. Muñoz 4045 (MO); river Calles, 77º4'51'' W, 4º35'56'' N, 1400- 1500 m, May 3 1995, Fonnegra, R. et al. 5536 (HUA); 1400 m, Jun. 15 1981, Albert de Escobar, L. 1714 (HUA).
Distinctive features: Columnea megafolia is close to C. purpurata, and both species have conspicuous orange bracts that usually dry purplish, both species do not have red spots on the larger leaves, which is unusual for species of section Collandra. Also both species have a tridentate dorsal nectary gland. However C. megafolia can be recognized by having robust shoots and villous indumenta on the surface of almost all organs. The larger leaf at each node is very large, 31-51 x 10- 16 cm (vs. 11-30 x 3- 12 cm in C. purpurata), usually with 15 veins on the larger side of the blade (vs. 9- 12 in C. purpurata); short petioles give the appearance of the leaves being sessile; the margin is serrate with glandular teeth, whose function might be to release excess water. The inflorescences are very conspicuous both in color which is red, and size; although the inflorescence buds are located in the axils of the larger leaf of each node, their size makes their distribution around the node appear verticillate; the bracts are numerous and of different sizes, having the same red color of the calyx and corolla, increasing the visual signal for pollinators. The corolla tube is sigmoid presenting two marked curvatures along the tube, i.e., one concave up, basally just before the gibbous, the other concave down, apically before the corolla limb; the corolla limb is curved downward closing the corolla entrance (vs. an open corolla in C. purpurata). This trait might be a trait to deter the entrance of certain floriverous insect - larvae of an unknown insect were found inside the corolla of several dissected flowers. The corolla limb is clearly zygomorphic (vs. an actinomorphic limb in C. purpurata), however the limb is not bilabiate as all the lobes are recurved down to close the corolla entrance, instead of forming upper and lower lips (Fig. 2). As mentioned before, C. megafolia has been found only in Colombia (Antioquia) at 1200- 1460 m, while C. purpurata has a Neotropical distribution and has been collected in Mexico, Nicaragua, Costa Rica, Panama, Venezuela, and Colombia at 80- 1750 m .
ACKNOWLEDGMENTS
The authors thank National University of Colombia, University of Aarhus, and Smithsonian Institution for the opportunity to do research, Juan Carlos Pinzón and Alice Tangerini for the elaboration of the drawings, and to the directors of the herbaria of COL, HUA, JAUM, MO, and US for their support and collaboration to consult the collections deposited in those herbaria. To Jim Smith and another anonymous reviewer whose valuable comments helped to improve the manuscript.
LITERATURE CITED
1. Amaya-Márquez, M. 1996. Sistemática y polinización del género Columnea (Gesneriaceae) en la Reserva Natural la Planada (Nariño). Tesis de maestría, Universidad Nacional de Colombia, Bogotá, Colombia. [ Links ]
2. Amaya-Márquez, M. 2010a. Una nueva especie de Columnea (Gesneriaceae) de la cordillera Oriental en los Andes Colombianos. Caldasia 32 (1): 113-116. [ Links ]
3. Amaya-Márquez, M. 2010b. Novedades Taxonómicas en el género Columnea (Gesneriaceae). Rev. Acad. Colomb. Ci. Ex. Fis. Nat. 34 (132): 301-307. [ Links ]
4. Amaya-Márquez, M. & J. L. Clark. 2011. Columnea bivalvis (Gesneriaceae), a new species from the eastern slopes of the Ecuadorian Andes. J. Bot. Res. Inst. Texas 5 (1):75-79. [ Links ]
