










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.37 no.2 Bogotá July/Dec. 2015
https://doi.org/10.15446/caldasia.v37n2.54379
WILLIAM TRUJILLO-C.
RICARDO CALLEJAS POSADA
Maestría en Ciencias Biológicas, Facultad de Ciencias. Pontificia Universidad Javeriana. Bogotá, Colombia. williamtrujilloca@gmail.com
Instituto de Biología, Universidad de Antioquia, Apartado Aéreo 1226, Medellín, Colombia. callejas542004@yahoo.com
RESUMEN
Se describe e ilustra Piper andakiensis W. Trujillo & Callejas, sp. nov. (Piperaceae) procedente del piedemonte oriental de la cordillera Oriental en la jurisdicción de los departamentos de Caquetá y Putumayo (SE de Colombia). La especie nueva se distingue por sus hojas basalmente lobadas y oblicuas sobre un lado, con lobos divergentes y más cortos que el pecíolo, la lámina foliar gruesa y cartácea, con tallos, haz de la hoja, pecíolos y pedúnculos glabros y con crecimientos epidérmicos en pecíolos y márgenes peciolares; se discuten sus relaciones taxonómicas, se presentan comentarios sobre su distribución y ecología y se ilustran sus caracteres diagnósticos.
Palabras clave. Piper, piedemonte andino, Colombia
ABSTRACT
Piper andakiensis W. Trujillo & Callejas, sp. nov. (Piperaceae) is described and illustrated from the eastern foothills of the eastern cordillera in the the Colombian Andes, Departments of Caquetá and Putumayo (SE Colombia). The new species can be distinguished by the leaves lobed at the base and oblique, with divergent lobes that are shorter than the petioles, by the thick chartaceous leaf blades, with the stems, the upper surface of the leaf, petioles and peduncles glabrous, and by the presence of epidermic growths on petioles and petiolar margins, the taxonomic relationships of the species are discussed, comments on its distribution and ecology are presented and its diagnostic characters are illustrated.
Key words. Piper, new species, Andean foothills, Colombia.
Recibido: 20/04/2015
Aceptado: 03/11/2015
INTRODUCCIÓN
Piperaceae Giseke es una familia de distribución pantropical, con cinco géneros y cerca de 3500 especies (Wanke et al. 2006, 2007, Samain et al. 2008). En Colombia se registran los géneros Piper, Peperomia y Manekia. En Piper, tres amplios clados, cada uno con un componente geográfico particular, han sido definidos (Jaramillo & Manos 2001): Neotropical (1300 spp), Asia-Tropical (600 spp) y Pacífico-Sur (10 spp). En el análisis filogenético más exhautivo hasta ahora realizado para el género Piper, Jaramillo et al. (2008) descubrieron al interior de las especies neotropicales del género Piper (clado Neotropical), un clado que corresponde a lo que Miquel (1843-1844) denominó la sección Macrostachys del género Artanthe Miq. El botánico francés de ascendencia suiza, Frederich Anton Miquel publicó entre 1843 y 1844 su "Systema Piperacearum", una obra que para entonces se convertía en la revisión taxonómica más completa realizada para la familia Piperaceae a nivel global. En su obra, Miquel reconoce en la familia Piperaceae dos grandes subfamilias: Piperae (especies cuyos pistilos poseen tres estigmas) y Peperomiae (especies cuyos pistilos poseen un solo estigma); estas dos subfamilias se corresponden casi exactamente a lo que se reconoce hoy como los géneros Piper y Peperomia. Al interior de Piperae, Miquel describió 25 géneros de los cuales el género Artanthe, acomoda una porción significativa de las especies noetropicales de Piperae y que incluyen todas aquellas especies que poseen frutos con estigmas sésiles. Al interior de Artanthe, Miquel describió diez secciones, una de las cuales, la sección Macrostachys contiene especies con hojas pinnatinervias y donde las nervaduras secundarias divergen a lo largo de la nevadura central entre la base y el tercio proximal, el punto medio o el tercio distal de la lámina, con lobos basales a menudo notables en su desarrollo, con pecíolos vaginados en toda su extensión en todos los nudos, con espigas péndulas, largas y compactas y con flores cuatro estaminadas. Desde la descripción original de Macrostachys, la diagnosis del grupo no ha cambiado ostensiblemnte; podría adicionarse sin embargo, que las especies de Macrostachys son generalmente arbustos o árboles de tamaño mediano, umbrófilos y con una reducción del prófilo al estar cubierto total o parcialmente por el pecíolo (Callejas 1997); asi mismo, en algunas especies de Macrostachys es frecuente la presencia de tallos (ramas) huecos ocupados por hormigas del género Pheidole (Letourneau 2004, Tepe et al. 2007) o especies con cuerpos fructíferos presentes en el pecíolo, asociadas a especies del mismo linaje de hormigas. Miquel (1843'1844) incluyó en su sección Macrostachys a 21 especies, un número que se ha incrementado ostensiblemente desde entonces, estimándose al presente la existencia de 250–300 especies al interior de clado (Jaramillo et al. 2008, Callejas 2014).
