










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.36 no.1 Bogotá Jan./June 2014
https://doi.org/10.15446/caldasia.v36n1.43894
http://dx.doi.org/10.15446/caldasia.v36n1.43894
JULIO MARIO HOYOS
CATALINA MANTILLA
DIANA GALINDO
LINA SALGAR
Pontificia Universidad Javeriana, Departamento de Biología, Unidad de Ecología y Sistemática (UNESIS), Apartado 56710, Bogotá Colombia. jmhoyos@javeriana.edu.co, dmant010@fiu.edu, linasalgar@hotmail.com
Universidad de los Andes, Departamento de Biología, Bogotá, Colombia. dmgalindo48@uniandes.edu.co
ABSTRACT
We present a phylogenetic analysis within the Pristimantis unistrigatus group (Anura, Craugastoridae) of Colombia . Characters from the superficial muscles of the hands and feet as well as external characters were taken for analysis. Most of the muscle characters were observed directly, and some were taken from the literature. Similarly, the external ones were taken mostly from the original descriptions and others from the literature as well. Two matrices were constructed, as the species belonging to this group have changed in recent years with respect to the initially proposed when the group was defined. The results lead us to conclude that the group is not monophyletic, although there are some relationships that are worth to survey because they are kept in the very last cladograms obtained for both proposals. It is suggested that these last relationships should be explored in particular, and the overall group in general, increasing the number of characters and taxa that belong to P. unistrigatus. An open question we left is whether actually is worth to keep these informal taxonomic hierarchy called group within the genera of anurans.
Key words. Neotropical frogs, Cordillera Oriental, superficial muscles, Pristimantis.
RESUMEN
Se presenta un análisis filogenético del grupo Prsitimantisunistrigatus (Anura, Craugastoridae) de Colombia. Para esto se tomaron caracteres de la musculatura superficial de manos y pies y caracteres externos. La mayoría de los caracteres musculares fueron observados directamente, y algunos fueron tomados de la literatura. Igualmente, los externos fueron tomados en su mayoría de las descripciones originales, y otros tomados de la literatura también. Se construyeron dos matrices, pues las especies que pertenecen al grupo han cambiado en los últimos años con respecto a lo propuesto inicialmente cuando el grupo fue definido. Los resultados nos llevan a concluir que el grupo no es monofilético, aunque existen algunas relaciones que vale la pena explorar, pues se mantienen en los cladogramas definitivos obtenidos para ambas propuestas. Se sugiere que deben explorarse más estas relaciones en particular, y la del grupo en general, aumentando los caracteres y el número de taxones que pertenecen a P. unirtsigatus. Igualmente se deja como interrogante si en realidad vale la pena mantener estos grupos informales dentro de los géneros de anuros.
Palabras clave. Ranas neotropicales, Cordillera Oriental, músculos superficiales, Pristimantis.
Recibido: 12/12/2011
Aceptado: 25/02/2014
INTRODUCTION
Frost et al. (2006) doubted the existence of the genus Eleutherodactylus Duméril and Bibron, 1841 as monophyletic, and suggested that it should be divided into many monophyletic genera. Heinicke et al. (2008) divided the former genus Eleutherodactylus plus the genus Brachycephalus Fitzinger, 1826 in three Clades: the South American Clade, the Caribbean Clade and the Middle American Clade. The Eleutherodactylus unistrigatus group was included in the South American Clade using the available name Pristimantis Jiménez de la Espada, 1870. This group is one of the richest Pristimantis groups, with members occurring in the Andes and the Amazonian basin, ranging from Bolivia to Colombia (Hedges et al. 2008, Padial et al. 2009).
In the literature related to the diagnosis of the unistrigatus group species of the Cordillera Oriental of Colombia, potential kinship and characteristics in common are referred (Duellman & Simmons 1977, Lynch1973, 1978, 1981, 1983, 1984, 1994a, 1994b, 1998, 2003, Lynch & Duellman1980, Lynch & Duellman 1997, Lynch & Suarez 2002, Pyburn & Lynch1981). However, there have been no formal relationship hypotheses supported by cladograms.
