










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.37 no.2 Bogotá July/Dec. 2015
https://doi.org/10.15446/caldasia.v37n2.53870
ANA MARÍA BENAVIDES
ALBA LORENA HERNÁNDEZ-G.
Corporación Para Investigaciones Biológicas - Unidad de Biodiversidad. Cra. 72A No. 78B-141, Medellín. Colombia. abenavides@cib.org.co
RESUMEN
En los Andes colombianos, un gran número de artesanías se fabrican a partir de raíces adventicias de plantas nativas que crecen de forma silvestre. Sin embargo, la deforestación y las prácticas no sustentables han diezmado la disponibilidad de plantas silvestres productoras de fibra. En este estudio evaluamos la disponibilidad de fibras de tres especies en Santa Elena (Antioquia, Colombia): Asplundia sarmentosa, Philodendron aff. sagittifolium y Clusia multiflora. Después de una búsqueda exhaustiva en la zona de estudio ubicamos menos de 30 individuos adultos de cada especie. Para cada planta se midieron las variables asociadas al tamaño de la planta y se registró el número y la longitud de raíces maduras e inmaduras. Se pesaron fragmentos de raíces para estimar la biomasa. El crecimiento de raíces de cada planta fue monitoreado mensualmente durante siete meses. La disponibilidad actual de fibra (raíces maduras) es de ca. 3 352 m (Asplundia 1 481 m, Clusia de 952 m, Philodendron 918 m), con esta cantidad de material se pueden elaborar entre 419 a 838 canastos. De acuerdo a la tasa de crecimiento mensual, una nueva raíz podría alcanzar un tamaño para ser cosechado entre el mes 38 y 71. Plantas más grandes tienen mayor cantidad de raíces maduras, sin embargo, la tasa de crecimiento no está asociada al tamaño de la planta. Nuestros resultados proveen evidencia de que las plantas en Santa Elena presentan una relación (tamaño de la planta, número de raíces maduras) que es similar a plantas que se encuentran en zonas no intervenidas, no obstante, la cosecha de fibras en Santa Elena no es una actividad sostenible debido al bajo número de individuos susceptibles de ser cosechados, la lenta tasa de crecimiento de raíces y la alta demanda de fibras. Es necesario implementar estrategias como el enriquecimiento del bosque con estas tres especies para garantizar una actividad sustentable para recolectores y artesanos en Santa Elena y en otros bosques de la cordillera cordillera Central de los Andes.
Palabras clave. Asplundia, Clusia, crecimiento de raíz, Fibras, Philodendron.
ABSTRACT
In the Colombian Andes many artisanal handicrafts are made from aerial roots of wild native plant species. However deforestation and unsustainable practices have reduced the availability of wild plants used as fiber source. In this study, we assessed the availability of fibers of three species in Santa Elena (Antioquia): Asplundia sarmentosa, Philodendron aff. sagittifolium and Clusia multiflora. After an exhaustive searched in the study area were located less than 30 adult plants of each species. For each plant we measured plant size variables and recorded the number and length of mature and immature roots. Fragments of roots were weighted to estimate biomass. Monthly root growth measures were done during seven months. The current availability of fiber (mature roots) is ca. 3 352 m (Asplundia 1 481 m, Clusia 952 m, Philodendron 918 m), and between 419 and 838 baskets can be made with this amount of material. According to the monthly root growth rates, a new aerial root could reach a harvestable size after 38 to 71 months. More mature roots were found in larger plants; however, growth rate is not associated with plant size. Our results provide evidence that plants in Santa Elena have a similar relationship in size and number of mature roots as those from undisturbed areas. However, the fiber harvest in Santa Elena is not sustainable due to the low amount of plants likely to be harvested, the slow root growth rate and the high demand for fiber. In fact, it is necessary to implement strategies such as forest species enrichment with these three species to ensure a sustainable activity for artisans in the Santa Elena and in other forests in the Central Cordillera of the Andes.
Key words. Asplundia, Clusia, fiber, Philodendron, root
Recibido: 28/03/2015
Aceptado: 22/10/2015
INTRODUCCIÓN
Los productos forestales no maderables (PFNM), que abarcan un amplio espectro de productos derivados de plantas y animales (Wickenes 1991), son para las comunidades rurales pobres una de las fuentes de generación de dinero (Ticktin 2004, Kusters et al. 2006). Los PFNM son en su gran mayoría obtenidos de la cosecha directa en ecosistemas naturales y usualmente provienen de fuentes limitadas (Linares et al. 2008, Ticktin 2004). En la región Andina se destaca el uso de fibras vegetales para la elaboración de cestería y artesanías en general (Macia 2006, Linares et al. 2008, Feuillet et al. 2011).
Las fibras se obtienen de la extracción de raíces, cortezas, pecíolos y/o tallos (Macia 2006, Linares et al. 2008). En Colombia, 85 de las 114 especies de plantas empleadas para obtener fibras son cosechadas de directamente del bosque (Linares et al. 2008). Recientes estudios muestran que la cosecha directamente en el bosque de PFNM produce una disminución en las poblaciones silvestres como consecuencia de la sobreexplotación (Ticktin, 2004, García & Galeano 2009a, García et al. 2013). Particularmente en la región Andina colombiana, esta explotación intensiva, sumada a los fuertes cambios en el paisaje (Eteer & van Wyngaarden 2000, Armenteras et al. 2003) ha generado un desabastecido general de materia prima para la elaboración de artesanías (Ramos 2000, Linares et al. 2008).