En este artículo se describe e ilustra a Piper andakiensis, una especie nueva perteneciente al clado Macrostachys, que crece en bosques subandinos de Caquetá y Putumayo en Colombia sobre la vertiente oriental de la cordillera Oriental. El área de distribución de P. andakiensis fue explorada intensivamente por el primer autor durante los años 2007–2011, permitiendo un inventario preliminar del género en la transición andino-amazónica del Caquetá (Trujillo 2013), del cual resultaron varias especies nuevas para la ciencia.
Piper andakiensis W. Trujillo & Callejas, sp. nov.
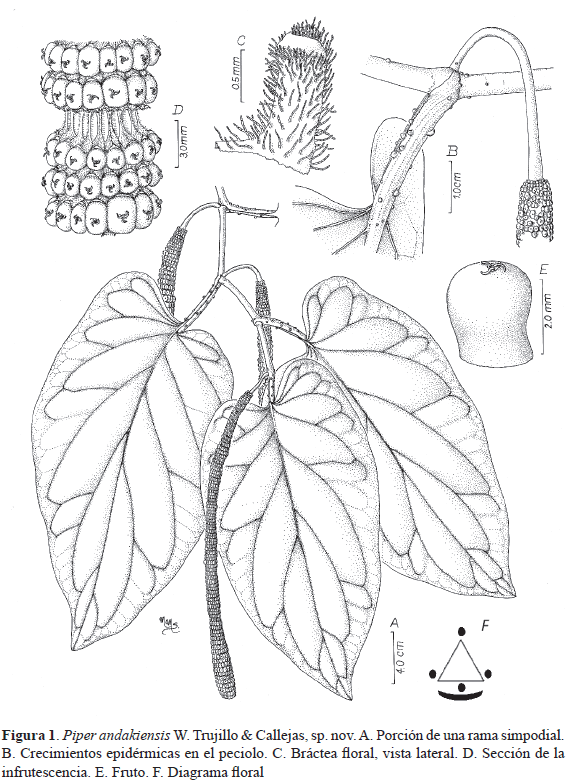
TIPO: COLOMBIA. Caquetá: Belén de los Andaquíes, Parque Bosque Microcuenca La Resaca, 1° 26' 23.5'' N, 75° 53' 24'' O, 482-720 m, 11 oct 2007, (fl, fr), W. Trujillo 1077 (holotipo HUA, isotipos COL, COAH, HUAZ). (Fig. 1 y Fig. 2)

Diagnosis
Treelets, 3–9 (12) m tall, with stems, petioles and peduncles glabrous, leaves 25–30 x 13–16 cm, truncate to very oblique at the base, with the lobes shorter than the petioles, divergent and leaving a lateral sinus, with farinose texture in fresh leaves similar to velvet, very thick chartaceous, with green to reddish petioles, with prominent epidermal projections, spikes pendent, 20–30 cm long, with the rachis fimbriate, and with fruits 2.5 x 3 mm, obpyramidal, glabrous and free from the rachis. Piper andakiensis exhibits a leaf shape, leaf base and number of veins also found in three species within the clade Macrostachys; P. auritifolium Trel. from Mesoamérica, a species that lacks the epidermal projections along the petiole and has membranaceous leaves which are densely pubescent on both sides; P. pallididorsum Trel. from the eastern slopes of the Peruvian Andes is quite similar to P. andakiensis but in P. pallididorsum the largest most basal lobe is paralel to the petiole and connivent, and the petioles and petiolar margins lack any epidermal growth and P. pennellii Trel. & Yunck.from the western slopes of the western cordillera in NW Colombia with juvenile leaves notably similar to the adult foliage of P. andakiensis but easily distinguished from the new species by the presence of epidermal growths in the stems, petioles, peduncles and leaf veins.
