











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Predicciones del clima terrestre afirman que la tasa de calentamiento en la troposfera inferior incrementa con la altitud debido a la doble concentración de dióxido de carbono preindustrial (CO2) (Bradley et al. 2006). Lo cual causa un incremento más pronunciado en la temperatura de alta montaña con relación a elevaciones más bajas, convirtiendo así este fenómeno climático en una amenaza directa para ecosistemas de mayor altitud. Las altas elevaciones son áreas de interés primario porque proporcionan desde un 90 a 100% de los recursos de agua potable, riego y suministro industrial en las zonas áridas y semiáridas, y además porque son focos de biodiversidad (Sánchez et al. 1989, Bradley et al. 2006, Ruiz et al. 2008).
Uno de los ecosistemas de alta elevación que ha sido alterado en una escala global por más de 300.000 km2, son las turberas a causa de prácticas agrícolas, alta deforestación comercial y explotación directa para combustibles o medios horticulturales (Charman 2002). Las turberas son ecosistemas de humedal cargados generalmente por agua y acumuladores de turba. Estos a su vez son una gran reserva y sumideros de carbono orgánico. Dichos sistemas naturales cubren más de 4.4 millones de km2 (3%) del área terrestre y ocurren principalmente en zonas subárticas y boreales (Wieder et al. 2006). El contenido de carbón de las turbas es aproximadamente 500 Pg lo que corresponde a un tercio del carbono del suelo del planeta (Gorham 1991). Esta gran cantidad de carbono almacenado en las turberas indica el gran impacto que tienen estos ecosistemas sobre el ciclo global del carbono (Yu et al. 2011).
Actualmente se conoce que el 11% de las turberas ocurre en los trópicos y almacenan allí una gran proporción del carbono en suelos de turba en el mundo (Page et al. 2011). A pesar de sus tamaños relativamente pequeños, las turberas en los Andes tienen un efecto desproporcionado en la regulación del agua porque son reservorios de suelos saturados y liberan agua que se utiliza más adelante en las actividades humanas (Holden 2005, Jauhiainen et al. 2008). El área global de las turberas se ha visto reducida significativamente durante los últimos cientos de años, resultando en una degradación severa de la función de los ecosistemas y en consecuencias ambientales y económicas adversas (Turetsky et al. 2002).
Las turberas son sistemas sensibles a cambios hidrológicos, climáticos y desequilibrio de sustratos. Dada una alteración en dichas condiciones ambientales, se transforma este ecosistema de un sumidero a una fuente de carbono y, posiblemente a uno de los mayores emisores de metano (CH4) (Jones & Yu 2010). Además, dichas condiciones ambientales afectan procesos como la distribución, composición y crecimiento de la vegetación en condiciones específicas a un sitio (Wieder et al. 2006). Las turberas se caracterizan por su habilidad única de construir y almacenar por milenios los remanentes muertos de plantas como musgos, juncos, pastos y árboles (Charman 2002), lo que permite observar un registro histórico de cambios establecidos como sucesiones vegetales. Asimismo, en estos humedales la producción primaria neta excede la tasa de descomposición durante cientos de años y como resultado de esto se da una materia orgánica rica en carbono (Clymo 1984).
Con este estudio buscamos entender las respuestas que las turberas de alta elevación tienen frente a extremos cambios climáticos. Nuestro estudio se basa en los cambios en la vegetación autóctona de turberas dominadas por Distichia muscoides Nees & Meyen en la Sierra Nevada del Cocuy (Colombia). Por tanto, queremos relacionar la ocurrencia de dicha vegetación con la variación climática de temperatura y precipitación durante los últimos 200 años. Nuestro objetivo más amplio es entender los factores climáticos que controlan los procesos de acumulación o atrición de turba a altas elevaciones para poder predecir los efectos que el cambio climático global tendrá sobre estos ecosistemas.