El oriente antioqueño, en la Cordillera Central de los Andes en Colombia, es una de las zonas con mayor actividad artesanal dentro de la región, especialmente los municipios aledaños a la ciudad de Medellín como el corregimiento de Santa Elena (Linares et al. 2008). En Santa Elena, conjuntos familiares campesinos se reúnen para elaborar cestos y canastas a partir de fibras de las raíces adventicias de plantas nativas de los géneros Clusia, Asplundia y Philodendron, que son luego comercializadas en los mercados locales de la ciudad (Gómez & Orozco 2001, Holos-Natura 2001, Londoño 2004). Generalmente, la fibra es suministrada a las familias artesanas por recolectores locales que usualmente extraen las raíces en bosques aledaños en épocas previas a la navidad o de acuerdo a la demanda del producto por comerciantes locales (Gómez & Orozco 2001).
El conocimiento de los factores que determinan la producción de fibras (por ejemplo: edad o tamaño de la planta) podría brindar elementos para generar los programas de manejo de estas especies (Plowden et al. 2003, Endress et al. 2004, Ticktin 2004). Así, el manejo sustentable de los PFNM podría ofrecer alternativas económicas para familias rurales y además podría ser una estrategia de conservación de los bosques donde se cosechan las plantas (Belcher 2005, Kusters et al. 2006). En Colombia, con excepción de especies de palmas (Galeano et al. 2010 Bernal et al. 2011, Navarro-López et al. 2014), solo para unas pocas especies de plantas nativas productoras de fibras han sido estudiados aspectos de su ecología que permitan desarrollar estrategias de manejo (Balcázar et al. 2015, García & Galeano 2009 ab, García et al. 2013).
En este estudio examinamos la disponibilidad de raíces adventicias de tres especies de los géneros Clusia, Philodendron y Asplundia, especies tradicionalmente cosechadas para la elaboración de artesanías en Santa Elena, Antioquia (Colombia). Nuestra hipótesis es que de acuerdo a la relación alométrica esperada entre el tamaño de las plantas y la tasa de crecimiento (Niklas et al. 2001), plantas más grandes pueden pasar más recursos a las raíces, que se vería reflejado en mayor biomasa, por lo tanto la cantidad de raíces y la tasa de crecimiento incrementarán con el tamaño de las plantas. Así mismo esperaríamos, de acuerdo al espectro económico de las construcción de las hojas (Royer 2008, Wright et al. 2004), que especies en las que la inversión en la construcción de sus raíces es menor (por ejemplo: menor peso seco por centímetro), crecerán a mayor velocidad, siendo por esto más factible su cosecha. Buscamos responder: (1) ¿Cuántas plantas con raíces factibles de cosechar existen actualmente en Santa Elena? (2.) ¿Cuántas raíces ancladas (factibles de cosechar) y sin anclar (en proceso de crecimiento) tiene en promedio cada planta? (3) ¿Cuál es la tasa de crecimiento de las raíces? (4.) ¿Existe una relación entre el tamaño de las plantas y la tasa de crecimiento y/o la cantidad de raíces que forma? (5.) ¿Existe una relación entre la biomasa de raíces de las especies con la tasa de crecimiento y/o la cantidad de raíces que forma? El propósito de este estudio es generar información que permita evaluar la sostenibilidad de las especies productoras de fibra en Santa Elena (Antioquia) y establecer lineamientos para su conservación.
MATERIALES Y MÉTODOS
Este estudio se realizó al norte de la Cordillera Central de los Andes de Colombia en el corregimiento de Santa Elena, Medellín (6°13' Norte, 75°34' Oeste). El área de estudio abarcó ca. de 6.4 km2 de extensión y se ubica entre los 2 200 y 2 600 msnm. El área de estudio incluyó el Parque EcoturísticoArví, las reservas El Robledal y La Aguada. Esta área corresponde a fragmentos de bosques andino en diferentes estados de conservación, en un mosaico de plantaciones de pino (Pinus patula Schldl. et Cham.), ciprés (Cupressu lusitánica Mill.), también es frecuente ver matrices de pastos, áreas de vivienda recreativa y en menor proporción cultivos (Duque et al. 2007). El bosque está compuesto principalmente por robledales (Quercus humboltii Bonpl.), además de otras especies de los géneros Clusia sp., Alfaroa sp., Drymis sp., Tibouchina sp., Palicurea sp., entre otras. El régimen de lluvias es bimodal con una precipitación promedio anual de 1 890 mm, los periodos lluviosos corresponden a los meses de marzo-abril-mayo y septiembre-octubre-noviembre. La temperatura promedio anual es de 14.4 °C y presenta una variación diaria entre 5° y 22°C (Duque et al. 2007, SIATA 2011). En el 2001, entre 7-11 recolectores ejercían la actividad de extracción de fibras con una frecuencia semanal y/o quincenal y se elaboraban las artesanías en seis talleres familiares con 21 artesanos, predominantemente ubicados en la vereda Mazo en el Sector Los Vásquez (Gómez & Orozco 2001).
Descripción de las especies
Se estudiaron tres especies que desarrollan largas raíces adventicias y son cosechadas en la zona de estudio. (1) Asplundia sarmentosa Galeano & R. Bernal. (Cyclanthaceae), es una liana nómada (sensu Zotz 2013) que alcanza hasta diez metros de longitud, ocasionalmente es rastreras. Localmente es conocida como cestillo o bejuco liso (Gómez & Orozco 2001) y presenta una amplia distribución en la cordillera central de Colombia sobre los 2 000 msnm (Galeano-Garcés & Bernal-González 1984, Tubergia com. pers.). (2) Clusia multiflora Kunth. (Clusiaceae) es un árbol que puede alcanzar los 20 metros de altura. Localmente esta planta es conocida como chagualo (Gómez & Orozco 2001). Se distribuye en Centroamérica desde Honduras a Panamá, y al norte de Suramérica desde Colombia a Bolivia especialmente sobre la Cordillera de los Andes (MOBOT 2014). En Colombia está presente en la Sierra Nevada de Santa Marta y en las tres cordilleras, entre 1 000 y 3 300 msnm. (3) Philodendron aff. sagittifolium Liebm. (Araceae) es una liana nómada que puede alcanzar hasta los doce metros de altura sobre el árbol hospedero. En la zona de estudio se conoce como trepadora o guaco (Gómez & Orozco 2001). Se distribuye en Mesoamérica, Centroamérica y el norte de Suramérica (MOBOT 2014). En Colombia se encuentra desde el nivel del mar hasta los 2 400 msnm. Se considera una especie morfológicamente variable así como muy adaptable (Croat 1997). En adelante en el texto mencionamos las especies de acuerdo a su género.