Descripción
Árbol umbrófilo de 3–9(12) m de alto, laxamente ramificado, entrenudos lisos, 8–17 cm de largo, 0.4–0.8 cm de diámetro, glabros, no punteados, de color verde cuando vivos y pardos cuando secos; prófilo no desarrollado. Hojas (23–)25–30 x (10–)13–16 cm, ovadas, medialmente simétricas, en la base equiláteras en los nudos del tallo principal o los nudos monopodiales y cordadas y lobadas, con los lobos redondeados a obtusos, mucho más cortos que el pecíolo, divergentes y dejando un seno central, o en la base inequiláteras en los nudos de los simpodios, lobadas y oblicuas sobre un lado y con un lobo 5–8 mm más largo que el otro, el lobo más largo obtuso y distante del pecíolo, el más corto truncado, mucho más cortos que el pecíolo, o en las hojas de simpodios más distales con la base casi equilátera y con las base truncada en ambos lado, en el ápice acuminadas, glabras, no punteadas, muy grueso cartáceas, ocasionalmente con textura farinosa cuando vivas, similar al terciopelo, primordios foliares de color rojizo en el envés cuando vivas, lámina secando negro–café a gris en la haz, pinnatinervias hasta la mitad de la longitud de la lámina, broquidódromo festonadas, con 4‒5 pares de nervaduras secundarias, las dos o tres nervaduras secundarias más proximales divergiendo entre la base y el tercio proximal, una cuarta o quinta nervadura secundaria divergiendo del punto medio, nervios terciarios exmedialmente ramificados, con ramificación dicotómica; nervios de cuarto orden no visibles. Pecíolos 3–5(6) cm de largo (los más largos en nudos estériles), estriados, glabros, no punteados, con crecimientos epidérmicos (Fig. 2b) a todo lo largo de su superficie, ocasionalmente de color rojizos cuando vivos, con la margen peciolar caduca a lo largo de toda su longitud ó hasta los 4/5 de su longitud (Fig. 2c), glabra, no punteada y con crecimientos epidérmicos. Espigas simples, terminales, de 20–30 cm de largo, 0.6–1 cm de diámetro, erectas en la antesis, eventualmente curvadas distalmente y péndulas en fruto, blancas en la antesis, verdes en fruto, las flores densamente agrupadas en todos los estadíos, formando bandas conspicuas alrededor del raquis; el raquis fimbriado, no punteado. Pedúnculo estriado, de 4.5–5 cm de largo, glabro, no punteado, verde. Brácteas florales 0.4–0.8 mm de diametro, cuculadas en la antesis, triangulares y deltoides en fruto, marginalmente cilioladas o fimbriadas, no punteadas, subsésiles, mucho más cortas que los frutos. Flores sésiles; 4 estaminadas; las anteras basifijas, 0.25 x 0,12 mm, con dehiscencia longitudinal en un plano oblicuo, el conectivo no prolongándose por encima de las anteras, los filamentos casi dos veces más largos que las anteras, filiformes; estilo ausente; estigmas 3, sésiles, laminares. Bayas de 2‒2.5(3) mm de largo x 2.5 mm de ancho, obpiramidales, apicalmente obtusas, lisas, glabras, no punteadas, negras cuando secas, libres del raquis.
