











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Studies on the Harpacticoida from Colombia have been historically sparse but have increased recently (e.g., Medellín-Mora & Navas 2010, Gaviria & Defaye 2012, Fuentes-Reinés & Zoppi de Roa 2013a, b, Fuentes-Reinés & Gómez 2014, Fuentes-Reinés & Suárez-Morales 2014a, b, Suárez-Morales & Fuentes-Reinés 2015a, b, Gaviria & Defaye 2015). As an ongoing effort aiming for a better knowledge of the Colombian biodiversity, biological surveys of the aquatic fauna of a costal system of northern Colombia were carried out in year 2015. These surveys yielded male and female specimens of an undescribed species of Tisbintra, and some specimens of Geehydrosoma brevipodum (Gómez, 2004), the latter known previously from the Mexican Tropical Eastern Pacific.
One of the species reported herein belongs to the family Tisbidae, currently composed of 127 valid species (Walter & Boxshall 2016), of which 17 are known from the Caribbean Sea (Suárez-Morales et al. 2006). Most species of Tisbidae are common epibenthic organisms living in costal marine ecosystems. They can be found inhabiting the sediment-water interface and on macroalgae of brackish systems (e.g. Tisbella and Tisbe) (Boxshall & Halsey 2004, Defaye & Dussart 2011), and some genera, such as Volkmannia and Bathydia, have been reported from the deep sea (Boxshall & Halsey 2004). The genus Tisbintra was erected for a single female of Tisbintra nankaurica Sewell, 1940 from Nicobar Islands. Two decades later, Ummerkutty (1960) presented the description of the female and male of a second species, T. jonesi Ummerkutty, 1960, found in marine aquarium tanks at the Central Marine Research Station (Ramanathapuram District, India), but also in plankton samples taken from the Gulf of Mannar. So far, the genus has been reported only from the Indian Ocean: Andaman Sea (Sewell 1940), Gulf of Mannar (Ummerkutty 1960) and Gazi Bay, Kenya (Demeulenaere 1991).
Based on a number of synapomorphies (Kim et al. 2014), the genus Geehydrosoma was created to accommodate its type species G. intermedia (Chislenko, 1978) (= Enhydrosoma intermedia Chislenko, 1978) known from Posyet Bay and Ryanda Bay, Russia and Gwangyang Bay, Korea (Kim et al. 2014), and G. brevipodum (= Enhydrosoma brevipodum Gómez, 2004) known from the Mexican Eastern Tropical Pacific (Gómez 2004). This is the first record of Tisbintra and Geehydrosoma for Colombian coastal systems, and constitutes also a range extension for G. brevipodum.
MATERIAL AND METHODS
Biological samples containing both species were taken from mangrove ecosystems and at oyster banks in Rodadero Bay, Magdalena, northern Colombia (11°14' North, 74°12' West) from August, 2015 to June, 2016. Water salinity, pH, and temperature were measured with a Multi 350i handheld meter (Wissenschaftliche-Technische Werkstätten GmbH, Weilheim, Germany). Water samples were collected manually using a 25 l bucket, sieved with a 45 urn zooplankton net, and preserved in 70% ethanol. Copepods were sorted manually and processed for taxonomical identification. Observations and drawings of the new species presented herein were made from whole and dissected specimens mounted in lactophenol under a Leica DMLB compound microscope equipped with phase contrast and a drawing tube at a magnification of 1000X. The material examined was deposited in the Copepoda collection of the Instituto de Ciencias del Mar y Limnología, Mazatlán Marine Station (ICML-EMUCOP). Morphological terminology follows Huys & Boxshall (1991). The following abbreviations are used throughout the text and figures: P1-P4ENP/EXP1-3, first (proximal) to third (distal) endopodal/exopodal segment of first to fourth leg; P5EXP/BENP, exopod/ baseoendopod of fifth leg; P6, sixth leg.
RESULTS
Taxonomy
Order Harpacticoida Sars, 1903
Family Tisbidae Stebbing, 1910
Subfamily Tisbinae Stebbing, 1910
Tisbintra Sewell, 1940
Tisbintra monroyi sp. nov.