Selección de plantas y monitoreo de crecimiento de raíces
El trabajo de campo se realizó entre mayo de 2013 y febrero de 2014. Para responder ¿Cuántas plantas son "cosechadas actualmente en Santa Elena? Principalmente en la vereda Mazo del corregimiento de Santa Elena consultamos ocho personas involucradas en la elaboración de artesanías con fibras, dos de ellas colectores que fueron reconocidos como los únicos que están realizando la actividad actualmente. La consulta la realizamos por medio de entrevista semi-estructuradas (Dean Hammer & Aaron Wildavsky 1990) que incluía preguntas sobre la práctica de cosecha, uso de las fibras y la ubicación de las plantas que se cosechan actualmente. En el área de estudio, adicionalmente realizamos en compañía de los artesanos y recolectores recorridos en los fragmentos de bosques remanentes en la zona y áreas aledañas (con cobertura de pastos o plantaciones forestales) con el fin de identificar la presencia de plantas de cada una de las especies que los colectores no tuviesen presente. Las colecciones botánicas de cada especie fueron depositadas en el herbario Universidad de Antioquia (HUA).
Para determinar el número de raíces por planta y estimar la tasa de crecimiento de raíces adventicias, procuramos en el área de estudio un mínimo de 30 individuos por especie, no obstante no fue posible conseguir el número mínimo para ninguna de las tres especies. Encontramos 29 individuos de Asplundia, 22 individuos de Clusia, 18 individuos de Philodendron. Para Asplundia y Philodendron definimos el individuo como una planta con uno o varios tallos sobre un árbol hospedero, debido a la imposibilidad de diferenciar sí los tallos provienen de un solo individuo biológico (Zotz 2013).
Como nuestro objetivo fue cuantificar el recurso actual (número de plantas y raíces) disponible para ser aprovechado por recolectores, seleccionamos los individuos teniendo en cuenta los criterios que emplean los recolectores locales que son: plantas adultas con raíces adventicias en contacto con el suelo (raíces ancladas conocidas como "maduras"). Para facilitar su búsqueda mensual y calcular la densidad de las poblaciones, georreferenciamos cada individuo y registramos su ubicación en un plano cartesiano. Para responder cuántas raíces ancladas y no ancladas tiene en promedio cada planta, contamos el número de raíces adventicias (ancladas y sin anclar) en cada planta.
En cada planta seleccionada por cada diez raíces no ancladas seleccionamos cinco raíces para medir su longitud al inicio del estudio y monitorear su elongación mensual durante siete meses (210 días). En el caso de plantas con menos de diez raíces, seleccionamos la mitad del total de raíces. Para cada raíz registramos la longitud total y cada raíz fue marcada con cinta aislante de colores distintivos, a una distancia de diez centímetros desde la cofia. Para responder la cuarta pregunta la cual busca determinar si existe una relación entre la tasa de crecimiento, el número de raíces y el tamaño de la planta, medimos para cada individuo la longitud total de tallo, contamos el número de hojas para Philodendron y Asplundia y estimamos el área de la copa proyectada para los arboles de Clusia.
Finalmente para describir la biomasa de raíces de las especies y discutir si esta tiene que ver con la tasa de crecimiento y/o la cantidad de raíces que forma, estimamos la biomasa seca, fresca de raíces no ancladas y porcentaje de agua en el tejido. Primero seleccionamos entre una y dos raíces en quince plantas por cada especie. Basados en todo el material colectado mediante una discriminación aleatoria seleccionamos la posición de corte de 60 fragmentos de 10 cm de longitud por especie. Pesamos los fragmentos frescos y posteriormente se secaron en horno a 70°C hasta alcanza un peso constante (con modificaciones, Pérez-Harguindeguy et al. 2013).
Análisis numéricos
La tasa de crecimiento, el número de raíces ancladas y no ancladas, la longitud de las raíces y biomasa de raíces entre las tres especies fueron analizados mediante la prueba de Shapiro-Wilk para probar si presentaban una distribución normal. Consecuentemente se utilizó la prueba de Kruskal-Wallis (análisis de varianza no paramétrico) para probar las diferencias entre las tres especies para cada una de las variables (tasa de crecimiento, el número de raíces ancladas y no ancladas, la longitud de las raíces y biomasa de raíces). Para probar la relación entre el tamaño de la planta con el número de raíces y el crecimiento de raíces tanto mensual como total acumulado de cada especie se realizaron pruebas empleando el coeficiente de correlación de Pearson. Los análisis se realizaron con el paquete Stats en el programa R (R Development Core Team 2008).
RESULTADOS
De acuerdo a la entrevista, los recolectores y los artesanos en la zona de estudio cosechan las raíces adventicias que han entrado en contacto (raíces ancladas) con el suelo que denominan "raíces maduras". Las raíces que no han entrado en contacto con el suelo (no ancladas) se denominan "niñas o inmaduras". La raíz seleccionada para la cosecha se somete a varias torsiones que ocasionan el corte. Luego de un proceso de pelado manual, se separa el córtex, que es la parte empleada en la elaboración de la artesanía. Para la elaboración de un canastos mediano los artesanos emplean entre cuatro a ocho metros de fibras bien sea una combinación de raíces de las tres especies o una o dos de las especies.