MATERIALES Y MÉTODOS
Área de estudio
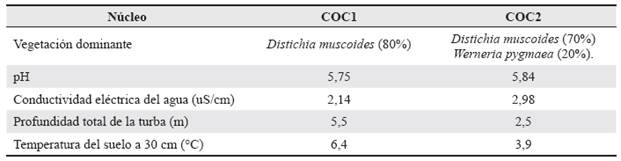
Los núcleos de turba se extrajeron en el año 2010 en el Parque Nacional Natural El Cocuy, que protege a la Sierra Nevada del Cocuy, situada al noroeste de los departamentos de Boyacá y Arauca, en el norte de la Cordillera Oriental de los Andes colombianos, aproximadamente a los 6° 27' N, 72° 17' O. Específicamente, el núcleo de turbera Cocuy 1 (COCI) fue obtenido en el "Valle de los Cojines" en 6°28'37.79" N, 72°16'42.14" O a 4202 metros de elevación sobre el nivel del mar, tomado del sector centro oriental del humedal el cual tiene un tamaño aproximado de 80 ha. El núcleo de turbera Cocuy 2 (COC2) se adquirió a la orilla del lago "Los Patos" a 6°29'52.02" N, 72°17'20.34" O a 4600 m (Fig. 1). La turbera donde se tomó el núcleo COC2 se encuentra en una terraza glaciar justo encima del Valle de los Cojines a menos de 1500 m lineales del núcleo COC1. La vegetación dominante en ambos sitios es Distichia muscoides (Juncaceae) con individuos dispersos de Senecio formosoides Cuatrec. (Asteraceae) y Werneria pygmaea Gillies ex Hook. & Arn. (Fig. 2). En el sitio COCI D. muscoides tiene una cobertura de aproximadamente el 80%, seguida por Werneria pygmaea con una cobertura del 10%, individuos dispersos de S. formosoides (<2%) y musgos de los géneros Campylopus y Breutelia (ca 5%). El sitio COC2 está dominado por D. muscoides (60%) seguido por W. pygmeae (30%), musgos del género Campylous (ca 5%) y unos pocos individuos de S. formosoides (<1%). El pH de ambos sitios es levemente ácido con valores por encima de 5 y con una conductividad eléctrica baja menor a 3 μS cm-1 (Tabla 1).
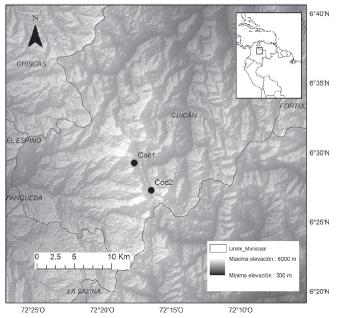
Figura 1 Mapa de la zona de estudio con los sitios de los dos núcleos marcados por círculos negros. El área de estudio se encuentra ubicada en la parte Oriental de la Sierra Nevada del Cocuy (Colombia).

Figura 2 Fotografías de las áreas donde se colectaron los núcleos en el PNN Cocuy. a. corresponde al núcleo COC1 en el Valle de los Cojines y b. corresponde al núcleo COC2 en la laguna de Los Patos (ver Tabla 1).
Cada núcleo fue extraído utilizando un tubo de PVC con puntas en los bordes, con 60 cm de largo y 10 cm de diámetro. Los núcleos comprenden 50 cm de turba. Al retirar el tubo se usó una tapa de PVC en la parte inferior ajustada manualmente. Las muestras se mantuvieron rígidas durante el traslado hasta el laboratorio debido a la dureza de D. muscoides. Los núcleos se congelaron por 48 horas para evitar la compresión de la turba. Para extraer los núcleos del molde se descongelaron por algunos minutos y seguidamente se fraccionan en fragmentos de 2 cm de espesor. Todas las muestras se secaron en el horno a 75°C durante 48 horas hasta que no hubo cambio en el peso. El extraer un solo núcleo de cada sitio limita nuestro entendimiento de la variabilidad espacial dentro del humedal pero estudios en otras turberas han mostrado la representatividad de un solo núcleo es suficiente para describir los procesos de cambio recientes (Turetsky et al. 2000).
Revisión de las muestras
Para el análisis de los macrofósiles se usó un Estereomicroscopio Zeiss DV4 con un zoom continuo de 8x a 32x y se examinó una porción de aproximadamente 2 cm de ancho y 2 cm de largo de cada segmento de turba si se podían diferenciar restos, o la muestra completa si el segmento del núcleo presentaba un alto grado de descomposición. En cada muestra separamos y medimos el porcentaje de raíces, musgos, juncos y restos no identificables. La proporción fue establecida bajo criterio del observador.
Usamos dataciones de cronologías continuas con 210Pb. Las cronologías de 210Pb nos permiten asignar una edad absoluta a las diferentes profundidades del núcleo usando la proporción de 210Pb activo en la muestra con respecto a la actividad del 210Pb en la superficie (Appleby et al. 1997). Las raíces de D. muscoides pueden intervenir en la movilización secundaria de cationes en la turba, sin embargo, evidencia de capas de ceniza proveniente de erupciones volcánicas ha corroborado de manera independiente las fechas asignadas por decaimiento radioactivo de 210Pb (Benavides et al. 2013), el cual hace parte del proceso de decaimiento del 238U. Durante el proceso de decaimiento de 238U que se encuentra de manera natural en los sustratos, se pasa por un estadio gaseoso en 226Ra el cual se distribuye en la atmósfera. El proceso de decaimiento radiactivo continua y luego de varios intermediarios de corta vida, el gas 226Ra pasa a 210Pb que se deposita inmediatamente y cubre la superficie del suelo. Este exceso de 210Pb con respecto a la concentración del substrato se denomina el exceso no soportado de 210Pb y la proporción en la cual este exceso disminuye con el tiempo es el principio de la datación. Uno de los supuestos principales del modelo es que la cantidad de 210Pb proveniente de la atmósfera es constante en el tiempo lo que permite realizar las estimaciones de la proporción del elemento que ha decaído a lo largo de la columna de turba. La concentración de 210Pb se determina disolviendo el sustrato en concentraciones de ácidos fuertes y luego agregando una superficie de cobre (2x2 cm) sobre la cual los átomos de 210Pb se adhieren. La placa de cobre con el recubrimiento de los átomos metálicos extraídos de la turba es llevada a un espectrómetro de partículas alfa ORTEC 576, el cual detecta las desintegraciones radioactivas provenientes de átomos de 210Pb en un intervalo de tiempo y con base en esto se determina la concentración de 210Pb en la muestra (Turetsky et al. 2004).