(Figs. 1-8)
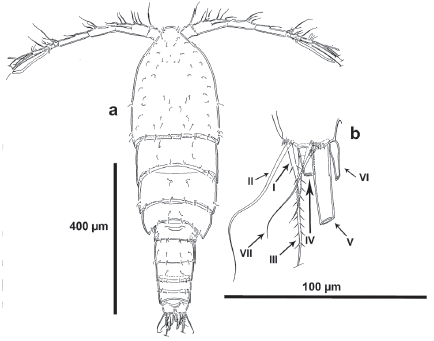
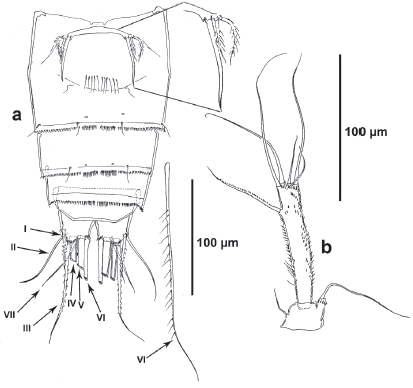
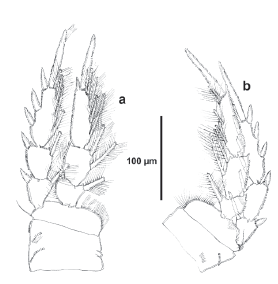
Figure 2 Tisbintra monroyi sp. nov., female. a. urosome, ventral, P5 bearing-somite omitted, showing caudal setae I-VII, b. P5, anterior.

Type material. Adult female holotype (ICML-EMUCOP-010815-06), male allotype (ICML-EMUCOP-010815-07), three females (ICML-EMUCOP-010815-02, IC- ML-EMUCOP-010815-03, ICML-EMU-COP-010815-04) and one male (ICML-EMUCOP-010815-05) dissected paratypes, and three male and five female paratypes (ICML-EMUCOP-010815-08) preserved in alcohol; Colombia, Magdalena, Rodadero Bay, 11°14' North, 74°12' West; August, 2015; coll. J. M. Fuentes-Reinés.
Diagnosis
Tisbidae. Female antennule seven-segmented; male antennule haplocer, nine segmented. Antenna with basis; exopod and endopod two-segmented. Mandible with one-segmented exopod and endopod. Maxillule with endopod and exopod fused. Maxilla with syncoxal endite represented by single seta; allobasis drawn out into strong pinnate claw with one accompanying element. Maxilliped with endopodal claw accompanied by two setae. P1-P4 with three-segmented rami. P1ENP elongated, nearly twice as long as exopod. Outer elements of P1EXP3 setiform. Proximal outer spine of P2EXP3 very reduced. Female and male P5 exopod with six, endopodal lobe with one seta. Caudal rami of female and male with seven setae.
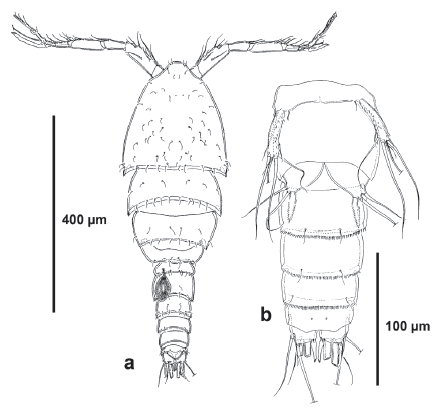
Description of female. Habitus cyclopoid in dorsal view, gradually tapering posteriorly. Total body length measured from anterior margin of rostrum to posterior margin of caudal rami ranging from 700 to 826 μm (mean, 812 urn; n, 3; holotype, 700 μm). Cephalothorax, free prosomites and first urosomite (P5 bearing-somite) without spinular ornamentation, with hyaline frill along posterior margin. Second and third urosomites (genital double-somite) separated dorsally (Fig. 1a), completely fused ventrally (Fig. 2a); first half of genital double-somite without spinular ornamentation dorsally (Fig. 1a), ventrally with transverse row of long and slender spinules medially (Fig. 2a); second half with spinular row interrupted medially close to posterior margin (Fig. 1a), ventrally with continuous longitudinal row of spinules close to posterior margin (with two sets of longer spinules medially) (Fig. 2a). Fourth urosomite ornamented as second half of genital double-somite. Fifth urosomite without spinules dorsally, ventrally as in preceding somite. Anal somite small, seemingly without ornamentation ventrally, with rounded anal operculum. Caudal rami (Figs. 1a, b, 2a) about as long as wide, with seven elements inserted distally as follows: seta I ventral to seta II, the latter nearly twice as long as the former; seta III ventral to seta I, almost aligned to seta IV; setae IV and V longest; seta VI arising on distal inner corner; dorsal seta VII biarticulated.