Después de una búsqueda exhaustiva en la zona de estudio (ca. de 6.4 km2), que incluyo tanto las plantas señaladas como cosechadas por artesanos y colectores, ubicamos menos de 30 individuos adultos con raíces adventicias de cada una de las especies. Los individuos se encontraron en bosques nativos en estadios secundarios. Localizamos 29 individuos de Asplundia, 22 de Clusia y 18 de Philodendron. Los individuos de Clusia se encontraron en una área de 0.470 Km2 y la distancia promedio entre plantas fue de 87.5 m, mientras que plantas de Philodendron se encontraron en dos sitios que distan entre sí 8.1 km. En el primer sitio las plantas se encontraron en un área de 0.050 Km2 y presentaron una distancia promedio entre las plantas de 64.3 m y en el segundo sitio se encuentran en un área de 0.0003 km2 con una distancia promedio entre plantas de 24 m. Las plantas de Asplundia se encontraron en un área de 0.014 Km2 con una distancia promedio de doce metros entre las plantas.
La tasa de crecimiento, el número de raíces ancladas y no ancladas, la longitud de las raíces y biomasa de raíces entre las tres especies presentaron una distribución no normal (p<0.05). El número de raíces maduras, aquellas que están ancladas las cuales representan el recurso actual de fibras, es similar entre las tres especies (Figura 1, Tabla 1), la mitad de las plantas presentaron 10 raíces ancladas o más. La longitud de las raíces al entrar en contacto con el suelo, no es diferente entre las tres especies y varía entre 3.3 y 4.7 m (Tabla 1). Al extrapolar la longitud de las raíces (sin cuantificar la porción enterrada) al número de raíces ancladas en cada planta, encontramos que la longitud actual de raíces disponibles para aprovechar es en promedio por planta es de 51.1 m en Asplundia, 43.3 m en Clusia, 51 m en Philodendron. Así, basado en el número de plantas encontradas, la cantidad y el promedio de raíces ancladas por planta, la disponibilidad actual de raíces es de ca. 3 352 m (Asplundia 1 481 m, Clusia 952 m y Philodendron 918 m), por lo que la oferta de fibras actual (una sola vez) alcanzaría para la elaboración de 419 a
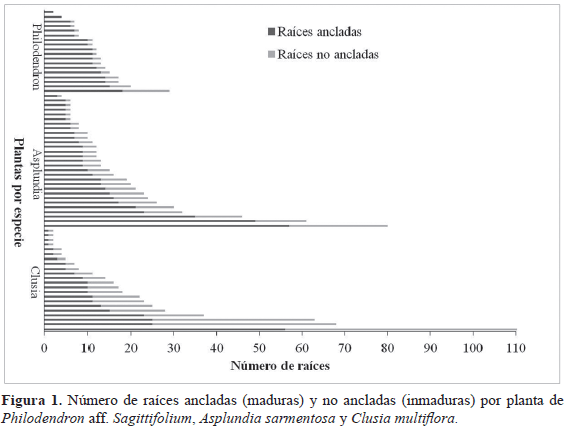
Las raíces inmaduras, aquellas que no estaban ancladas las cuales representan el recurso en desarrollo, son menos abundantes en Philodendron (la mitad de los individuos solo presentaba una raíz) que en Clusia y Asplundia en los cuales la mitad de los individuos presentaron entre cuatro y cinco raíces no ancladas (Figura 1, Tabla 1). La mediana de la longitud de las raíces adventicias no ancladas para Clusia y Asplundia es de 1.6 m y 1.7 m, respectivamente, mientras que en Philodendron alcanza 3.6 m de longitud (Tabla 1).
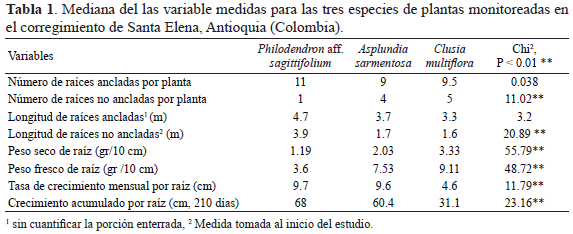
Para las tres especies existe una relación significativa entre el tamaño de la planta y el número de raíces ancladas (Figura 2). Para Clusia, tanto el tamaño del árbol como el área de la copa de los arboles presenta una relación positiva con el número de raíces ancladas y sin anclar (área de la copa: raíces ancladas, r=0.47, p=0.025; área de la copa: raíces sin anclar, r= 0.48, p=0.020; tamaño: raíces sin anclar, r=0.43, p=0.044). El número de hojas de Asplundia y Philodendron no presenta una relación con el número de raíces ancladas o raíces sin anclar (Pearson p>0.05).
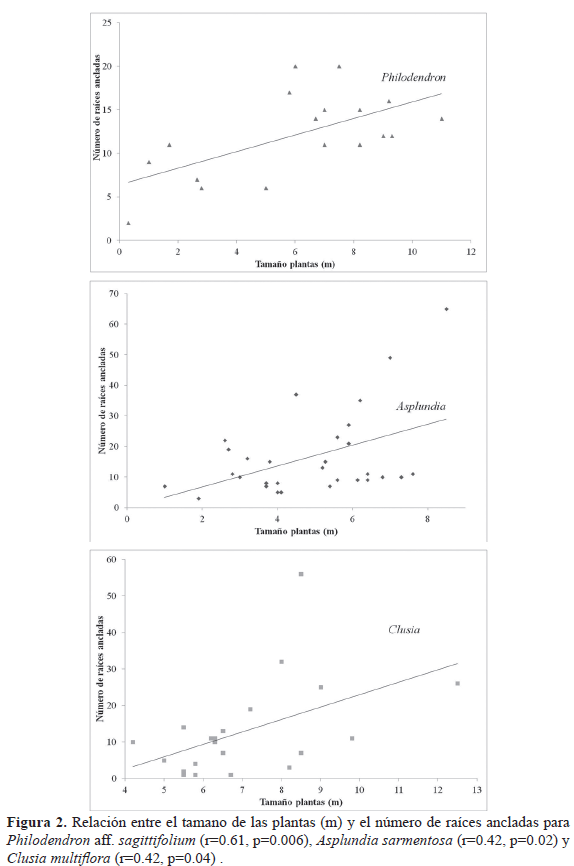
La tasa de crecimiento mensual por raíz es menor para Clusia y es significativamente diferente entre las especies (Tabla 1). El crecimiento en longitud después de 210 días en promedio por una raíz aérea por planta fue mayor en Philodendron, seguido de Asplundia y Clusia (Figura 3; Tabla 1). Asumiendo un crecimiento lineal de acuerdo a la tasa de crecimiento mensual y a la longitud promedio de las raíces que alcanzan el suelo, una nueva raíz adventicia de Asplundia alcanzaría el suelo luego de 38 meses, en Philodendron a los 48 meses y Clusia lo alcanzaría a los 71 meses.