It is perhaps for this Pristimantis group that were proposed the highest number of hypotheses of relationships in Colombia , because it is the richest in species number in this country. From the perspective of phylogenetic systematics of the genus Pristimantis, the work that has a growing number of Colombian species for the unistrigatus group (sensu Lynch & Duellman 1997) of the Cordillera Oriental of Colombia has been that of Galindo (2004), based on external characters. However, the papers of Duellman & Simmons (1977), Lynch (1973, 1975a, 1975b, 1978, 1981, 1983, 1984, 1994a, 1994b, 1998, 2003), Lynch & Duellman (1980), Lynch & Suárez (2002 ) and Pyburn & Lynch (1981) have revealed potential relationships for species of the unistrigatus group, some with formal cladograms, but the majority without them. Lynch (1984) proposed a hypothesis of relationship for the unsitrigatus group without including any of the species used here, but some belonging to the "assemblages" pyrrhomerus and myersi, suggesting these should be combined by a series of synapomorphies found in external characters. Finally, recently Padial et al. (2009) considered that the Pristimantis unistrigatus group is not monophyletic.
Other groups, which have launched hypotheses of relationship, either with or without explicit cladograms are: discoidalis (Lynch 1989) (with explicit cladogram), using external and cranial characters, and sulcatus (Lynch 1997) (with explicit cladogram), finding for these relationships nine characters from the skull. As this paper aims to use morphology as a fundamental element to establish kinship hypothesis, the discussion will not be based on specific morphological differences between what we found and what other authors have found, but between the relationships proposed here based on potential synapomorphies, and other hypotheses and characters used to construct them.
From morphology's point of view, the contributions are all innovative in the sense that no previous research has been dedicated solely to the study of muscles of hands and feet for the phylogenetic analysis of the Pristimantis genus. Similarly, although there are some previous descriptions about hand muscles of some species of the genus (e.g.Burton 1998a, Dunlap 1960), these were mainly for comparative purposes (Dunlap 1960, Salgar et al. 2009), or with the aim to find characters to establish hypotheses of relationships (Burton 1998a) but there was not a genus-specific analysis performed.
Previous studies in the literature, using both muscles of hands and forearms and feet to propose phylogenetic relationships in frogs are very rare; in fact, there is only one (Faivovich 2002), but in the genus Scinax (Hylidae). The most comprehensive descriptive work in this regard have been those of Gaup (1886), Dunlap (1960), Andersen (1978) and, more recently, Burton (1996,1998a,b, 2001, 2004). Most of them propose (tacitly or explicitly) some assumptions about the potential systematic value of characters taken from hands and feet in anuran amphibians, without reaching cladistic analysis themselves. Only in one of his most recent work, Burton (2004) developed a cladistic analysis using only the muscles of the feet in the family Hylidae. The only existing descriptions of this musculature in Colombian species of frogs has been conducted by Salgar (2003) and Salgar et al. (2009). Although these last two analyses are brief, for the first time, possible synapomorphies for the genus Pristimantis (intrageneric relationships) based on the muscles of hands and feet were revealed. Lynch (1984) proposed some characteres but for relationships between species groups, based on osteological characters and some of the muscles (but not hands and feet).
Hedges et al. (2008) redefined the Pristimantis unistrigatus group changing the species included to it, assigning 193 species to this group, changing some of the species previously identified into the group by Lynch (1976a, Lynch & Duellman 1997, Duellman & Pramuk 1999), and by Lynch & Duellman (1997) and Acosta (2000) for Colombia.
Taking into account that there are two different classifications for the Pristimantis unistrigatus group, our aim is to compare our results in using morphological characters from hand and foot muscles in both proposals, because there are not enough consensus about which is the best classification. It is for this reason that both the number of species and characters in both analyses differs (see results). The only classification that include all species of our study is that by Lynch & Duellman (1997). The number of species in classification by Hedges et al. (2008) is lower, because they do not consider some of the species by Lynch & Duellman (1997) as belonging to the P. unistrigatus group. Owing to this account, the number of characters also changed. In comparing our results with those by Heinicke et al. (2008), and Hedges et al. (2008), we can conclude that they agree that P. unistrigatus group is not monophyletic.
MATERIALS AND METHODS
A total of 42 adult specimens (Table 1) representing 17 species of Pristimantis of the Cordillera Oriental of Colombia and one of Craugastor were dissected. Specimens belong to the Museo de Historia Natural (MUJ) of the Pontificia Universidad Javeriana in Bogotá, Colombia , and to the Instituto de Ciencias Naturales (ICN) of the Universidad Nacional de Colombia in Bogotá.
