










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.38 no.1 Bogotá Jan./June 2016
https://doi.org/10.15446/caldasia.v38n1.57834
PERLA NATALIA RAMÍREZ-NARVÁEZ
PATRICIA VELASCO-LINARES
Subdirección Científica, Jardín Botánico José Celestino Mutis, Bogotá, D. C., Colombia. perlanatalia8@gmail.com, patriciavelascolinares@gmail.com
RESUMEN
La floración de los bambúes es poco frecuente, tiende a ser esporádica cuando algunos culmos de la población florecen o gregaria, donde todos los individuos florecen al mismo tiempo y en diferentes zonas. Algunas especies de bambúes leñosos mueren después de florecer y otras no. Chusquea scandens Kunth es un bambú leñoso muy abundante en zonas de alta montaña cercanas a Bogotá. En cada población se evaluaron las etapas fenológicas y la tasa de crecimiento de diez culmos en estado vegetativo y diez en inicio de la floración. En la población que presentó un área extensa de flores, se indagó qué le sucedió a C. scandens después de la floración. La asociación de C. scandens con otras especies tanto de bosque nativo como exóticas, pudo ser uno de los factores que indujo su floración. El parche florecido presentó secamiento y posterior colonización por especies pioneras. Durante el tiempo de estudio C. scandens, presentó evidencias de ser una especie monocárpica. La estrategia de floración de C. scandens fue diferente a otras especies del género Chusquea, respecto al menor tiempo que duró un culmo en pasar de botones a frutos con semillas y en que presentó características de floración esporádica.
Palabras clave. Fenología, bambú, sucesión, monocarpía.
ABSTRACT
Flowering of bamboos is rare, it tends to be sporadic when some culms flower or gregarious, when all the individuals in the population flower at the same time and in different areas. Some species of woody bamboos die after flowering while others do not. Chusquea scandens Kunth is a woody bamboo very abundant in high mountain ecosystems and populations in the study area were flowering. To determine the characteristics of the flowering of C. scandens, we selected two populations located in a rural area of Bogotá. On each population we evaluated the phenological stages and growth rate often vegetative culms and ten in the beginning of flowering. In the population that presented a wide area of flowers, it was investigated what happened to C. scandens after flowering. The association of C. scandens with other species of both native and exotic forests, could be one of the factors influencing its flowering. The flowered patch showed drying and subsequent colonization by pioneer species. During the study period C. scandens showed evidence of being a monocarpic species. The strategy of flowering C. scandens, was different from other species of Chusquea, in the shorter time a culm developed from a bud to a fruit with seeds well as in its sporadic flowering.
Key words. Phenology, bamboo, succession, monocarpic.
Recibido: 01/05/2015
Aceptado: 25/02/2016
INTRODUCCION
Chusquea es uno de los géneros de Poaceae más diversos, con especies distribuidas desde México hasta Chile y Argentina (McClure 1978, Clark & Ely 2011). En Colombia se halla entre los más ricos en especies de la familia Poaceae (Giraldo-Cañas 2011). La mayoría de los bambúes, incluyendo especies del género Chusquea, tardan varios años en florecer. El evento de floración tiende a iniciar en algunas plantas aisladas; posteriormente, la mayor parte florece simultáneamente y presenta una muerte sincrónica de los individuos de la población (Janzen1976). Las especies con esta forma de reproducción son denominadas monocárpicas, porque mueren después de generar semillas (González 2001). La floración puede iniciar en algunos parches y después cubrir extensas áreas (Janzen1976). Como lo menciona González (2001), en Chile varias de las especies del género Chusquea florecen en un período estimado de 15 a 70 años, donde Chusquea quila Kunth al florecer cubrió aproximadamente un millón de hectáreas. Especies como Chusquea culeou É.Desv. en Argentina, pueden tardar más de 60 años en florecer y morir; el parche florecido tiende a cubrir más de doscientas mil hectáreas (Victoria et al. 2009). La monocarpía presenta ventajas adaptativas, al permitir a una especie mantenerse en un sitio previamente ocupado, con regeneración de su población (Montti et al. 2011). También evita que continuamente sean traslocados recursos para fructificar, no dejando que los culmos sean debilitados y por ende, más susceptibles a ataques de hongos e insectos (Edwards-Widmer 1998).