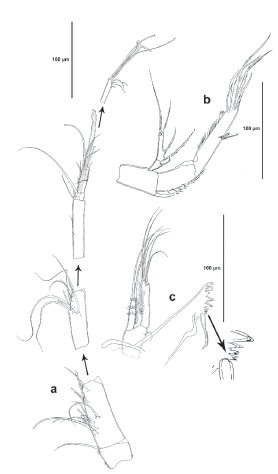
Antennule (Fig. 3a). Seven-segmented; segments 2, 3 and 4 longest. Armature formula as follows: 1(1); 2(14); 3(9); 4(2+(1+ae)); 5(2); 6(8); 7(5+(2+ae)).
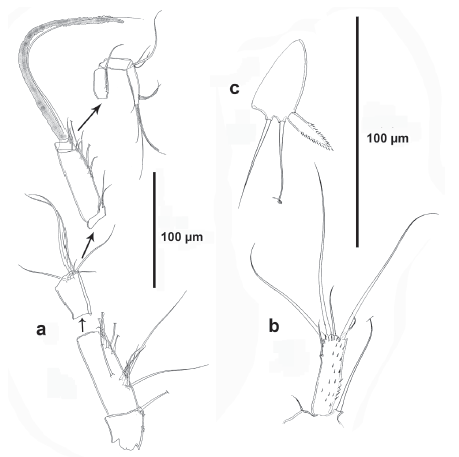
Antenna (Fig. 3b). Basis with one abexopodal long seta. Exopod two-segmented; first segment with one, second segment with two lateral and three apical setae (one of them minute). Endopod two-segmented; first segment with one inner seta; second segment elongated, as long as basis and first endopodal segment combined, outer margin with longitudinal row of spinules and with subdistal hyaline frill, with three inner lateral spines, and six apical elements as shown.
Mandible (Fig. 3c). Coxa well-developed. Gnathobase with bicuspidate teeth and with short row of small spinules, two spines and one pinnate long seta. Basis with one slender small seta. Exopod onesegmented, ornamented with spinules as shown, with two subequal apical setae. Endopod elongated, with longitudinal rows of spinules, with three inner subdistal setae (two of them fused), and two sets of two fused setae apically.
Maxillule (Fig. 4a). Elongated, with some long spinules proximally, and with row of small spinules medially. Arthrite of praecoxa with two surface setae, one pinnate lateral seta, one long posterior seta, two slender anterior setae, and three strong apical spines. Coxal endite, endopod and exopod fused, seemingly without trace of division making homologation of setae difficult to define, but most likely as follows: coxal endite and exopod represented by one well-developed seta each, endopod represented by three setae (two of them fused), basis with three sets of two fused setae each.
Maxilla (Fig. 4b). Syncoxal endite represented by one seta. Allobasis drawn out into strong pinnate claw with one accompanying element.
Maxilliped (Fig. 4c). Coxa unornamented. Basis with a row of spinules. Endopod one-segmented, with long pinnate claw, one posterior and one anterior seta.