Taking into account the species belonging to the unistrigatus group sensu Hedges et al. (2008), we decided to compare results considering the former classification made by Lynch & Duellman (1997).We chose the following two ingroups: sensu Lynch & Duellman (1997) (LD): Pristimantis anolirex (Lynch, 1983): ICN 33522, 15444; P. bacchus (Lynch, 1984): ICN 33164, 33170, 34232; P. bicolor ( Rueda-Almonacid and Lynch, 1983): ICN 14474, 26321; P. bogotensis (Peters, 1863): MUJ 052, 465, 481, 687; P. elegans (Peters, 1863): MUJ 036; P. frater (Werner, 1899): ICN 40579, 49911; P. jorgevelosai (Lynch, 1994): ICN 15299, 15318; P. lynchi (Duellman and Simmons, 1977): MUJ 1480, 1538, 1507, 2036; ICN 01932, 01937; P. merostictus (Lynch, 1984): ICN 34233, 34235; P. miyatai (Lynch, 1984): ICN 50062, 50065, 50069; P. nervicus (Lynch, 1994): MUJ 023,1176; P. nicefori (Cochran and Goin, 1970): ICN 20994, 21119; P. prolixodiscus (Lynch, 1978): ICN 10104, 10107; P. pugnax (Lynch, 1973): ICN 22981, 23187; P. spilogaster (Lynch, 1984): ICN 12445, 12446; P. taeniatus (Boulenger, 1912): MUJ 2452, 2420; P. tamsitti (Cochran and Goin, 1970): ICN 22949, 22951, 23638; Sensu Hedges et al. (2008) (HEA): P. anolirex (Lynch, 1983), P. bacchus (Lynch, 1984), P. bogotensis (Peters, 1863) , P. elegans (Peters, 1863), P. lynchi (Duellman and Simmons, 1977), P. merostictus (Lynch, 1984), P. nervicus (Lynch, 1994), P. nicefori (Cochran and Goin, 1970), P. pugnax (Lynch, 1973), P. spilogaster (Lynch, 1984), P. taeniatus (Boulenger, 1912), P. tamsitti (Cochran and Goin, 1970) .
Outgroups: Craugastor raniformis: MUJ 2871, 2689 for the LD matrix, and P. frater for the HEA matrix (Table 2 and Table 3).
The new classification proposed by Hedges et al. (2008) for some of the species formerly assigned to unistigatus group is as follows: P. frater: frater group; P. bicolor: species series P. (Hypodiction) ridens; P.miyatai: P. frater group; P. jorgevelosai: species series P. (Hypodiction) ridens; and P. prolixodiscus: lacrimosus group.
Muscles
We observed about 90 small hand and feet muscles, both superficial and deep, of which about 50 superficial were chosen. For choosing the characters and states, we did comparisons between muscles from ten species of Pristimantis used for the study with respect to different aspects that are related below. Dissections were done in hands, feet, and legs and forearms in order to examine those muscles associated morphologically with the hands and feet. We used a dissecting microscope and a reversible Weigert's iodine solution that stains muscles (Bock & Shear1972) to see the direction of the fibers, obtaining a clear differentiation of muscles and tendons, including the origins and insertions. For all specimens both right and left hands and feet superficial muscles were dissected and examined, following the protocols proposed by Raikow et al. (1990). For each muscle we identified: a. origin; b. insertion; c. spatial arrangement with respect to the hand and foot, and d. spatial relationship with adjacent muscles. Although we explored mainly superficial musculature, we also dissected some deep muscles considering that we took into account characters proposed by Burton (1998a, 2004). The nomenclature was taken from that proposed by Dunlap (1960), Andersen (1978), Burton (1998a, 2004), and Salgar et al. (2009). We used for most cases, at least one male and one female of each species to the extent that the number of specimens of the collection allowed it, to determine the existence of sexual dimorphism. Finally, we drew pictures of characters considered as potential synapomorphies using photographs and the Adobe Illustrator software.
For the accuracy of the origin and insertion of muscles, we made use of clearing and staining skeletons of P. bogotensis.
External characters
These were taken from taxonomic descriptions after Cochran & Goin (1970), Lynch (1983), Lynch (1984), Rueda & Lynch (1983), Dunn (1944), Hoyos (1991), Pyburn & Lynch (1991), Lynch (1994), Duellman & Simmons (1997), Lynch (1998), Lynch (1978), and Lynch & Duellman (1980), and after our own observations.