Una de las características de las especies monocárpicas es que se secan posterior a un evento de floración (González 2001) y se tiende a generalizar que Chusquea scandens Kunth (chusque) por pertenecer a los bambúes leñosos es una especie monocárpica. Sin embargo, se ha reportado en Miyazaki et al. (2009) que no todos los bambúes leñosos cumplen con este rasgo de historia de vida.
En el bosque altoandino tropical, C. scandens es una de las especies con mayor abundancia (Delgado et al. 2009). Se encuentra en quebradas, nacimientos de agua y en bosques alterados, donde limita el establecimiento de otras especies nativas (Trujillo et al. 2007). Por lo tanto, se sugiere que en el borde de bosque con C. scandens, sea controlada para promover la expansión del bosque (Montenegro & Vargas 2008).
Chusquea scandens presenta una distribución desde Colombia hasta Bolivia (Ohrnberger & Goerrings 1984), con rangos altitudinales en los Andes entre los 2500 y los 3500 m (Giraldo-Cañas 2011). En el presente estudio en algunas localidades de Bogotá, se observaron parches de C. scandens en proceso de secamiento que habían presentado floración. Por ende, la presente investigación tuvo como objetivos conocer qué características tienen las etapas fenológicas de C. scandens durante su floración, cuál es la frecuencia de floración y tasa de crecimiento de los culmos florecidos. También después de la floración, evaluar la colonización de especies, la generación de semillas, establecimiento de plántulas y contenido hídrico. Se tuvo como hipótesis que la estrategia de floración de C. scandens, es diferente respecto a otras especies del género Chusquea. Lo anterior contribuye al conocimiento fenológico de una especie, que ocupa extensas áreas de ecosistemas de la alta montaña tropical andina y que florece de manera poco frecuente.
MATERIALES Y MÉTODOS
Área de estudio
Se seleccionaron dos poblaciones donde C. scandens es dominante, en zonas adyacentes al área urbana de Bogotá, que presentaron eventos de floración. La primera población fue en la Reserva Forestal El Delirio y la segunda en la Vereda el Verjón. La Reserva Forestal El Delirio se localiza en la Localidad San Cristóbal en estribaciones del Páramo de Cruz Verde, a 2810 m (4°33'2.29"N y 74°35'67"O); hace parte de la superficie de reserva forestal protectora de los Cerros Orientales de Bogotá. Es el lugar de nacimiento del río Fucha, presenta ecosistemas de subpáramo y bosque altoandino. La Vereda el Verjón se ubica en la Localidad Santa Fé en la vía Choachí, a 3044 m (4°35'2.98"N y 74°22'41"O). Posee vegetación de bosque altoandino, subpáramo y páramo. Ambas poblaciones presentan actividades humanas relacionadas con protección de la fauna y flora, caminatas ecológicas y educación ambiental. Sin embargo, las áreas de estudio han tenido una historia de disturbio, ocasionada por cultivos, siembra de especies exóticas y construcción de obras civiles. En la vegetación exótica, dominan especies como Acacia melanoxylon, Acacia decurrens, Eucalyptus globulus, Genista monspessulana y Ulex europeaus. Las áreas presentan un comportamiento climático bimodal, con dos épocas de presencia de lluvias alternando con periodos secos. Las épocas de mayor cantidad de lluvias se presentan de abril a mayo y de octubre a noviembre.
Diseño de muestreo
Características de las etapas fenológicas de C. scandens durante su floración. Las poblaciones de estudio presentaron la tendencia a distribuirse de manera homogénea, ocupando parches de aproximadamente 300 metros de distancia y con eventos de floración en sus culmos. Se evaluaron los culmos, por ser un tallo de la planta formado por yemas, nudos y entrenudos; donde en los nudos se originan las hojas y espigas florales.
En cada población se realizó un transecto de 1500 x 2 metros, en el área donde se presentó la floración. La ubicación del transecto se seleccionó, con base en el inicio de la presencia de puntos de floración y la posición fue aleatoria respecto a los tipos de cobertura vegetal en la zona. Estudiar el fenómeno de floración en más localidades de la región, hubiera permitido contar con un estudio replicado, sin embargo, no se presentaron eventos grandes de floración en otras poblaciones.
De los culmos presentes en el transecto, fueron seleccionados y marcados veinte culmos de C. scandens de manera aleatoria para ser evaluados periódicamente, examinando los cambios fenológicos asociados a su floración; diez culmos en estado vegetativo y los otros diez en etapa de botón o inicio de la floración. El estudio de la fase vegetativa inició en noviembre del 2012 y se tomaron datos de los cambios fenológicos, cada quince días a partir del inicio de la floración de los culmos, desde agosto del 2013 hasta mayo del 2014. En mayo del 2015, se evaluó si había presencia de nuevos eventos de floración en las poblaciones de estudio.