PARATIPOS: COLOMBIA, Caquetá: Mun. El Paujil, vereda La Sonora, 1° 35' 86'' N, 75° 22' 43'' O, 1173 m, 1 oct 2005 (fl), M. Correa et al. 4980 (HUAZ). Mun. Florencia, vereda Las Brisas, kilómetro 26 vía Florencia-Suaza, 1° 42' 68'' N, 75° 42' 86'' O, 1350 m, 18 may 2009 (fl), D.C. Guevara et. al. 33 (HUAZ). Mun. Florencia, vereda El Caraño, 1° 44' 14'' N, 75° 40' 35'' O, 1100 m, 29 abr 2012 (fl), W. Trujillo & O. Perdomo 2069 (HUAZ). Mun. Florencia, vereda Las Brisas, vía Florencia–Suaza, 1392 m, 1° 44' 33'' N, 75° 40' 38'' O, 25 jun 2014 (fl), W. Trujillo et al. 3148 (HUA). Mun. Florencia, vereda El Caraño, finca Las Brisas, 1° 44' 14'' N, 75° 40' 36'' O, 1270 m, 23 jun 2014 (fl), W Trujillo et al. 3011 (HUA). Mun. Florencia, vereda Tarqui, cerca al monumento Divino Niño, vía antigua Florencia – Neiva, 1° 50' 03'' N, 75° 39' 55'' O,1570 m, 10 jul 2014 (fr), W. Trujillo et al. 3146 (HUA, COL, COAH). Mun. Florencia, corregimiento El Caraño, Vereda Quindío, 1° 42' 18'' N, 75° 38' 33'' O, 15 abr 2014, W. Trujillo et al. 3145 (COL). Putumayo: Mun. Mocoa, vereda Alto Campucana, corregimiento San Antonio, finca La Mariposa, 1° 12' N, 76° 38' O, 1350 m, 20 abr 1994 (fl), P. Franco et al. 5481 (COAH). Mun. Mocoa, Centro Experimental Amazónico (CEA), 1° 4'42'' N, 76° 37' 54'' O, 600 m, 29 ene 2012 (st), W. Trujillo & L. Rivera 2036 (HUAZ). Centro Experimental Amazónico (CEA), Jardin Botánico de Plantas Medicinales, 1° 4' 40'' N, 76° 37' 45'' O, 550 m, L. Rivera & Manuel Mueses 361 (COAH).
Etimología
El nombre de la especie hace referencia a la zona que comprende una parte de su distribución geográfica en el municipio de Belén de los Andaquíes, departamento del Caquetá, antiguo territorio ancestral de la etnia indígena Andakí.
Distribución y ecología
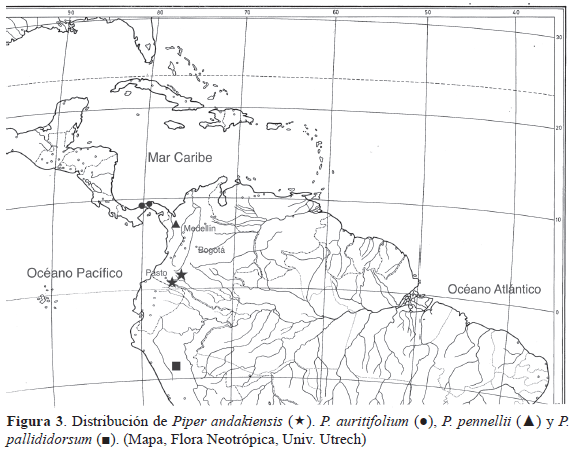
Piper andakiensis prospera en el sotobosque, sobre pendientes moderadas a muy pronunciadas en la vertiente oriental de la cordillera Oriental de Colombia, en el área de transición andino–amazónica de los departamentos de Caquetá y Putumayo entre 480–1500 m. Su distribución se circunscribe al distrito de Florencia, de la provincia biogeográfica de la Amazonia, de acuerdo con los unidades biogeográficas definidas por Hernández-Camacho et al. (1992). La localidad tipo, en el municipio de Belén de los Andaquíes, Caquetá, es un área declarada como área de reserva del orden municipal, sin embargo, presenta señales evidentes de intervención en el pasado reciente.
Fenología
Se observan periodos de floración en abril, junio, octubre y noviembre, y fructificación en julio y noviembre.
Comentarios
Piper andakiensis exhibe claramente los atributos de una especie perteneciente al clado Macrostachys, en paticular por los pecíolos vaginados en toda su extensión en todos los nudos, las márgenes peciolares que se extienden a lo largo de todo el pecíolo, las hojas basalmente lobadas, a menudo muy oblicuas sobre un lado, las inflorescencias de apreciable tamaño (20–30 cm de largo), las flores 4 estaminadas y los frutos con tres estigmas; todos estos atributos consituyen una combinación de caracteres mediante la cual se puede reconocer especies del clado. En adición a lo anterior en P. andakiensis son notorias: la presencia de hojas juveniles rojas en el envés, los pecíolos rojos en estadios juveniles y la presencia de crecimientos epidérmicos sobre el pecíolo y las margenes peciolares. No existe, de acuerdo a nuestro muestreo, ninguna otra especie descrita en el género Piper que pueda ser referida al clado Macrostachys y que exhiba la combinación de caracteres arriba indicada, sin embargo, lo anterior no necesariamente revela la posición filogenetica de esta nueva especie al interior de Macrostachys.