Phylogenetic analysis
For the phylogenetic analysis, we compared each result taking into account two matrices (see below). We did heuristic searches in TNT, performing TBR branch swapping on 100 addition replicates. Matrices were running considering characters with the same weight. We did not find many intraspecific (polymorphic) variations or many multistate characters, however we use all of those we found for the phylogenetic analyses. Polymorphic characters were coded according to Faivovich (2002) as follows: A=0/1; B=0/2, and C=1/2, and multistate characters were all considered nonadditive and unordered. We did not get whether retention (RI) or consistency indices (CI), given that we are comparing groups of different sizes (e.g. Conrad 2008), so the value for the CI will be greater in the HEA ( Sensu Hedges et al., 2008) group than in the LD (sensu Lynch & Duellman, 1997) group because we considered less species in the first group than in the second one, and the RI will be less in the HEA for the same reason.
We choose one outgroup species for each analysis, following recommendation by Nixon & Carpenter (1993: 419) about the fact that " a third pervasive myth, the notion that more than one outgroup is somehow required for correct polarity determination", and taking into account that "Outgroups and polarity are the same problem" (Nixon Nixon & Carpenter 1993: 420). We only show the unambiguous synapomorphies.
Results
We scored 81 characters, from which 64 are muscle characters (44 from the manus and 20 from the pes), and 17 external characters. Four manus characters were taken from Burton (1998a), and five pes characters from Burton (2004). Based on this matrix, we eliminated those characters according to changes in the taxa for each matrix as we describe below.
Phylogenetic analysis
LD P. unistrigatus group
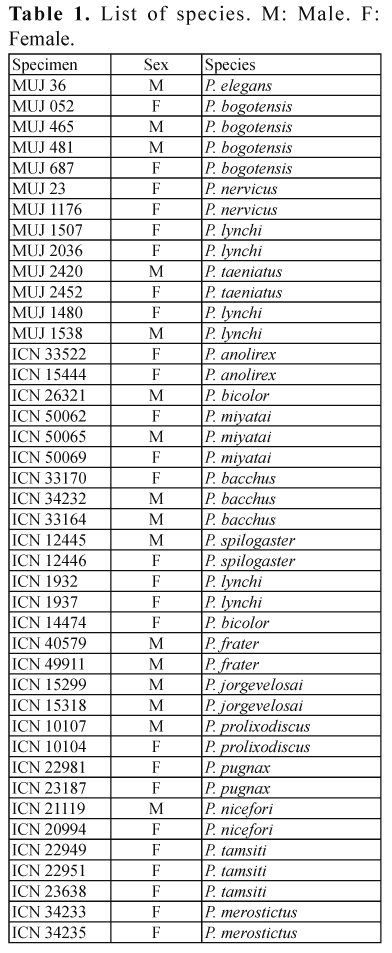
By eliminating those characters with only one character state present in the outgroup and ingroup, we took 70 from the 81 characters (Appendix 1). The cladistic analysis resulted in one fully resolved most-parismonious tree (Fig. 1) with a length of 202. We found character states 62(0) (First finger shorter than second) and 23(1) (The origin of the m. adductor policis is not covered by the aponeurosis palmaris) (Fig. 2) as the only two unambiguous putative synapomorphies. The first one included almost all species leaving away P. nicefori, and the second one is a character shared by P. prolixodiscus and P. bicolor.
HEA P. unistrigatus group (Appendix 2)
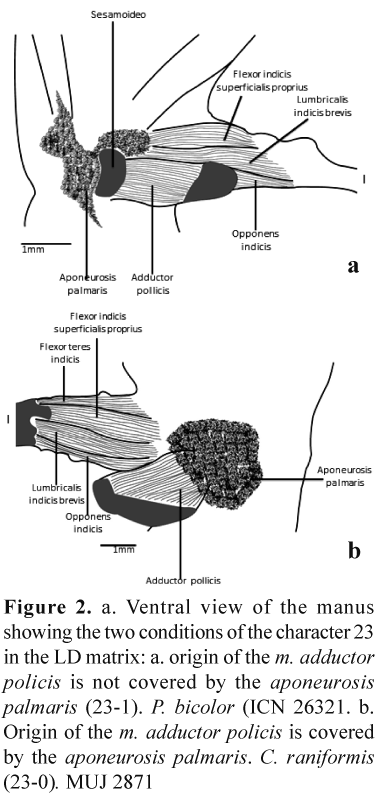
We took 60 characters for the analysis (Appendix 2). The cladistic analysis resulted in 20 most-parismonious trees of 139 steps. The strict consensus tree (Fig. 3) shows a node 14 not resolved, consisting of taxa P. lynchi, P. nervicus, P. elegans, P. bogotensis, P. nicefori, P. anolirex, P. bacchus, and P. taeniatus.