Las fases fenológicas se definieron así: i) vegetativa donde los culmos no presentaron floración; ii) botón, como el período inicial de emisión de los tallos florales; iii) floración, donde fue posible apreciar los estambres expuestos; y iv) frutos secos con semillas (Figura 1). También en el transecto se evaluó la presencia de especies vegetales vasculares, asociadas al parche en floración.

Frecuencia de floración de C. scandens. En el transecto de 1500 x 2 metros se midió la frecuencia de floración de C. scandens, definida como el número de veces que se presentaron culmos florecidos.
Crecimiento de los culmos de C. scandens en floración. Cada mes en los culmos en floración seleccionados y marcados, se midió la altura desde la base del culmo hasta la yema del ápice más alto.
Características después de la floración de C. scandens. En el transecto ubicado en la Vereda el Verjón, se presentó un área extensa de floración, que permitió en mayo del 2014 evaluar en una parcela de 20x20 m, la cobertura de todas las especies y la presencia o ausencia de rebrotes en los culmos florecidos. La cobertura de cada especie se estableció como el porcentaje que ocupaba en cada estrato. Se establecieron seis estratos propuestos por Rangel &Velázquez (1997), rasante (<0,3 m), herbáceo (0,3-1,5 m), arbustivo (1,5-5m), subarbóreo (5-12m), arbóreo inferior (12-25 m) y arbóreo superior (>25 m). No fue posible replicar más parcelas en la localidad u otras áreas de la región, porque no se presentaron eventos de floración del tamaño evaluado. En la parcela de 20x20 m se estableció el porcentaje de generación de semillas, seleccionando 300 frutos de los culmos secos y posteriormente se extrajo manualmente las semillas. Estas semillas fueron sembradas en condiciones de invernadero, superficialmente en una bandeja germinadora con tierra negra tamizada y esterilizada. Dos veces por semana durante seis meses, se realizó un seguimiento para determinar el número de semillas germinadas por día.
El establecimiento de plántulas de C. scandens se evaluó en mayo del 2014, porque paso un periodo seco y comenzó la época de lluvias, lo cual se consideró un indicio para que las semillas germinaran. Se realizaron observaciones cada metro cuadrado de la parcela de 20x20 m, con un cuadrante de 1x1 m con divisiones en forma de retícula cada 10 cm, teniendo un total de 400 repeticiones.
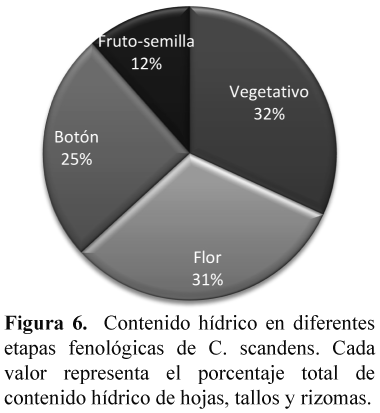
Para determinar el contenido hídrico después de la floración, de tres individuos tomados al azar de cada fase fenológica (estado vegetativo, botones, flor y frutos con semillas), se realizó la toma de muestras de hojas, tallos y rizomas. Posteriormente se pesaron 5 gr de cada órgano (hojas, tallos y rizomas) en cada fase fenológica, conformados de los tres individuos muestreados, determinando el peso fresco y se llevaron a la estufa a temperatura de 80 °C, durante 24 horas para establecer el peso seco.
Análisis de datos
Para determinar las características de las etapas fenológicas de C. scandens durante su floración, en cada población de los diez culmos evaluados, se determinó el promedio ± error estándar de botones, flores y frutos con semillas; y se realizó una representación gráfica de las fases fenológicas de C. scandens en ambas poblaciones. Se analizó la frecuencia de floración de C. scandens y las especies vegetales asociadas.
Para establecer el crecimiento de los culmos, se calculó la tasa de crecimiento de la altura aplicando la ecuación, presentada por Hunt (1990): TC= (Variable final-Variable inicial/Tiempo final-Tiempo inicial). Se determinó la media y error estándar de las características morfológicas de los culmos. Con pruebas de Kolmogorov-Smirnov, utilizando el programa estadístico PAST (Hammer et al. 2001), se evaluó la existencia de diferencias entre las poblaciones.