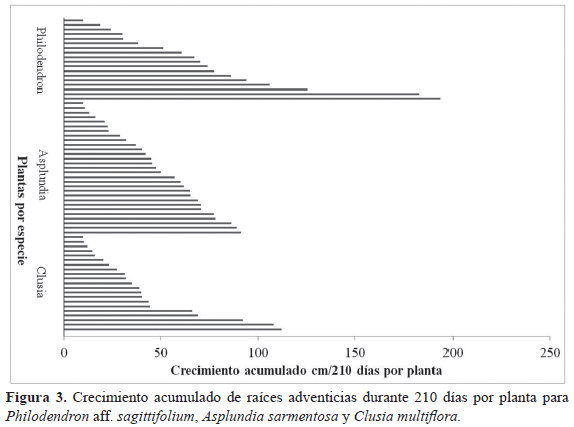
El promedio de crecimiento de raíces por planta para las tres especies no está relacionado con el número de raíces ancladas que presenta la planta (correlación de Pearson, p > 0.05). Además, el crecimiento de raíces tanto mensual como total acumulado de cada especie no está relacionado con ninguna de las variables de tamaño de las plantas (correlación de Pearson, p > 0.05).
Encontramos diferencias significativas en la biomasa seca y fresca de raíces en las tres especies. Los fragmentos de raíz de Asplundia y Philodendron presentan un menor peso seco, en relación a las raíces de Clusia (Tabla 1).
DISCUSION
La disponibilidad de raíces de ca. de 3 352 m (Asplundia 1 481 m, Clusia 952 m y Philodendron 918 m), es una oferta relativamente baja para la posible demanda de productos elaborados con estas fibras naturales en la zona. Al parecer de acuerdo Gómez & Orozco (2001), esta escasez de fibra podría ser relativamente reciente y ser una consecuencia directa de una sobrexplotación durante las última décadas, puesto que recolectores actuales recuerdan la extracción de toneladas de fibras de estas especies. Por ejemplo recolectores reportaron la extracción que realizaron durante dos años de cerca de 5 000 bultos que equivaldría a cerca de 86 toneladas de raíces de Philodendron (Gómez & Orozco 2001).
El bajo número de plantas productoras de fibras en Santa Elena como resultado de una posible consecuencia histórica de sobrexplotación se suma a una dramática perdida de bosque en la zona. Todos los individuos se encontraron en bosque nativo que en la zona de estudio se encuentra altamente intervenido, disperso y no sobrepasa el 7.6% de la cobertura (Holos-Natura 2001, Duque et al. 2007), indicando posiblemente que la fragmentación y pérdida dramática de la cobertura de los bosques en el área de estudio ha agudizado la escasez de plantas. Las plantas como Asplundia y Philodendron dependen de la presencia de árboles para su desarrollo (Benavides et al. 2013, Cockle 2001, Ray 1992), al perderse el bosque la posibilidad de conservación para estas especies disminuye. En el caso de Clusia, un árbol dioico, los procesos de polinización son altamente dependientes de la conectividad del bosque y la fragmentación actual podría estar afectando la producción de frutos (Liebig et al. 2001, de Faria et al. 2006).
Nuestra hipótesis sobre la esperada relación entre el tamaño de la planta con el número de raíces y su crecimiento, se cumple parcialmente. En este estudio encontramos que no hay una relación entre el tamaño de las plantas y el crecimiento de las raíces adventicias no ancladas. No obstante, encontramos una relación positiva entre el tamaño de la planta y el número de raíces ancladas. García y Galeano (2009b) reportaron un patrón similar en plantas de Philodendron longirrhizum que se encontraban en áreas no intervenidas. En el mismo estudio, plantas en áreas intervenidas no mostraron una relación positiva entre el número de raíces maduras y el tamaño de la planta. La relación encontrada entre el tamaño de las plantas y el número de raíces ancladas sugiere que las plantas en Santa Elena no están siendo fuertemente cosechadas y un patrón natural se conserva. Si bien Santa Elena es un área bastante intervenida, evidenciamos una disminución de artesanos activos en la zona de acuerdo a lo reportado por (Gómez & Orozco 2001) que podría estar permitiendo una recuperación de las plantas.
La falta de relación entre el crecimiento de la raíz y el tamaño de las plantas podría indicar que la elongación de la raíz de las tres especies estudiadas depende de factores externos a la planta. Diferentes estudios brindan evidencias sobre la mediación de la fitohormona auxina en la activación y crecimiento de raíces adventicias (Negi et al. 2010, Lanteri et al. 2006, Visser et al. 1996). Esta hormona responde a señales que se activan con estímulos como la gravedad y otros relacionados con el crecimiento de la planta, este último es regulado en respuesta a estímulos ambientales (Vanneste & Friml 2009). Por tanto, el crecimiento de raíces, en este caso adventicias, puede ser regulado por condiciones ambientales particulares a cada planta como disponibilidad de agua, luz y/o nutrientes (Poorter & Nagel 2000, Lynch 1995). La falta de relación entre el crecimiento y el número de hojas indica que la cantidad de hojas per se no es un indicador del tamaño de plantas nómadas.