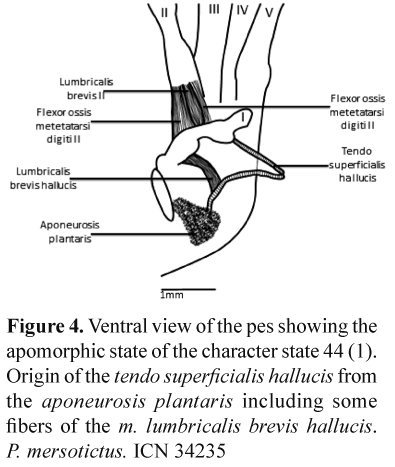
The only unambiguous putative synapomorphy (found in all trees) is the character state 44(1) (Origin of the tendo superficialis hallucis from the aponeurosis plantaris including some fibers of the m. lumbricalis brevis hallucis) (Fig. 4) relating P. spilogaster and P. merostictus.
DISCUSSION
As shown in the cladograms obtained for both unistrigatus groups, the LD (Fig. 1) group is more resolved than the strict consensus of the HEA (Fig. 3), but there are no synapomorphies that support it, except partially for nodes 21 and 32 in LD, and exclusively for node16 in HEA, this means that all topologies are weakly supported by synapomorphies. In node 22 of DL no synapomorphies were found, whereas for the other nodes, potential synapomorphies are actually reversals or convergences.
Taking into account the results and species considered by HEA and LD, we could say that the assigned HEA unistrigatus group is weaker because the relationships are less resolved, even if the number of steps is smaller (it has a smaller number of taxa). On the other hand, even with a larger number of species involved, the proposed LD unistrigatus group can be considered stronger if we take the fact that we have got a single tree completely resolved; this does not mean that we are clear about the putative synapomorphies involved, as we saw above. It is remarkable that the relationships between P. pugnax and P. tamsitti, and between P. spilogaster and P. merostictus are retained in both cladograms, meaning that these species maybe considered sisters, but without finding putative synapomorphies to support the relationship between P. pugnax and P. tamsitti.
Limb muscles and phylogeny
All characters and character states obtained were chosen bearing in mind character distribution and conditions deemed to be worth for the analysis. Burton's (1998a, 2004) characters and character states were used just as he defined them, but in some cases we believed that some had to be redefined due to the fact that they were not clearly named. Then, we think that most of these characters are new and very useful for future analyses in frog morphology and phylogeny studies. It is fascinating how many characters have found on the hands and feet, although only the superficial muscles have been observed. More characters were found in the hands (44, including four of Burton) than in the feet (21, including six of Burton). Similarly, after making a clearance based on external descriptions and original observations, we obtained 26 characters. Faivovich (2002) used 14 characters from the manus and seven from the pes, from which he found two synapomorphies for Scinax Wagler, 1830 , both from the manus. However, although we did not find any useful characters taken from hand and foot musculature, this does not mean that these characters should be dismissed from phylogenetic studies in frogs. Burton (1998a) found some hand muscle character conditions useful to support intrafamiliar relationships in Leptodactylids. Likewise, Burton (1998b) found a hand muscle character condition within Ranidae Rafinesque, 1814, stressing that superficial muscles can vary more than deep ones, but that these muscle can contribute to get frog relationships. It is apparently historically clear that superficial hand muscles may play a role more important to resolve frog relationships than superficial feet muscles, although we found muscles of both that could be putative synapomorphies.
External characters
Phylogenetic analysis including external characters is rare in frogs, although identification of most of the species groups and all species have been based on these characters. The results show that characteristics of the fingers are still important, and can be used in establishing kinship, but were obviously not essential to support the monophyly of the unistrigatus group. Apparently, the other external characters are not useful because, in any case, it was established that they could participate as potential synapomorphies. Maybe we should pay attention to the condition of tympanic annulus and to the dorsal view of the snout because for the two analyses that show that these characters appear as potential synapomorphies (nodes 21 and 23 respectively in LD group, and node 15 in HEA group) , although actually they are homoplasies.