Para analizar las características después de la floración, en la población ubicada en la Vereda el Verjón, se determinó el porcentaje de cobertura de las especies que colonizaron el parche florecido. Respecto a la generación de semillas y plántulas, se realizó una descripción del proceso, porque se obtuvieron datos cualitativos. Se calculó el contenido hídrico, con base en la ecuación propuesta por Sánchez & Aguirreolea (2000): CH= (Pf-Ps/Ps)100, donde CH es contenido hídrico, Pf peso fresco de la muestra y Ps peso seco de la muestra.
RESULTADOS
Características de las etapas fenológicas de C. scandens durante su floración
C. scandens presentó flores bisexuales, inflorescencia terminal, por lo general con un eje central diversamente ramificado, tres estambres y una lemna. Se mantuvo la fase vegetativa desde noviembre del 2012 hasta agosto del 2013, momento en que se dio inicio a la floración en ambas poblaciones. No se presentaron diferencias significativas entre la sumatoria de botones, flores y frutos con semillas entre ambas poblaciones (α=0,05; K-S=0,21; p=0,10). Sin embargo, los valores más altos se hallaron en la población ubicada en la Vereda el Verjón, porque el tamaño de los culmos fue mayor (Figura 2).
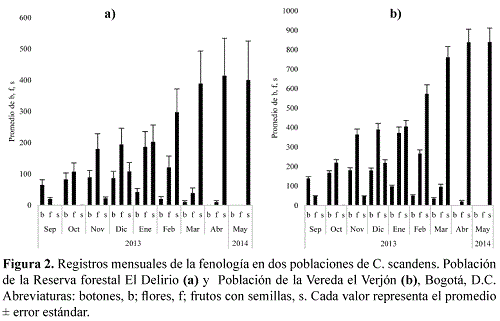
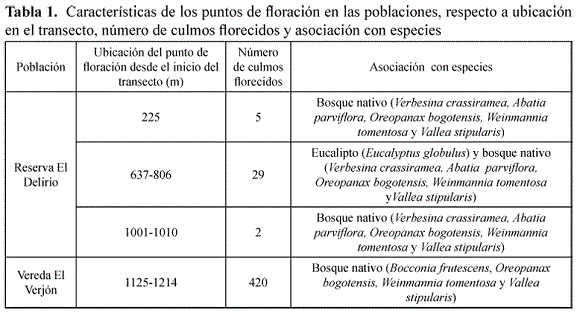
En ambas poblaciones en el primer mes del primer año de muestreo de floración, se halló la mayor cantidad de botones, pocas flores y ningún fruto seco. Se observó un incremento gradual de botones y flores de septiembre a diciembre del 2013 y a partir de enero del 2014 una disminución. Se halló un aumento en el número de frutos con semillas, posterior al mes de enero del 2014; después los frutos con semillas comenzaron a caer. En mayo del 2015 no se volvieron a presentar eventos de floración en las dos poblaciones. En el transcurso de un mes, hubo paso de botones a flores en ambas poblaciones. Cerca de ocho meses duró un culmo en fase de botones, para pasar a fase de frutos con semillas, si hubo polinización (Figura 3). Las especies vegetales asociadas con el parche en floración, se presentan en la Tabla 1.

Frecuencia de floración de C. scandens
Se presentaron diferencias significativas en la frecuencia de floración, entre las dos poblaciones (α=0,05; K-S=0,04; p=0,00). La población de la Reserva Forestal El Delirio presentó el número de culmos florecidos en menor cantidad y ubicados de manera separada. En la Vereda el Verjón el punto de floración fue continuo y con mayor número de culmos florecidos. En ambas poblaciones en parches monoespecíficos de C. scandens no se presentó floración. La floración se dio cuando en el parche había otras especies, tanto nativas como exóticas (Tabla 1).
Crecimiento de los culmos de C. scandens en floración
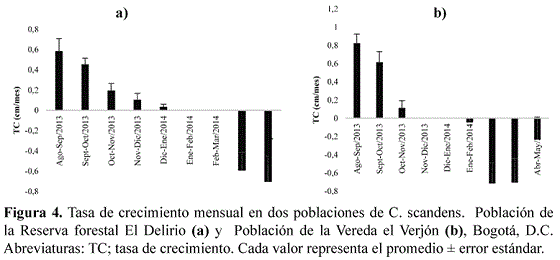
En la población ubicada en la Reserva forestal El Delirio, desde agosto hasta enero del 2014 algunos culmos en floración presentaron crecimiento. En los meses siguientes se detuvo el crecimiento y el culmo inició a secarse, representado por valores negativos, ya que los ápices tuvieron la tendencia a partirse. En la población de la Vereda el Verjón se presentó un aumento en la tasa de crecimiento de agosto hasta noviembre; posteriormente se detuvo el crecimiento y se secaron los culmos (Figura 4). Se determinó que no hubo diferencias significativas entre las tasas de crecimiento de las dos poblaciones (α=0,05; K-S=0,22; p=0,95).