Los resultados sobre la tasa de crecimiento de las raíces adventicias están en el rango bajo reportado para otros estudios relacionados (115, 52.8 y 116.4 cm/ año para raíces de Asplundia, Clusia, Philodendron, respectivamente), esta investigación se realizó en bosques montanos sobre los 2100 msnm donde las tasas de crecimiento son más bajas que en bosques de tierras bajas (Bermeo 2010). Plowden et al. (2003) reportan en Heteropsis flexuosa (Araceae) un promedio de crecimiento de raíz por planta de 76.6 ± 22.3 cm/año en la amazonia brasilera. Patiño et al. (1999) reportan en Panamá (1300 m) para algunas especies de Clusia un crecimiento entre 73 y 182 cm/año por raíz y para algunas especies de Araceae entre 146 y 328 cm/año por raíz. García y Galeano (2009b) en Colombia (entre 1600 m y 2200 m) reportan para Philodendron longirrhizum tasas de crecimiento mensual de 31 cm/mes en raíces inmaduras, asumiendo un crecimiento lineal en un año podría alcanzar 372 cm. Si bien el presente estudio abarco siete meses del año (que correspondieron tanto con una fase lluviosa como con una fase de menos lluvia) los valores totales obtenidos podrían eventualmente ser ligeramente distintos en tasas anuales.
De acuerdo a la segunda parte de la hipótesis, esperamos mayores tasas de crecimiento de raíz en aquellas especies que realizan una menor inversión de recursos en la construcción de sus raíces (ejemplo: menor peso seco por cm). Encontramos que efectivamente Asplundia y Philodendron que invierten menos en la construcción de sus raíces, lo cual se refleja en un menor peso seco en sus raíces comparadas con Clusia, sus raíces crecen más rápidamente. El crecimiento acelerado de las raíces adventicias en algunas especies de lianas nómadas (sensu Zotz 2013) de aráceas al parecer está relacionado con la senescencia de los tallos, por lo cual, la formación de nuevas raíces y el crecimiento acelerado de las mismas corresponde a una estrategia adaptativa (Patiño et al. 1999, López-Portillo et al. 2000, Turriago-García 2013). No obstante, si bien las raíces de Philodendron crecen rápidamente en la zona de estudio, el bajo número de raíces no ancladas por individuo ocasiona una baja tasa de regeneración y en consecuencia una baja producción neta de fibras en relación con las otras dos especies. De acuerdo con estos resultados, entre las tres especies, Asplundia podría presentar mejores resultados en programas de aprovechamiento, sin embargo es necesaria cautela en la interpretación de estos resultados ante el desconocimiento de muchos aspectos biológicos de las especies como sus ciclos de vida y tasas de crecimiento, entre otros.
En conclusión, consideramos que el estado actual de las plantas factibles de cosechar de Philodendron, Clusia y Asplundia en el corregimiento de Santa Elena es bajo (<30 plantas). Encontramos que si bien el número de raíces por planta es considerable (M=10), el bajo número de plantas afecta la cantidad de fibra total disponible. Adicionalmente al bajo número de plantas encontradas, el tiempo de formación de una nueva raíz con el tamaño adecuado para ser cosechada supera los tres años para las tres especies. Así que el tiempo de renovación que necesitan las plantas para formar nuevas raíces "maduras" es relativamente alto, de ahí que la cosecha de realizarse, requiere un manejo controlado para garantizar la viabilidad de las plantas a largo plazo.
Este estudio muestra que la sostenibilidad de las especies productoras de fibra en Santa Elena es crítica. De acuerdo a nuestros resultados, es claro que el bajo número de individuos adultos de las especies y el crecimiento relativamente lento de las raíces no permiten desarrollar un modelo sostenible bajo las circunstancias actuales. No obstante, la implementación de planes de manejo a largo plazo para estas tres especies podría salvaguarda tanto las especies como el conocimiento tradicional asociados a ellas (Ticktin 2004). Evidencias en África, Asia y Latinoamérica demuestran que es posible encontrar la viabilidad ambiental y económica para la comercialización de PFNM (Kusters et al. 2006).
Adicionalmente, el conocimiento del uso de la biodiversidad y los productos elaborados a partir de las fibras de raíces adventicias podrían generar un valor agregado para los artesanos de la zona, el cual podría promover la conservación de los bosques, como se ha dado con otras experiencias en el mundo (Kusters et al. 2006). Sugerimos que programas de planeación regional y local deben incluir programas de cultivo (con material en viveros e in vitro) que incluyan enriquecimiento del bosque (Moegenburg et al. 2002, Guariguata et al. 2009) con estas tres especies, para garantizar una actividad sustentable para recolectores y artesanos en Santa Elena y otros bosques de la cordillera Central de los Andes.
AGRADECIMIENTOS
Este estudio se llevó a cabo gracias al apoyo recibido por el Programa de Colciencias "jóvenes investigadores N 566 de 2012", la Corporación Parque Arví y la Corporación para Investigaciones Biológicas. Agradecemos el apoyo recibido por la Corporación Parque Arví en especial a Diana Milena Amaya, quien facilitó la logística en el área. Agradecemos a las familias de la comunidad de los Vásquez en Santa Elena, especialmente a Jesús Parra y Jorge Vásquez y familia, a los guías del Parque Arví por la colaboración en la toma de datos. Agradecemos a la fundación Sentir por permitirnos el acceso a la Reserva El Robledal y a la Corporación Autónoma Regional del Centro de Antioquia (Corantioquia) por permitirnos el acceso a la Reserva Aguada. Este estudio contó con el permiso de investigación otorgado por Corantioquia número 17891.