To compare our results with those by Heinicke et al. (2008) and Hedges et al. (2008) is difficult, because most of the species included in our work were not used by these authors. However, our results agree in identifying the unistrigatus group as non monophyletic. Heinicke et al. (2008) only included six spececies belonging to the former P. unistrigatus group (Lynch, 1976; Lynch et al., 1997). Taking into account this classification, their results showed that this group is monophyletic. However, based on Hedges et al. (2008) classification P. ockendeni, in the P. unistrigatus group by Heinicke et al. (2008), belongs to the P. frater group; in this sense, the group would become not monophyletic.
CONCLUSIONS
Although the muscles of hands and feet and external characters useful in taxonomy failed to identify the Pristimantis unistrigatus as monophyletic, this does not mean they are not useful for establishing relationships within the genus. It is possible that including more unistrigatus group species in the study, the situation changes, however, it seems also essential to include the deep muscles of the hands and feet and external characters as well. It is possible that other characters facilitate to define relationships among species within the genus. However, it is necessary to explore deeper the relationship P. spilogaster-P. merostictus, and P. pugnax-P. tamsitti, because we need whether to confirm or falsify if these clades actually are monophyletic. By detecting the difficulties that several researchers (e,g, Hedges et al. 2008; Padial et al. 2009) and our own study have found to discover the monophyly of the P. unistrigatus group and other ones in frogs, the question we pose is: Is it worthwhile to keep this informal taxonomic hierarchy called group in systematics of frogs?
LITERATURE CITED
1. Acosta, A. R. 2000. Ranas, Salamandras y Caecilias (Tetrapoda: Amphibia) de Colombia. Biota Colombiana 1 (3): 289-319. [ Links ]
2. Andersen, M. 1978. The comparative myology and osteology of the carpus and tarsus of selected anurans. Volumes I and II. Ph.D. Dissertation.University of Kansas, Department of Systematics and Ecology. Kansas, USA . 310pp. [ Links ]
3. Bernal, X. & F. Guzmán. 1999. The advertisement calls of three Eleutherodactylus species (Anura: Leptodactylidae) in Colombian highland community. Revista Academia Colombiana de Ciencias Exactas Físicas y Naturales 23(suplemento especial): 261-264. [ Links ]
4. Bock, W. & O.R. Shear. 1972. A staining method for gross dissection of vertebrate muscles. Anatomischer Anzeiger 130: 222-227. [ Links ]
5. Burton, T.C. 1996. Adaptation and evolution in the hand muscles of Australo-Papuan Hylid Frogs (Anura: Hylidae: Pelodryadinae). Australian Journal of Zoology 44: 611-623. [ Links ]
6. Burton, T.C. 1998a. Variation in the hand and superficial throat musculature of neotropical Leptodactylidae frogs. Herpetologica 54: 53-72. [ Links ]
7. Burton, T.C. 1998b. Pointing the way: the distribution and evolution of some characters of the finger muscles of frogs. American Museum Novitates 3229: 1-13. [ Links ]
8. Burton, T.C. 2001. Variation in the foot muscles of frogs of the family Myobatrachidae. Australian Journal of Zoology 49: 539-559. [ Links ]
9. Burton, T.C. 2004. Muscles of the pes of Hylid frogs. Journal of Morphology 260: 209-233. [ Links ]
10. Cochran D.M. & C.J. Goin 1970. Frogs of Colombia . United States National Museum Bulletin 288, Smithsonian Institution Press, Washington D.C. 655 pp. [ Links ]
11. Conrad, J.L. 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin of the American Museum of Natural History 310: 1-182. [ Links ]
12. Duellman, W.D. & J.B. Pramuk. 1999. Frogs of the Genus Eleutherodactylus: Anura: Leptodactylidae) in the Andes of Northern Peru. Scientific Papers, Natural History Museum, The University of Kansas (13):1-78. [ Links ]
13. Duellman, W.D. & J.E. Simmons. 1977. A new species of Eleutherodactylus: Anura: Leptodactylidae) from the Cordillera Oriental of Colombia. Proceedings of the Biological Society of Washington 90: 60-65. [ Links ]