5. Amaya-Márquez, M. & O.H. Marín-Gómez. 2012. Columnea rangelii (Gesneriaceae), a new species from the Serranía de los Paraguas in the Colombian Andes. Caldasia 34 (1): 69-74. [ Links ]
6. Amaya-Márquez, M. & J.F. Smith. 2012. A rare new species of Columnea (Gesneriaceae) from" Cordillera Occidental " in the Colombian Andes. Rev. Acad. Colomb. Ci. Ex. Fis. Nat. 23 (139): 13-16. [ Links ]
7. Amaya-M, M, L.E. Skog & L.P. Kvist. 2004. Novae Gesneriaceae Neotropicarium XIII: Four New Species of Columnea (Gesneriaceae) Section Collandra from Colombia. Edinburgh J. Bot. 60: 415-424. [ Links ]
8. Clark, J.L. & L. Clavijo. 2012. Columnea antennifera, a new species of Gesneriaceae from the Cordillera Central of the Colombian Andes. J. Bot. Res. Inst. Texas 6(2):385-389. [ Links ]
9. Dyer, L.A., M.S. Singer, J.T. Lill, J.O. Stireman, G.L. Gentry, R.J. Marquis, R.E. Ricklefs, H.F. Greeney, D.L. Wagner, H.C. Morais, I.R. Diniz, T.A. Kursar & P.D. Coley. 2007. Host specificity of Lepidoptera in tropical and temperate forest. Nature 448: 696-699. [ Links ]
10. Jones, C.E. & P.V. Rich 1972. Ornithophily and extrafloral color patterns in Columnea florida (Morton) Morton (Gesneriaceae). Bull. S. Calif. Acad. Sci. 7: 220-243. [ Links ]
11. Kastinger, C. & A. Weber. 2000. Attraction of hummingbirds by extrafloral cues in some Costa Rican Species of Columnea (Gesneriaceae) Linzer Biol. Beitr. 32(2): 652-653. [ Links ]
12. Kriebel, R. 2005. A new species of Columnea and range extension in the Gesneriaceae from Costa Rica. Brittonia 57(1): 39-42. [ Links ]
13. Kvist, L.P. & L.E. Skog. 1993. The genus Columnea (Gesneriaceae) in Ecuador. Allertonia 6: 327-400. [ Links ]
14. Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. de Fonseca & J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. [ Links ]
15. Morley, B.D. 1971. A hybrid swarm between two hummingbird-pollinated species of Columnea (Gesneriaceae) in Jamaica. J. Linn. Soc. Bot. 64: 81-96. [ Links ]
16. Rangel-Ch., J.O & O. Rivera-Díaz. 2004. Diversidad y riqueza de espermatófitos en el Chocó Biogeográfico: 83-99. In: Rangel-Ch.,J.O. (ed.). Colombia Diversidad Biótica IV. El Chocó Biogeográfico/Costa Pacífica. Universidad Nacional de Colombia. Bogotá [ Links ].
17. Schemske, D.W. 2002. Ecological and evolutionary perspectives on the origins of tropical diversity. pp. 163-173. In: Chazdon, R.I. & T.C. Whitmore (eds.). Foundations of Tropical Forest Biology. Univ. Chicago Press. Chicago, Illinois, USA. [ Links ]
18. Schemske, D.W. 2009. Biotic interactions and speciation in the tropics: 219-239. In: Butlin, R.K., J.R. Bridle & D. Schluter (eds.). Speciation and patterns of diversity. Cambridge University Press, Cambridge. [ Links ]
19. Schemske, D.W., G.G. Mittelbach, H.V. Cornell, J.M. Sobel & J.M. Roy. 2009. Is there a latitudinal gradient in the importance of biotic interactions? Ann. Rev. Ecol. Evol. Syst. 40: 245-269. [ Links ]
20. Schuchman, K.L. 1999. Trochilidae. Handbook of the Birds of the World. Volume 5. Lynx Editions. [ Links ]
21. Skog, L.E. & J.K. Boggan. 2007. World Checklist of Gesneriaceae. Washington, DC : Dept. of Botany, Smithsonian Institution Washington, DC. http://botany.si.edu/Gesneriaceae/Checklist [ Links ]
22. Stiles, F.G. & C.E. Freeman. 1993. Patterns in floral nectar characteristics of some bird-visited plant species from Costa Rica. Biotropica 25:191-205 [ Links ]
23. Thompson, J.N. 1994. The Coevolutionary Process. Univ. Chicago Press, Chicago, Illinois, USA. [ Links ]