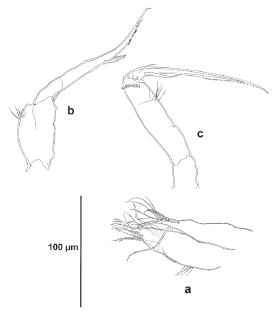
P1 (Fig. 5a). Coxa robust, ornamented with several short spinular rows as shown. Basis robust, with minute spinules at the base of the exopod and endopod, with comparatively stronger spinules at base of inner spine, outer element setiform, inner margin rounded and ornamented with long and slender spinules. Endopod three-segmented; first segment about 2.5 times as long as wide measured at its widest part, reaching middle of EXP3, segmented; first segment without inner armature, outer spine very long, nearly as long as EXP1 and EXP2 combined, rather setiform; second segment robust, with inner seta and outer normal spine; third segment with six elements, outermost element reduced.
P2 (Fig. 5b). Coxa with rows of small spinules as shown, with posterior set of slender long spinules. Basis with spinules at base of exopod, between rami, and close to inner margin; inner margin rounded and ornamented with long and slender spinules. Endopod three-segmented, longer than exopod; first segment with one, second segment with two inner setae, third segment with two inner setae, and two setae and one spine apically; outer apical corner of segments produced into acute projection (of third segment longest). Exopod threesegmented; first and second segments with outer spine and one inner seta; third segment with two inner setae, one seta and one spine apically and three outer spines, of which proximalmost very reduced.
P3 (Fig. 6a). Coxa and basis as in P2 except for lack of spinules on inner rounded expansion of basis and between rami. Endopod and exopod subequal. Endopod as in P2, except for four instead of three inner setae on ENP3. Exopod as in P2 except for three instead of two inner setae on EXP3, and for well-developed proximal outer spine.
P4 (Fig. 6b). Coxa and basis as in P3. Endopod longer than exopod, EXP3 reaching middle of ENP3. Endopod as in P2. Exopod as in P3.
P5 (Fig. 2b). Baseoendopod squarish, with long outer seta, inner endopodal lobe with one seta. Exopod elongated, about six times as long as wide, ornamented with spinules as shown, with one inner and five apical setae (one of them arising posteriorly).
P6 (Fig. 2a). Represented by elongate plate on anterior part of first half of genital double-somite, represented by one inner long spiniform element and two small pinnate setae.
Description of male. Habitus (Fig. 7a) as in female except for separated second and third urosomites in the male. Total body length measured from anterior margin of rostrum to posterior margin of caudal rami ranging from 552 to 617 μm (mean, 596 μm; n, 4; allotype, 604 μm). Cephalothorax, prosomites and fifth pedigerous somite without spinular ornamentation. Second urosomite (P6 bearing-somite) with lateral row of small spinules, without spinules ventrally and dorsally (Fig. 7a, b), with large spermathophore on the left side, conferring to the somite a globulous appearance on this side (Fig. 7a). Third and fourth urosomites with medially interrupted spinular row dorsally (Fig. 7a), continuous ventrally (Fig. 7b). Fifth urosomite without spinules dorsally (Fig. 7a), with continuous row of spinules ventrally (Fig. 7b). Anal somite and caudal rami (Fig. 7a, b) as in female.
Antennule (Fig. 8a). Nine-segmented, haplocer. Armature formula as follows 1(1); 2(14); 3(7); 4(2); 5(6+(1+ae)); 6(2); 7(2); 8(1); 9(6+(2+ae)).
Antenna, mandible, maxillule, maxilla and maxilliped, and P1-P4. As in female (not shown).
P5 (Fig. 8b). General shape as in female, but about 3.5 as long as wide.
P6 (Fig. 7b, 8c). Each leg represented by large ovoid plate, with one inner bipinnate strong spine, one apical and one outer seta.
Etymology. The species is named after Dr. José Daniel Monroy González (Bogotá) in recognition to his work on zooplankton from Colombian systems, and for his legacy and leadership of new generations of Colombian planktologists.