14. Dunlap, D. 1960. The comparative miology of the pelvic appendage in Salientia. Journal of Morphology 160: 1-76. [ Links ]
15. Dunn, E. R. 1944. Herpetology of Bogotá Area. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 6: 68-81. [ Links ]
16. Egan, D. 1995. The comparative osteology and miology of the carpus of the Australian Leptodactylidae. Honors Dissertation. University of Adelaide. Adelaide, Australia . 58pp. [ Links ]
17. Faivovich, J. 2002. A cladistic analysis of Scinax (Hylidae). Cladistics 18: 367-393. [ Links ]
18. Frost, D., T. Grant, J. Faivovich, R. Bai, A. Haas, C. Haddad, R. De Sá, A. Channing, M. Wilkinson, S. Donnellan, C. Raxworthy, J. Campbell, B. Blotto, P. Moler, R. Drewes, R. Nussbaum, J. Lynch, D. Green & W. Wheeler 2006. Tree of life. The amphibian tree of life. Bulletin of the American Museum of Natural History 297: 1–370. [ Links ]
19. Galindo, D. 2004. Filogenia de especies del grupo unistrigatus (sensu Lynch), del género Eleutherodactylus (Amphibia: Anura: Leptodactylidae) de la cordillera Oriental de Colombia. Trabajo de Grado de Pregrado. Pontificia Universidad Javeriana, Bogotá, Colombia. 190 pp. [ Links ]
20. Gaupp, E.1896. A. Ecker's und R. Wiedersheim's Anatomie des Frosches. Friedrich View und Sohn, Braunschweig. [ Links ]
21. Glaw, F.M., M. Vences, F. Andreone & D. Vallan. 2001. Revision of the Boophis majori group (Amphibia : Mantellidae) from Madagascar , with the description of five new species. Zoological Journal of the Linnean Society133: 495-529. [ Links ]
22. Hedges, S.B., W.E. Duellman & M.P. Heinicke. 2008. New World direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 1737: 1-182 pp [ Links ]
23. Heinicke, M.P., W.E. Duellman & S. B. Hedges. 2007. Major Caribbean and Central American frog faunas originated by ancient oceanic dispersal. Proceedings of the National Academy of Sciences of United States of America 104 (24): 1092-1097. [ Links ]
24. Hoyos, J.M. 1991. Aspectos taxonómicos y patrones de diseño de Eleutherodactylus bogotensis del Parque Nacional Natural Chingaza. Cuadernos Divulgativos 18: 1-12 [ Links ]
25. Kwet, A. & J. FAIVOVICH. 2001. Protoceratophrys bigibbosa species group (Anura: Leptodactylidae), with description of new species. Copeia 2001: 203-215. [ Links ]
26. Lynch, J.D. 1973. A new species of Eleutherodactylus (Amphibia, Leptodactylidae) from Andean Ecuador. Bulletin of the Southern California Academy of Sciences 72: 107-109. [ Links ]
27. Lynch, J.D. 1975a. A review of the broad-headed eleutherodactyline frogs of South America (Leptodactylidae). Occasional Papers of the Museum of Natural History University of Kansas 38: 1-46. [ Links ]
28. Lynch, J.D. 1975b. The identity of the frog Eleutherodactylus conspicillatus (Günther) with description of two related species from Northwestern Soth America (Amphibia, Leptodactylidae). Natural History Museum of Los Angeles Contributions in Science 272: 1-19. [ Links ]
29. Lynch, J.D. 1976.The species groups of the South American frogs of the genus Eleutherodactylus (Leptodactylidae). Occasional papers of the Museum of Natural History 61: 1-24. [ Links ]
30. Lynch, J.D. 1978. A new Eleutherodactyline frog from the Andes of Northern Colombia (Leptodactylidae). Copeia 1978: 17-21. [ Links ]
31. Lynch, J.D. 1981. Two new species of Eleutherodactylus from Western Colombia (Amphibia, Anura, Leptodactylidae). Occasional Papers of the Museum of Zoology, University of Michigan 697: 1-12. [ Links ]
32. Lynch, J.D. 1983. A new Leptodactylid frog from the Cordillera Oriental of Colombia. In: Rhodin, A. G. J. & Williams, E. E. Advances in Herpetology and evolutionary biology: 52-57. Museum of Comparative Zoology, Cambridge, Massachussetts. [ Links ]
33. Lynch, J.D. 1984. A new species of Eleutherodactylus (Amphibia, Anura: Leptodactylidae) from Southern Andean Colombia. Herpetologica 40: 234-237. [ Links ]
34. Lynch, J.D. 1989. Intrageneric Relationships of Mainland Eleutherodactylus (Leptodactylidae) I. A Review of the Frogs Assigned to the Eleutherodactylus discoidalis species group.Contributions in Biology and Geology, Milwaukee Public Museum 79: 1-25. [ Links ]