Características del parche después de la floración de C. scandens
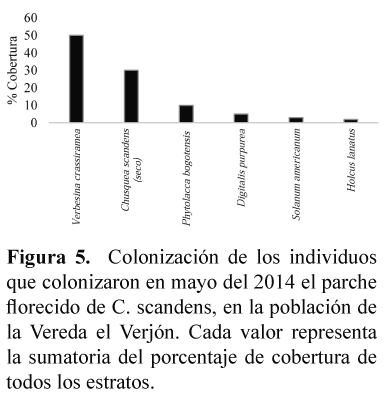
Las especies que colonizaron en mayor porcentaje de cobertura el parche en secamiento ubicado en la Vereda el Verjón, en el estrato arbóreo fueron Verbesina crassiramea y en el estrato herbáceo Phytolacca bogotensis (Figura 5). Alrededor del parche florecido, se observó que los árboles V. crassiramea y Abatia parviflora cuya altura superaba los 18 metros, no fueron cubiertos por C. scandens. Otros culmos de C. scandens en la misma población no florecieron y tuvieron la tendencia a colonizar las zonas que se secaron.
La generación de semillas fue baja, ya que de 300 frutos secos solo un 7% presentó semillas. Durante la disección de los frutos secos se halló que varios tenían en lugar de semillas larvas, que probablemente consumieron la semilla. No se presentaron plántulas en la prueba de germinación, ni en la parcela de evaluación del parche florecido.
El mayor porcentaje de contenido hídrico lo tuvieron las muestras de las hojas, tallos y rizomas del estado fenológico vegetativo; seguido de las muestras en estado de flor, botón y fruto-semilla (Figura 6).
DISCUSIÓN
Características de las etapas fenológicas de C. scandens durante su floración
De septiembre a diciembre del 2013 hubo mayor paso de botones a flor, lo que coincidió con el inicio del periodo de lluvias y en el periodo seco comenzó el mayor paso de flores a frutos con semillas. En especies como Chusquea talamancensis, también en la estación lluviosa aumentó la formación de flores y en la estación seca la maduración de frutos (Edwards-Widmer 1998). El proceso de floración de C. scandens fue dinámico en el tiempo y el paso promedio de un culmo de botones a frutos con semillas fue cercano a los ocho meses. Este tiempo fue menor al reportado en otras especies del género Chusquea, como C. quila que presenta un período reproductivo desde la floración hasta la diseminación de frutos aproximado de veinte meses (González 2001). También fue menor que en otros bambús, donde la fructificación puede requerir hasta un año a partir de la iniciación de la floración, hasta la caída de la última semilla (Janzen 1976).
Frecuencia de floración de C. scandens
Los puntos de floración se presentaron cuando C. scandens estuvo asociada a otras especies tanto nativas como exóticas. Por ende, se postula la hipótesis que la floración fue inducida por un tipo de estrés causado por la competencia con las otras especies, ya que en los parches continuos de crecimiento vegetativo no se observaron culmos florecidos. En Mercedes (2006) se presentan otras teorías que tratan de explicar las causas de la floración en bambús: a) la teoría periódica que asegura que el ciclo de regeneración del bambú, a través de métodos asexuales por medio de rizomas y el alargamiento de la caña, alcanza la madurez cuando la planta florece; b) la teoría de mutación que considera que la regeneración del bambú a través de cualquier método de propagación sexual es una mutación que provoca la floración; y c) la teoría de la nutrición que propone que la floración y la fructificación son el resultado de problemas fisiológicos, que provienen mayormente de un pobre crecimiento de las células, ocasionado por un desbalance de la relación Carbono-Nitrógeno. Otra teoría propuesta en Janzen (1976) para explicar el ciclo de floración de los bambúes, gira en torno a un mecanismo de saciar a los depredadores con abundancia de alimento representada por toneladas de semillas, obteniendo una mayor probabilidad de germinar las semillas que no son consumidas.