Remarks
Sewell (1940) created the genus Tisbintra to allocate a single female of his newly found T. nankaurica from the Nicobar Islands (Bay of Bengal and Andaman Sea). Two decades later, Ummerkutty (1960) described a second species, T. jonesi found in the Gulf of Mannar (southern India and western Sri Lanka). At this point, the known distribution of the genus was restricted to the Bay of Bengal but, in an unpublished postgraduate dissertation, Demeulenaere (1991) reported two females of T. nankaurica in samples taken from Gazi Bay (Kenya), extending the distribution of the genus to the Indian Ocean. Demeulenaere (1991) noted in the material of T. nankaurica from Kenya that: (1) the first and the second exopodal segments of the antenna possess one and four setae, respectively, instead of two and three setae as in Sewell's (1940) description, (2) the first exopodal segment of P1, not the second as in Sewell's (1940) description, bears a very long outer element, and that the third exopodal segment bears five instead of seven setae as in Sewell (1940), (3) the exopod of P5 bears six setae instead of four as in Sewell's (1940) original description of the species, and (4) the three-segmented condition of the P1ENP, instead of two-segmented as in Sewell's (1940) account. On the other hand, Demeulenaere (1991) confirmed the eight-segmented condition of the female antennule, the presence of one element only on the female P6, and the lack of armature on the endopodal lobe of the female P5. Ummerkutty (1960: 154) presented some differences between T. nankaurica and his T. jonesi. However, these differences were based on the erroneous description by Sewell (1940) and should be taken cautiously for species delineation. Sewell's (1940) and Ummerkutty's (1960) descriptions of the armature complement of the mandible, maxillule, maxilla and maxilliped lack the necessary detail and comparison with the new species presented herein could cause confusion. Also, note that Sewell (1940) and Ummerkutty (1960) described the P1ENP as two-segmented, and that Demeulenaere (1991) described this ramus as three-segmented in T. nankaurica. This is the condition for the new species described herein, and is probably the right condition in T. jonesi. Ummerkutty (1960) described the female P5EXP with five setae only, but this might be erroneous and the posterior small seta could have been overlooked. At this point, it seems that the only reliable differences between T. nankaurica and T. jonesi are the number of setae/spines on the P1EXP3, with five elements in T. nankaurica if correct, but six in T. jonesi, and the number of setae on the female P5EXP, with six setae in T. nankaurica, but five in T. jonesi if correct.
Should the armature complement of P1 EXP3 of T. nankaurica and of the female P5EXP of T. jonesi prove to be incorrect, and the presence of six setae on each ramus is confirmed, T. jonesi could be a synonym of T. nankaurica, but this requires confirmation.
The new species presented herein was attributed to the genus Tisbintra based on the combination of the following characters: exopod of the antenna two-segmented with one and five setae on the first and second segment, respectively, P1EXP1 with outer element very long, P1ENP three-segmented, P2-P4 with three-segmented rami, P2-P4ENP2 with two inner setae, P2EXP3 with proximal outer spine very reduced, female and male P5 with one seta only on the endopodal lobe, and female P6 with at least one long, spiniform element.
The relationships between the three species of Tisbintra remain obscure. However, given the present evidence, the new species, T. monroyi sp. nov., can be separated from the other two species by the number of segments of the female antennule, eight-segmented in T. nankaurica and T. jonesi, but clearly seven-segmented in T. monroyi sp. nov.; by the number of segments of the male antennule, seven-segmented in T. jonesi, but nine-segmented in T. monroyi sp. nov.; by the relative length of the setae on the P5EXP, subequal in T. nankaurica and T. jonesi, but of different lengths in the new species; and by the number of setae/spines in the female P6, with only one strong element in T. nankaurica and T. jonesi, but with one inner strong spine and two small slender outer setae in the new species.