35. Lynch, J.D. 1994a. A new species of High-altitude frog (Eleutherodactylus Leptodactylidae) from the Cordillera Oriental of Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 19: 195-203. [ Links ]
36. Lynch, J.D. 1994b. A new species of frog (Genus Eleutherodactylus: Leptodactylidae) from the cloud forest in Departamento de Santander, Colombia . Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 19: 205-20. [ Links ]
37. Lynch, J.D. 1997. Intrageneric relationships of mainland Eleutherodactylus II. A review of the Eleutherodactylus sulcatus group. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 21: 353-372. [ Links ]
38. Lynch, J.D. 1998. A new frog (Genus Eleutherodactylus) from cloud forests of southern Boyacá. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 22: 429-432. [ Links ]
39. Lynch, J.D. 2001. Four osteological synapomorphies within Eleutherodactylus (Amphibia: Leptodactylidae) and their bearing on subgeneric classifications. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 25: 127-136. [ Links ]
40. Lynch, J.D. 2003. New species of frogs (Eleutherodactylus: Leptodactylidae) from the Cordillera Oriental of Norte de Santander and Santander, Colombia . Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 27: 449-460. [ Links ]
41. Lynch, J.D. & M.C. Ardila-Robayo. 1999. The Eleutherodactylus of the taeniatus complex in Western Colombia: taxonomy and distribution. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23(89): 615-624. [ Links ]
42. Lynch, J.D. & W. Duellman. 1980. The Eleutherodactylus of the Amazonian slopes of the Ecuadorian Andes (Anura, Leptodactylidae). The University of Kansas, Lawrence, Kansas 69: 1-86. [ Links ]
43. Lynch, J.D. & W. Duellman. 1997. Frogs of the genus Eleutherodactylus in Western Ecuador. Special publication, No. 23, Natural History Museum, The University of Kansas, Lawrence, Kansas. [ Links ]
44. Lynch, J.D. & B. Myers. 1983. Frogs of the fitzingeri group of Eleutherodactylus in Eastern Panama and Chocoan South America (Leptodactylidae). Bulletin of the American Museum of Natural History 175: 484-568. [ Links ]
45. Lynch, J.D. & A.M. Suárez. 2002. A new frog (Eleutherodactylus: Leptodactylidae) from the Southern part of the Cordillera Oriental de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 24: 289-293. [ Links ]
46. Nixon, K. & J.M. Carpenter. 1993. On outgroups. Cladistics 9: 413-426. [ Links ]
47. Padial, J. M., S. Castroviejo-Fisher, J. Köhler, C. Vilá, J. C. Chaparro & I. de la Riva. 2009. Deciphering the products of evolution at the species level: the need for an integrative taxonomy. Zoologica Scripta 38: 431-447. [ Links ]
48. Pyburn, W.F. & J.D. Lynch. 1981. Two little-known species of Eleutherodactylus (Amphibia, Leptodactylidae) from the Sierra de la Macarena, Colombia . Proceedings of the Biological Society of Washington 94: 404-412. [ Links ]
49. Raikow, R., A. Bledsoe, B. Myers & C. Welsh. 1990. Individual variation in avian muscles and its significance for the reconstruction of phylogeny. Systematic Zoology 39: 362-370. [ Links ]
50. Reichle, S., S. Lötters & I. de la Riva. 2001. A new species of the discoidalis group of Eleutherodactylus (Anura, Leptodactylidae) fron Inner-Andean Dry Valley of Bolivia. Journal of Herpetology 35: 21-26. [ Links ]
51. Rueda, J.V. & J.D. Lynch. 1983. Una nueva especie de Eleutherodactylus (Amphibia: Leptodactylidae) para la Cordillera Oriental de Colombia. Lozania 42: 1-6. [ Links ]
52. Salgar, L. 2003. Miología de manos y pies de Hyla labialis (Peters, 1883) y de Eleutherodactylus bogotensis (Peters, 1883). Trabajo de Grado, Departamento de Biología, Pontificia Universidad Javeriana Bogotá D.C. 258 pp. [ Links ]
53. Salgar, L., J.M. Hoyos & A. Acosta. 2009. New conditions and intraspecific variations in hand and foot muscles in Pristimantis bogotensis (Peters, 1863) (Amphibia:Anura: Strabomantidae). Zootaxa 2025: 56-68. [ Links ]