Crecimiento de los culmos de C. scandens en floración
La disminución del crecimiento hasta detenerse y posterior secamiento de los culmos florecidos, son evidencias que C. scandens es una especie monocárpica, al igual que otras especies del mismo género como C. quila (González 2001). Se ha de tener en cuenta que dicho comportamiento en C. scandens, fue a nivel de un ramet (equivalente a un culmo) y no a nivel de un genet (individuo genético).
Características después de la floración de C. scandens
Los culmos de C. scandens en la misma población que no florecieron y tuvieron la tendencia a colonizar las zonas que se secaron, son evidencias que pueden coincidir con el estudio llevado a cabo por Miyazaki et al. (2009), donde se excavaron los rizomas de varios culmos florecidos y se conectaban a culmos que no florecieron, revelando que no se secó todo el individuo. Esto los llevó a plantear la hipótesis, que los culmos se secaron por el alto costo de reproducción y no porque fuera una especie monocárpica.
La mayor extensión de floración se presentó en la Vereda el Verjón, ya que en la Reserva el Delirio, quizás porque continuamente realizan podas, no se observó un parche grande florecido. En otras poblaciones ubicadas en la Quebrada Yomasa, Quebrada Bolonia, Quebrada Santa Librada, Monserrate y Usaquén, se presentaron en cada población en el mismo año, cerca de cinco culmos florecidos, de manera aislada, los cuales se secaron. Sin embargo, la población entera no se secó, porque permanecieron los otros culmos en estado vegetativo.
Los resultados obtenidos sugieren que C. scandens tiene un tipo de floración esporádica, porque no todos los culmos de la población florecieron. Estas características no coinciden con otras especies de bambúes, que presentan floración gregaria de los individuos de la población como Phyllostachys bambusoides (Mercedes 2006), C. quila (González 2001) y C. culeou (Victoria et al. 2009).
En la vereda El Vejón, posterior a la formación de frutos con semillas, se dio la colonización de V. crassiramea, P. bogotensis y Digitalis purpurea, todas ellas heliófilas. El crecimiento de V. crassiramea coincide con la descripción de Franco & Vargas (2009), que presenta una rápida elongación del tallo y plasticidad morfológica, que le permiten aprovechar los claros y ubicar su follaje fuera de la matriz. P. bogotensis, tuvo una cobertura homogénea, quizás influenciada porque forma bancos de semillas que contribuyen en la regeneración del bosque (Cantillo et al. 2008) y que se encuentra en vegetación de ribera (Rangel-Ch. & Aguirre-C 1983). La presencia de D. purpurea, pudiera estar asociada a que domina el banco de semillas en bordes de bosque con presencia de C. scandens (Trujillo & Vargas 2007).
La producción de semillas fue baja, probablemente porque en los frutos secos se halló que varios contenían en lugar de semillas larvas de insecto, que como producto de la depredación pueden haber consumido la semilla, como se ha detectado en otras plantas (Kitzberger et al. 2007; Perea 2012). La baja producción de semillas, además, pudo estar asociada a la poca polinización durante la floración, ya que en otras especies de bambú como Melocanna baccifera, se ha reportado que presenta pocos estigmas e ineficiente polinización (Ramanayake & Weerawardene 2003). También es reportada una baja producción de semillas en el primer año de floración de C. talamancensis, sin embargo, en los dos años subsecuentes vuelve a florecer y deja descendencia (Edwards-Widmer 1998). Aunque las poblaciones de C. scandens evaluadas no volvieron a florecer, en un lapso cercano a los tres años, como sí ocurrió con C. talamancensis. Otros bambúes también reportan producción de semillas baja (0,1%), quizás por haber invertido un alto costo de reproducción en florecer (Miyazaki et al.2009).
No se halló germinación de plántulas, lo que tiende a estar asociado al área de muestreo pequeña, debido a que no se presentaron más eventos continuos de floración, que permitieran réplicas en otras poblaciones. A que falto evaluar en un lapso mayor de tiempo, porque en especies como C. talamancensis las semillas germinan dos años después de la floración (Edwards-Widmer 1998). También, puede asociarse al reporte de (Pérez 2006), que los cariópsides sembrados no hayan presentado embriones maduros.
La tendencia de los parches florecidos de C. scandens, fue a ser colonizados por especies pioneras, como V. crassiramea que posee semillas que logran germinar en ausencia de luz, estableciéndose rápidamente en claros (Franco & Vargas 2009); por lo tanto, pudieran generar competencia para el establecimiento de las plántulas de C. scandens.