Given all the above, an amended diagnosis for the genus Tisbintra is presented:
Amended diagnosis. Tisbidae; body cyclopiform. Rostrum fused to cephalothorax. Female antennule seven- or eight-segmented; male antennule haplocer, seven- or nine-segmented. Antenna with basis; exopod two-segmented, first segment with one, second segment with five setae. Mandible with one-segmented endopod and exopod. Maxillule elongated; coxa, basis, endopod and exopod fused. Maxilla as for the family, syncoxal endite represented by single seta. Maxilliped subchelate, basis without armature, endopodal claw with two accompanying setae. P1 with threesegmented rami; outer spine of EXP1 very long, of EXP2 normal; EXP3 with five(?) or six setae/spines; ENP1 and ENP2 with one inner seta; ENP3 reduced, with three setae. P2-P4 with three-segmented rami; P2-P4ENP1 with one, ENP2 with two inner setae; P2-P4ENP3 with five, six and five setae/spines; P2-P4EXP1 and EXP2 with one inner seta; P2-P4EXP3 with seven, eight and eight setae/spines; proximal outer spine of P2EXP3 strongly reduced. Female and male P5EXP elongated, with 5(?) or six setae; endopodal lobe with one seta only. Female P6 with one strong element only, or with one inner strong element and two outer slender setae; male P6 with one inner strong spine and two slender outer setae.
Caudal rami short, with seven setae. Sexual dimorphism expressed in the genital doublesomite, antennule and P6, and to a lesser extent in the P5 and spinular ornamentation of urosomites ventrally.
Distribution and ecology. Tisbintra monroyi sp. nov. is currently known from a single locality only, Rodadero Bay, on the Caribbean coast of Colombia. It was found around of mangrove ecosystem with a small oyster bank, at 0.70 m depth where water temperature varies over the seasons from 30 to 32 °C, water salinity is 36.1 psu, and pH 8.3.
Family Cletodidae T. Scott, 1904 sensu Por (1986)
Geehydrosoma Kim, Trebukhova, Lee & Karanovic, 2014
Geehydrosoma brevipodum (Gómez, 2004) Syn. Enhydrosoma brevipodum Gómez, 2004
Material examined. One dissected adult female (ICML-EMUCOP-010815-10), and two adult males and three adult females preserved in alcohol (ICML-EMUCOP-010815-09); Colombia, Magdalena, Rodadero Bay, 11°14' North, 74°12' West; August, 2015; coll. J. M. Fuentes-Reinés.
Distribution. Mexico: Urías system, Sinaloa state, north-western Mexico (Gómez 2004); Colombia: Rodadero Bay, Magdalena.
Remarks
Gómez (2004) described E. brevipodum from an estuarine system in southern Sinaloa state, north-western Mexico. Following Gómez (2004), his newly described species belonged to Gee's (1994) type one of Enhydrosoma, characterized, among other things, by the lack of sexual dimorphism in the male P3ENP. Interestingly, Gómez (2004) did not detect any variability in the length:width ratio of the female caudal rami, and was erroneously estimated by him to be about 8.3 times. Re-examination of the type material of G. brevipodum revealed that the length:width ratio of the female caudal rami varies from 5.5 in the holotype (ICML-EMUCOP-090301-73) to about 6.9 in the paratype (ICML-EMUCOP-090301-61).
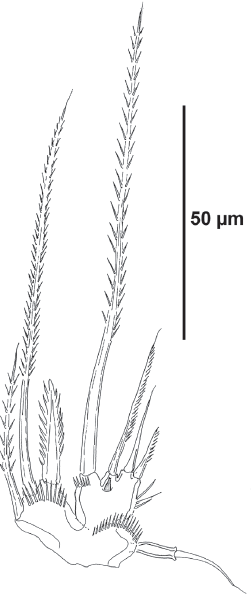
The length:width ratio of the female caudal rami of the Colombian population was observed to vary from 5.8 to 8.3. Kim et al. (2014) reallocated E. brevipodum and E. intermedia, the later known from Posyet Bay and Ryanda Bay, Russia, and Gwangyang Bay, Korea, in their newly created genus Geehydrosoma, primarily characterized by the reduced female P5EXP (Fig. 9). The distribution of the genus Geehydrosoma extends from Gwangyang Bay, South Korea, and Posyet Bay and Ryanda Bay, Russia, in the Japan Sea (G. intermedia) (Kim et al. 2014), to northwestern Mexico in the Eastern Tropical Pacific (Gómez 2004), and the Caribbean coast of Colombia (G. brevipodum, present study).