Los análisis filogenéticos para el género Piper son aún incompletos, y el número de terminales examinados en clados como Macrostachys igualmente muy pobre. En el análisis filogenético de Jaramillo et al. (2008), Macrostachys contiene escasamente 40 terminales, de los cuales solo nueve prosperan en Sur América. En Colombia se ha detectado la presencia de casi 90 especies pertenecientes a clado Macrostachys pero de estas solo cinco han sido exploradas en análisis filogeneticos (Jaramillo et al. 2008); el muestreo es entonces aún muy exiguo e incompleto y en consecuencia no es fácil establecer con alguna certeza las relaciones filogenéticas de P. andakiensis; asi pues, no podemos sino discutir las similitudes que esta nueva especie exhibe con otros taxones en el clado Macrostachys.
P. andakiensis prospera en un área donde es simpátrica con P. mocoanum Trel. & Yunck., una especie del clado Macrostachys pero que exhibe hojas pubescentes en el envés, pecíolos pubescentes y que no posee crecimientos epidermicos en el pecíolo o en las márgenes peciolares. Un examen de las especies de Macrostachys revela sin embargo al menos tres especies en este clado que exhiben notorias similitudes con P. andakiensis: P. auritifolium de Mesoamérica, P. pennellii del piedemonte occidental de la cordillera Occidental en el sur de Colombia y P. pallididorsum del piedemonte oriental andino en Perú (Fig. 3). Piper pennellii posee hojas juveniles que son prácticamente idénticas a las hojas adultas de P. andakiensis, pero en P. pennellii las hojas exhiben crecimientos epidérmicos en tallos, pecíolos, pedúnculos y nervaduras en el envés de las hojas; P. auritifolium de las tierras bajas de la costa Caribe en Panamá y Costa Rica es muy similar tanto en caracteres vegetativos (hojas oblicuas en la base con lobos más cortos que el pecíolo), como en caracteres florales, (brácteas florales triangulares y ciliadas marginalmente), pero las hojas son pubescentes en ambas caras y los pecíolos y margen peciolar no poseen crecimientos epidérmicos, y P. pallididorsum del Perú que posee hojas con la forma, tamaño y nervación similares a P. andakiensis pero el lobo más largo es paralelo y connivente al pecíolo y no presenta crecimientos epidérmicos. El contraste entre esas tres especies y P. andakiensis se consigna en la siguiente clave:
1. Tallos, pecíolos y pedúnculos pubescentes.
2. Ramas jóvenes, pecíolos, pedúnculos y nervaduras en el envés de las hojas
con crecimientos epidérmicos, hojas ampliamente ovadas, 40–50 cm de largo, 20–30 cm de
ancho ........................................P. pennelli
2. Ramas jóvenes, pecíolos, pedúnculos y nervaduras en el envés de las hojas sin crecimientos
epidérmicos, hojas ovadas o elíptico-ovadas, 18–23 cm de largo, 12–14 cm de ancho ...................................P. auritifolium
1. Tallos, pecíolos y pedúnculos glabros.
3. Pecíolos y márgenes peciolares con crecimientos epidérmicos, el lobo más largo divergente, no paralelo o connivente al pecíolo ...........................P. andakiensis
3. Pecíolos y márgenes peciolares sin crecimientos epidérmicos, el lobo más largo paralelo y casi
connivente con el pecíolo..P. pallididorsum
De acuerdo con las características de campo, Piper andakiensis se diferencia de otras especies de Macrostachys que crecen en su área de distribución, por las hojas fuertemente coriáceas, gruesas, verde oscuras en la haz y de textura farinosa (semejante al terciopelo); crece hasta 12 m y alcanza 10 cm de diámetro en el tallo, siendo probablemente una de las especies del género con mayor talla en el piedemonte andino del Caquetá, con individuos reproductivos desde los 3 m de altura y pecíolos rojizos (Fig. 2c) en la mayoría de individuos que crecen entre los 1000 y 1500 m de altitud.