El menor porcentaje de contenido hídrico lo presentaron las muestras de los culmos que se hallaron en fruto con semillas, lo cual es uno de los indicios que confirman la monocarpia de C. scandens, puesto que la pérdida de humedad en hojas, tallos y rizomas, representa la muerte del culmo. Se estableció que los culmos de C. scandens posterior al florecimiento se secaron y murieron; como es reportado en otras especies monocárpicas del mismo género (González 2001).
Se sugiere continuar con el estudio a largo plazo para determinar la dinámica de la floración de C. scandens. Es prioritario establecer si C. scandens es monocárpica, realizando la excavación de los rizomas florecidos y determinar si se conectan a culmos que no florecieron. Esta información se puede complementar mediante estudios genéticos, para establecer si los culmos que florecen pertenecen al mismo individuo que permanece en estado vegetativo. Finalmente, estas nuevas investigaciones permitirán plantear nuevas teorías, sobre los mecanismos que activan la floración de C. scandens.
AGRADECIMIENTOS
A la Subdirección Científica del Jardín Botánico José Celestino Mutis por su apoyo en la investigación y al Acueducto de Bogotá por autorizar el ingreso a la Reserva el Delirio. También a los evaluadores por sus valiosas observaciones.
LITERATURA CITADA
1. Boom, A., R. Marchant, H. Hooghiemstra & J. Damste. 2002. CO2- and temperature-controlled altitudinal shifts of C4- and C3-dominated grasslands allow reconstruction of palaeoatmospheric pCO2. Palaeogeography, Palaeoclimatology, Palaeoecology 177:151-168. [ Links ]
2. Cantillo, E. E., V. Castiblanco, D.F. Pinilla & C.L. Alvarado. 2008. Characterization and evaluation of the regeneration potential of the seed bank of the Cárpatos Forest Reserve (Guasca, Cundinamarca). Revista Colombia Forestal 11(21): 45-70. [ Links ]
3. Clark, L.G. & F.U. Ely. 2011. Géneros de bambúes leñosos (Poaceae: Bambusoideae: Arundinarieae, Bambuseae) de Venezuela.Acta Botánica Venezuelica 34: 79-103. [ Links ]
4. Delgado, A., L. Galindo, L.M. Buitrago, N. Céspedes, E.M. Vargas, M. Suarez. & E. Ussa. 2009. Reflexiones en torno a la Educación Ambiental en Bogotá a partir de la implementación de Escenarios Vivos de Aprendizaje, EVA, en siete quebradas del Distrito Capital. Perez Arbelaezia 19: 15-34. [ Links ]
5. Edwards-Widmer, Y. 1998. Flowering phenology of Chusquea-bamboos with special reference to Chusquea talamancensis in Costa Rica. En: Y. Edwards-Widmer. 1999. The ecological role of Bamboo (Chusquea spp.) in the old-growth Quercus forests of the Cordillera de Talamanca, Costa Rica: 115-138. Swtss Federal Institute of Technology Zürich.Dissertation for the degree of Doctor of Natural Sciences, Zürich. [ Links ]
6. Franco, L & O. Vargas. 2009. Rasgos de Verbesina crassiramea blake de importancia en estrategias de control de especies invasoras en los alrededores del embalse de Chisacá. En: O.V argas, O. León & A. Díaz (eds). Restauración Ecológica, en zonas invadidas por retamo espinoso y plantaciones forestales de especies exóticas: 148-176.Universidad Nacional de Colombia, Bogotá D.C. [ Links ]
7. Giraldo-Cañas, D. 2011. Catálogo de la familia Poaceae en Colombia. Darwiniana 49(2): 139-247. [ Links ]
8. González, E. 2001. Fenología de Chusquea quila durante su floración gregaria en la zona centro-sur de Chile. Bosque 22(2): 45-51 [ Links ]
9. Hammer, O., D. Harper & P. Rian. 2001. Past: Palaeonthological statistics software package for education and data analysis. Versão. 1.37. http://palaeo-electronica.org/2001_1/past/issue1_01.htm (último acesso: 10/02/2016). [ Links ]
10. Hunt, R. 1990. Basic growth analysis: plant growth analysis for beginners.Unwin Hyman, Londres.112 pp. [ Links ]