LITERATURA CITADA
1. Armenteras, D., F. Gast & H. Villareal. 2003. Andean forest fragmentation and the representativeness of protected natural areas in the eastern Andes, Colombia. Biological Conservation 13: 245-256. [ Links ]
2. Balcázar-Vargas, M.P., T.R. Van Andel, P. Westers & P.A. Zuidema. 2015. What drives the vital rates of secondary hemiepiphytes? A first assessment for three species of Heteropsis (Araceae) in the Colombian Amazon. Journal of Tropical Ecology 31:251-265. [ Links ]
3. Belcher, B., M. Ruíz-Pérez & R. Achdiawan. 2005. Global patterns and trends in the use and management of commercial NTFPs: implications for livelihoods and conservation. World development 33:1435-1452. [ Links ]
4. Benavides, A.M., J.H.D. Wolf & J.F. Duivenvoorden. 2013. Colonization of forest clearings and tree-fall gaps in lowland rain forests of Colombia by hemiepiphytic aroids: experimental and transect studies. Journal of Tropical Ecology 29: 123-130. [ Links ]
5. Bernal, R., C. Torres, N. García, C. Isaza, J. Navarro, M.I. Vallejo & H. Balslev. 2011. Palm management in South America. The Botanical Review 77:607-646. [ Links ]
6. Bermeo-E, D.F. 2010. Determinación y caracterización de tipos funcionales de plantas (TFPs) en bosques secundarios dentro de un gradiente altitudinal y su relación con variables bioclimáticas. Tesis de Maestría, escuela de posgrado, Centro Agronómico Tropical de Investigación y Enseñanza. Turrialba, Costa Rica 126 pp. [ Links ]
7. Cockle, A. 2001. The dispersal and recruitment of Cyclanthaceae and Philodendron (Araceae) understorey root-climbing vines. Págs. 251-264 En: Nouragues. Springer Netherlands. Dordrecht, The Netherlands. [ Links ]
8. Croat, T. B. 1997. A revision of Philodendron subgenus Philodendron (Araceae) for Mexico and Central America. Annals of the Missouri Botanical Garden 84: 311-704. [ Links ]
9. Duque, A.J., T. Lobo-E, J.G. Marín, C. Zarate, J.L. Toro & F. Colorado. 2007. Introducción al aprovechamiento sostenible de musgos en el área de Piedras Blancas, Antioquia. Primera ed. A. J. Duque M, T. Lobo E, J. G. Marín, C. Zarate, J. L. Toro, & F. Colorado (eds.). Corporación Académica Ambiental, Universidad de Antioquia, Medellín. [ Links ]
10. Endress, B.A., D.L. Gorchov & R.B. Noble. 2004. Non-Timber Forest Product Extraction: Effects of Harvest and Browsing on an Understory Palm. Ecological Applications 14: 1139-1153. [ Links ]
11. Etter, A. & W. Van Wyngaarden. 2000. Patterns of Landscape Transformation in Colombia, with Emphasis in the Andean Region. Ambio 29: 432-39. [ Links ]
12. De Faria, A.P.G., G. Matallana, T. Wendt & F.R. Scarano. 2006. Low fruit set in the abundant dioecious tree Clusia hilariana (Clusiaceae) in a Brazilian restinga. Flora-Morphology, Distribution, Functional Ecology of Plants 201: 606-611. [ Links ]
13. Feuillet Hurtado, C., D. Macías Pinto & E. Chito Cerón. 2011. Plantas útiles para la elaboración de artesanías en el departamento del Cauca (Colombia). Boletín Científico. Centro de Museos. Museo de Historia Natural 15: 40-59. [ Links ]
14. Galeano, G., R. Bernal, C. Isaza, J. Navarro, N. García, M.I. Vallejo & C. Torres. 2010. Evaluación de la sostenibilidad del manejo de palmas. Ecología en Bolivia 45: 85-101. [ Links ]
15. Galeano, G. & R. Bernal-González. 1984. Nuevas Cyclanthaceae de Colombia. Caldasia 14: 27-35. [ Links ]
16. García, N. & G. Galeano. 2009a. Effect of the extraction of aerial roots on the populations of "tripeperro" (Philodendron longirrhizum, Araceae) in the Central Andes in Colombia. Caldasia 31: 19-29. [ Links ]
17. García, N. & G. Galeano. 2009b. Extracción sostenible de "tripeperro" (Philodendron longirrhizum M. Mora & Croat, Araceae) en los Andes centrales de Colombia. Revista Colombia Forestal 12: 25-36. [ Links ]
18. García, N., G. Galeano, R. Bernal & H. Balslev. 2013. Management of Astrocaryum standleyanum (Arecaceae) for handicraft production in Colombia. Ethnobotany Research & Applications 11:85-101. [ Links ]
19. Gómez Gómez, L.M. & C.M. Orozco-Castañeda 2001. Plantas de importancia artesanal en el centro del parque regional Arví. Corantioquia, Medellín 116 pp. [ Links ]
20. Guariguata, M.R., C.G. Nasi, R. Sheil, D. Herrero-Jáuregui, C. Cronkleton, P. Ndoye & O. Ingram-V. 2009. Hacia un manejo múltiple en bosques tropicales: Consideraciones sobre la compatibilidad del manejo de madera y productos forestales no maderables. CIFOR. Bogor, Indonesia. [ Links ]
21. Hammer, D. & A. Wildavsky. 1990. La entrevista semi-estructurada de final abierto. Aproximación a una guía operativa. Historia y fuente oral 4: 23-61. [ Links ]
22. Holos-Natura. 2001. Plan Maestro Parque Regional Arví. Informe de investigación. Medellín. [ Links ]
23. Kusters, K., R. Achdiawan, B. Belcher & M. Ruiz Pérez. 2006. Balancing development and conservation? An assessment of livelihood and environmental outcomes of nontimber forest product trade in Asia, Africa, and Latin America. Ecology and Society 11: 20. [ Links ]
24. Lanteri, M. L., Pagnussat, G. C., & Lamattina, L. 2006. Calcium and calcium-dependent protein kinases are involved in nitric oxide-and auxin-induced adventitious root formation in cucumber. Journal of Experimental Botany 57:1341-1351. [ Links ]