Usamos un trazador de 208Po para determinar la eficiencia de la extracción agregando una cantidad conocida de 208Po al inicio del proceso para determinar el porcentaje final de 210Pb extraído de la muestra. Las dataciones de turba usando 210Pb presentan dos fuentes de error importantes: la primera es la incertidumbre en la detección de los eventos de desintegración ya que estos ocurren a intervalos no periódicos. La segunda proviene del ajuste del modelo de entrada constante de 210Pb al sistema. Ambas fuentes de error son incluidas en el cálculo de la incertidumbre de las fechas estimadas (Appleby et al. 1997, Turetsky et al. 2004).
Las dataciones se realizaron sobre secciones circulares de 2 cm de espesor y 10 cm de diámetro. Debido a la compactación de la turba los primeros centímetros del núcleo representan intervalos de tiempo cortos (meses-años), a mayor profundidad la descomposición y compactación de la turba hace que cada sección de dos centímetros represente varias décadas. Así, la compresión de la turba causa una distribución asimétrica de la representatividad de las diferentes épocas con una alta resolución de los últimos 20 años.
Nosotros tomamos como variables climáticas de interés los registros históricos de precipitación y temperatura, para esto usamos una combinación de registros históricos de estaciones nacionales e interpolaciones globales construidas con Modelos de Circulación Globales (GCM). Tomamos los datos usando el portal de manejo de datos del Panel Intergubernamental de Cambio Climático (Intergovernmental Panel of Climate Change-IPCC "http: http://ipcc-data.org/"). Los datos de temperatura promedio anual fueron obtenidos del Climate Prediction Center (CPC "http: http://www.cpc.ncep.noaa.gov/products/precip/realtime/GIS/retro.shtml"). Datos de la precipitación anual para el periodo 1900-2010 se obtuvieron del Earth System Research Laboratory (http://climate.geog.udel.edu/~climate/html_pages/download.html#P2011rev). La resolución espacial de los datos usados fue de 0,5° por lo que ambos sitios tienen el mismo valor de temperatura y precipitación a pesar de estar a más de 500 metros de distancia horizontal y 300 metros de diferencia en elevación. La resolución temporal que usamos para los registros climáticos proviene de interpolaciones mensuales de temperatura global desde enero de 1900 hasta diciembre de 2010. Nosotros usamos datos promediados de precipitación y temperatura por década de 1900 a 1990. Los últimos 20 años de registros los analizamos anualmente para que correspondieran de manera más cercana con las dataciones obtenidas. Nuestro estudio se centra en la variabilidad temporal de la temperatura y precipitación y se debe reconocer que estos valores son promedios sobre el área cubierta.
Análisis de datos
Los cambios en la proporción de los diferentes tipos de vegetación observados en los núcleos fueron relacionados con registros históricos de precipitación y temperatura. Los registros históricos de las variables climáticas son producto de interpolaciones debido a la ausencia de registros continuos en las estaciones meteorológicas cercanas. Todos los análisis estadísticos y las figuras fueron llevados a cabo usando R x64 3.1.1 (R Core Team 2016).
RESULTADOS
Sucesión vegetal en turberas
Las velocidades de acumulación de turba se aceleran notablemente durante los últimos 20 años indicando cambios importantes en los factores que controlan la producción y la descomposición (Fig. 3). La distribución de los macrofósiles en los dos núcleos estuvo dominada por D. muscoides durante los últimos 20 años con evidencia de raíces de esta especie desde el año 1900. Los musgos diferentes a Sphagnum muestran un aumento en su porcentaje de participación dentro del núcleo durante el periodo 1980 - 2000 aproximadamente, para luego desaparecer casi completamente; aunque en el núcleo COC1 se evidencia un aumento en las tasas de acumulación durante los últimos 10 años (Fig. 3). Sphagnum fue poco abundante y solo se observaron unos pocos fragmentos de tallo y hojas de Sphagnum pylaesii Brid. el cual no es formador de turba (Fig. 4 a-d).