Cabe señalar que los crecimientos epidérmicos encontrados en la superficies caulinares o foliares en especies del género Piper se restringuen solo a algunas especies del clado Macrostachys y de su grupo hermano Hymenopodium (constituido por c. 20 especies que incluye entre otras a especies de amplia distribución como P. tuberculatum Jacq.) y exclusivamente a especies que prosperan en la zona andina de Colombia, Venzuela, norte de Ecuador y las planices costeras del Caribe en Panamá y Costa Rica. Analisis filogeneticos sugieren que especies con crecimientos epidemicos en Macrostachys no conforman un grupo monofilético y que han surgido numerosoas veces de forma independiente en la evolución del grupo.
AGRADECIMIENTOS
El primer autor agradece a la Universidad de la Amazonía por el apoyo durante las salidas de campo que permitieron las primeras recolecciones de Piper en el Caquetá, al Herbario Amazónico Colombiano del Instituto Sinchi por facilitar la consulta de especimenes y el estudio de piperáceas de la Amazonía colombiana durante los años 2010 y 2011 y a la Pontificia Universidad Javeriana donde realizamos parte de la descripción y la ilustración de la especie. A Erasmo Gonzales y a la Fundación Tierra Viva por su asistencia en campo y por proveer apoyo logístico para el ingreso a varias localidades de Belén de los Andaquíes. A tres evaluadores anónimos que ayudaron a mejorar el manuscrito. Ricardo Callejas agradece al Laboratorio de Biotecnología de la Sede de Investigaciones Universitarias (SIU) por su continuo apoyo en el estudio de las piperáceas neotropicales, y a los curadores y personal administrativo del Herbario de la Universidad de Antioquia (HUA).
LITERATURA CITADA
1. Callejas, R. 2014. Piperaceae In: B.E. Hammel, M. H. Grayum, C. Herrera & N. Zamora (eds.) Manual de Plantas de Costa Rica. Vol. VII. Pp-6-326. Missouri Botanical Garden-INBIO [ Links ]
2. Jaramillo, M. A., R. Callejas, C. Davidson, J. F. Smith, A. C. Stevens & E. J. Tepe. 2008.A phylogeny of the tropical genus Piper using ITS and the Chloroplast Intron psbJ–petA. Systematic Botany 33: 647–660. [ Links ]
3. Jaramillo, M.A. & P.S. Manos. 2001. Phylogeny and patterns of floral diversity in the genus Piper (Piperaceae). American Journal of Botany 4: 706–716. [ Links ]
4. Letourneau, D. K. 2004. Mutualism, antiherbivore defense, and trophic cascades: Piper ant-plants as a mesocosm for experimentation in: L. A. Dyer & A. D. Palmer (eds), Piper: A model genus for studies of phytochemistry, ecology and evolution, Kluwer Academic Publishers, Nueva York. [ Links ]
5. Miquel, F. A. G. 1843–1844. Systema Piperacearum. Kramer, Rotterdam. [ Links ]
6. Samain, M. S., G. Mathieu, S. Wanke, C. Neinhuis & P. Goetghebeur. 2008. Verhuellia revisitedunravelling its intricate taxonomic history and a new subfamilial classification of Piperaceae. Taxon 57: 583–587. [ Links ]
7. Tepe, E. J., M.A. Vincent & L.E. Watson. 2007. Stem diversity, cauline domatia, and the evolution of ant–plant associations in Piper sect. Macrostachys (Piperaceae). American Journal of Botany 94: 1–11. [ Links ]
8. Trujillo-C., W. 2013. El género Piper L. (Piperaceae) en el Alto Caquetá, zona de transición andino amazónica de Colombia. Revista Colombia Amazónica 6: 111-121. [ Links ]
9. Wanke, S., M. Samain, l. Vanderschaeve, G. Mathieu, P. Goetghebeur & C. Neinhuis. 2006. Phylogeny of the genus Peperomia (Piperaceae) inferred from the trnK/matK region (cpDNA). Plant Biology 8: 93–102. [ Links ]
10. Wanke, S., M. Jaramillo, T. Borsch, M. Samain, D. Quandt & C. Neinhuis. 2007. Evolution of Piperales-matK gene and trnK intron sequence data reveal lineage specific resolution contrast. Molecular Phylogenetics and Evolution 42: 477–497. [ Links ]