25. Liebig, M., F. Scarano, E. De Mattos, H. Zaluar & U. Lüttge. 2001. Ecophysiological and floristic implications of sex expression in the dioecious neotropical CAM tree Clusia hilariana Schltdl. Trees 15: 278-288. [ Links ]
26. Linares, E.L., G. Galeano, Y. Figueroa & N. Garcia. 2008. Fibras vegetales empleadas en artesanías en Colombia. Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales, Artesanías de Colombia S. A. Ministerio de Comercio, Industria y Turismo (Bogotá, Colombia), Bogotá D.C., Colombia. 328 pp. [ Links ]
27. Lynch, J. 1995. Root architecture and plant productivity. Plant physiology 109: 7-13. [ Links ]
28. Londoño, M. 2004. La comunidad de la Cuenca alta de Piedras Blancas y la explotación de los productos forestales no maderables de la zona, una evaluación participativa. Universidad Nacional de Colombia sede Medellín. Documento de Informe. [ Links ]
29. López-Portillo, J., F. Ewers, G. Angeles & J.B. Fisher. 2000. Hydraulic architecture of Monstera acuminata: evolutionary consequences of the hemiepiphytic growth form. New Phytologist 145: 289–245. [ Links ]
30. Macía, M.J. 2006. Las plantas de fibra. Botánica Económica de los Andes Centrales. Universidad Mayor de San Andrés, La Paz. [ Links ]
31. Mobot. 2014. Tropicos.org. Available at: http://www.tropicos.org (Acceso marzo 21, [ Links ]
32. Moegenburg, S.M. & D.J. Levey. 2002. Prospects for conserving biodiversity in Amazonian extractive reserves. Ecology Letters 5: 320-324. [ Links ]
33. Navarro López, J. A., G. Galeano & R. Bernal. 2014. Management of the chonta palm (Iriartea deltoidea Ruiz & Pav.) in the Amazon foothills of Colombia, perspectives for sustainable harvesting. Colombia Forestal 17:5-24. [ Links ]
34. Negi, S., P. Sukumar, X. Liu, J.D. Cohen & G.K. Muday. 2010. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. The Plant Journal 61: 3-15. [ Links ]
35. Niklas, K.J. & B.J. Enquist. 2001. Invariant scaling relationships for interspecific plant biomass production rates and body size. Proceedings of the National Academy of Sciences 98: 2922-2927. [ Links ]
36. Patiño, S., G.S. Gilbert, G. Zotz & M.T. Tyree. 1999. Growth and survival of aerial roots of hemiepiphytes in a lower montane tropical moist forest in Panama. Journal of Tropical Ecology 15: 651-665. [ Links ]
37. Pérez-Harguindeguy, N., S. Díaz, E. Garnier, S. Lavorel, H. Poorter, P. Jaureguiberry, M.S. Bret-Harte, W.K. Cornwell, J.M. Craine, D.E. Gurvich, C. Urcelay, E.J. Veneklaas, P.B. Reich, L. Poorter, I.J. Wright, P. Ray, L. Enrico, J.G. Pausas, A.C. De Vos, N. Buchmann, G. Funes, F. Quétier, J.G. Hodgson, K. Thompson, H.D. Morgan, H. Ter Steege, M.G.A. Van der Heijden, L. Sack, B. Blonder, P. Poschlod, M.V. Vaieretti, G. Conti, A.C. Staver, S. Aquino & J.H.C. Cornelissen. 2013. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany 61: 167-234. http://dx.doi.org/10.1071/BT12225 [ Links ]
38. Plowden, C., C. Uhl & F.D.A. Oliveira. 2003. The ecology and harvest potential of titica vine roots (Heteropsis flexuosa: Araceae) in the eastern Brazilian Amazon. Forest Ecology and Management 182: 59-73. [ Links ]
39. Poorter, H. & O. Nagel. 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Functional Plant Biology 27:1191-1191. [ Links ]
40. Ramos, A. 2000. Hacia un uso sostenible de las materias primas vegetales artesanales. Biosintesis 17: 1-4. [ Links ]
41. Ray, T.S. 1992. Foraging behaviour in tropical herbaceous climbers (Araceae). Journal of Ecology 80: 189–203. [ Links ]
42. R Development Core Team. 2008. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Viena, Austria. [ Links ]
43. Royer, D.L. 2008. Nutrient turnover rates in ancient terrestrial ecosystems. Palaios 23: 421-423. [ Links ]
44. Siata. 2011. Sistema de alerta temprana del Área Metropolitana del Valle de Aburrá. disponible en: http://siata.gov.co/newpage/index.php. [ Links ]
45. Ticktin, T. 2004. The ecological implications of harvesting non-timber forest products. Journal of Applied Ecology 41: 11–21. [ Links ]
46. Turriago-García, J.D. 2013. Ecología funcional de raíces aéreas absorbentes del Yare (Heteropsis spp (Kunt)) en bosques de tierra firme de la Amazonia Colombiana. Universidad Nacional de Colombia Sede Amazonia. [ Links ]
47. Wickens, G.E. 1991. Management issues for development of non-timber forest products. Unasylva 42: 3-8. [ Links ]
48. Wright, I.J., P.B. Reich, M. Westoby, D.D. Ackerly, Z. Baruch, F. Bongers & R. Villar. 2004. The worldwide leaf economics spectrum. Nature 428: 821-827. [ Links ]
49. Vanneste, S. & J. Friml. 2009. Auxin: a trigger for change in plant development. Cell 136: 1005-1016. [ Links ]
50. Visser, E.J., J.D. Cohen, G.W. Barendse, C.W. Blom & L.A. Voesenek. 1996. An ethylene-mediated increase in sensitivity to auxin induces adventitious root formation in flooded Rumex palustris Sm. Plant Physiology 112:1687-1692. [ Links ]
51. Zotz, G. 2013. 'Hemiepiphyte': a confusing term and its history. Annals of Botany 111:1015-1020. [ Links ]