11. Janzen, D.H. 1976. Why Bamboos wait so long to flower.Annual Review Ecology Systematics 7: 347-391. [ Links ]
12. Kitzberger, T., E.J. Chaneton & F. Caccia. 2007. Indirect effects of prey swamping: differential seed predation during a bamboo masting event. Ecology 88(10): 2541–2554. [ Links ]
13. Mercedes, J. 2006. Guía Técnica Cultivo del Bambú. Centro para el Desarrollo Agropecuario y Forestal, Inc. CEDAF. Serie Recursos Naturales - Guía Técnica. República Dominicana Santo Domingo. 37 pp. [ Links ]
14. Miyazaki, Y., N. Ohnishi, H. Takafumi & T. Hiura. 2009. Genets of dwarf bamboo do not die after one flowering event: evidence from genetic structure and flowering pattern. Journal of Plant Research 122:523–528. [ Links ]
15. Montenegro, A.L & O. Vargas. 2008. Caracterización de bordes de bosque altoandino e implicaciones para la restauración ecológica en la Reserva Forestal de Cogua (Colombia). Rev. Biol. Trop. 56(3): 1543-1556. [ Links ]
16. Montti, L., P.I. Campanello. & G. Goldstein.2011. Flowering, die-back and recovery of a semelparous woody bamboo in the Atlantic Forest. Acta Oecologica 37: 361-368. [ Links ]
17. Ohrnberger, D. & J. Goerrings. 1984.The Bamboos of the World- A Preliminary Study of the Names and Distribution of the Herbaceous and Woody Bamboos (BambusoideaeNees v. Esenb.) Documented in Lists and Maps.Journal of the American Bamboo Society 5: 1-2. [ Links ]
18. Perea, R. 2012. Dispersión y predación de semillas por la fauna: Implicaciones en la regeneración forestal de bosques templados. Ecosistemas 21 (1-2): 224-229. [ Links ]
19. Pérez, B. 2006. Determinar los factores ecofisiológicos de la germinación y crecimiento de especies andinas y consolidar los protocolos de manejo, en condiciones de banco plántular. Informe final-Contrato 165 del 2006. Jardín Botánico José Celestino Mutis, Bogotá D.C. 102 p. [ Links ]
20. Ramanayake, S.M.S.D. & T.E. Weerawardene. 2003. Flowering in a bamboo, Melocannabaccifera (Bambusoideae: Poaceae). Botanical Journal of the Linnean Society 143: 287–291. [ Links ]
21. Rangel-Ch., O & J. Aguirre-C. 1983. Comunidades acuáticas altoandinas-I vegetación sumergida y de ribera en el Lago de Tota, Boyacá, Colombia. Caldasia 13(65): 719-742. [ Links ]
22. Rangel-Ch., J.O. & A. Velázquez. 1997. Métodos de estudio de la vegetación. En: J.O. Rangel-Ch. (ed.). Diversidad Biótica II. Instituto de Ciencias Naturales:59-87. Universidad Nacional de Colombia. Bogotá D.C. [ Links ]
23. Sánchez, M. & A. Aguirreolea. 2000. El agua en la planta. En: J. Azcon & M. Talon (eds). Fundamentos de Fisiología Vegetal: 17-30.McGraw-Hill Interamericana, Barcelona. [ Links ]
24. Statical Graphics Corp. 1999. Statgraphics Plus, Statical Graphics Corp. [ Links ]
25. Trujillo, L., A. López & O. Vargas. 2007.Lluvia de Semillas en Bordes de Bosque. En: O. Vargas & Grupo de Restauración Ecológica Universidad Nacional de Colombia (Eds).Estrategias para la Restauración del Bosque Altoandino: El Caso de la Reserva Forestal de Cogua (Cundinamarca):282-293. Universidad Nacional de Colombia. Bogotá D.C. [ Links ]
26. Trujillo, L. & O. Vargas. 2007. Bancos de semillas en bordes de bosque. En: O. Vargas & Grupo de Restauración Ecológica Universidad Nacional de Colombia (eds). Estrategias para la Restauración del Bosque Altoandino: El Caso de la Reserva Forestal de Cogua (Cundinamarca):294-306. Universidad Nacional de Colombia. Bogotá D.C. [ Links ]
27. Victoria, M., E. Osvaldo & A. Amy. 2009. Ecological consequences of a massive flowering event of bamboo (Chusquea culeou) in a temperate forest of Patagonia, Argentina. Journal of Vegetation Science 20:424–432. [ Links ]