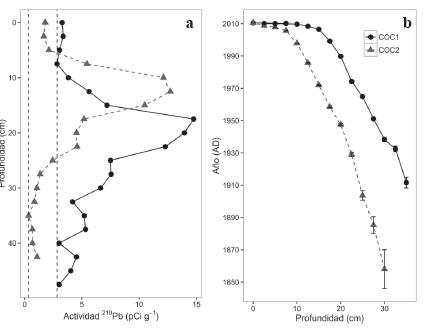
Figura 3 a. Cambio en la actividad de 210Pb con la profundidad. Las líneas punteadas verticales corresponden a los puntos límites de 210Pb soportado por el sustrato para cada uno de los núcleos. b. Curvas de acumulación de turba usando decaimiento de 210Pb. Barras de error corresponden a incertidumbre de la datación (Wieder 2001).
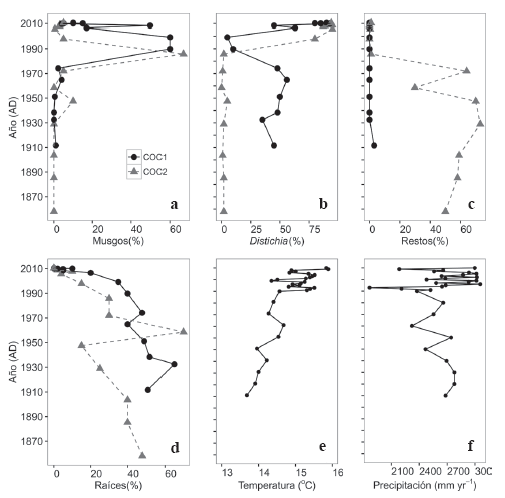
Figura 4 a-d. Distribución (en porcentaje) de los principales tipos de macrofósiles identificados en dos núcleos de 50 cm de profundidad en turberas de Distichia muscoides en la Sierra Nevada del Cocuy. Cronologías reconstruidas con 210Pb. COC1 líneas continuas y círculos, COC2 líneas punteadas y triángulos. e. Reconstrucción de la temperatura por décadas y años en la Sierra Nevada del Cocuy desde 1900 hasta 2010. f. Reconstrucción de la precipitación promedio anual por décadas y años en la Sierra Nevada del Cocuy desde 1900 hasta 2010 (detalles en el texto).
La disminución en la dominancia de D. muscoides estuvo acompañada de un aumento de materia orgánica en diferentes estados de descomposición sin una afinidad taxonómica particular o por un incremento en la abundancia de musgos diferentes a Sphagnum. Entre los musgos diferentes a Sphagnum se encontraban en orden descendente de abundancia las siguientes especies: Campylopus anderssonii (Müll. Hal.) A. Jaeger, Breutelia chrysea (Müll. Hal.) A. Jaeger, Straminergon stramineum (Dicks. ex Brid.) Hedenãs y Bryum laevigatum Hook. f. & Wilson.
Los registros para los porcentajes de D. muscoides difieren entre las dos localidades. En COC1 en el Valle de los Cojines se observan restos de D. muscoides dominando el humedal durante los últimos 100 años. En COC2 D. muscoides es dominante en los últimos 25 años. Durante el periodo comprendido entre 2000 y 2005 D. muscoides disminuyó su dominancia en COC1 y aumentó su dominancia en COC2. Entre los años 2005 y 2010 D. muscoides fue dominante en ambos sitios (Fig. 4 a-d).
Los restos vegetales no identificados ("debris") son abundantes en las partes más profundas del núcleo COC2 y por el contrario son casi asuntes en el núcleo COCI; allí por el contrario abundan los restos identificables de D. muscoides. Los restos de plantas no identificables son remanentes in situ de plantas descompuestas diferentes a D. muscoides, debido a que no se observan las vainas de las hojas que son la parte más resistentes a la descomposición del pasto (Fig. 4 a-d).
Temperatura y precipitación
Variables climáticas registradas entre el periodo de 1990 hasta el 2010 permiten evidenciar el efecto del comportamiento climático en los ecosistemas de la Sierra Nevada del Cocuy, posibilitando la reconstrucción de ambientes pasados por medio de la relación del cambio climático y las sucesiones vegetales de las turberas, al considerar la temperatura y precipitación promedio registradas para la región (Fig. 4 e-f).
En el periodo registrado (1900-2010), la temperatura de la Sierra Nevada del Cocuy presenta un incremento global de más de 1°C para los últimos cien años. Este incremento expone diversas fluctuaciones, que van desde pequeños aumentos hasta abruptas disminuciones. Los registros de precipitación para los últimos 100 años comprenden un incremento global de precipitación con un valor de aproximadamente 300 milímetros por año y un comportamiento general de extremas fluctuaciones.
Relaciones climáticas con la sucesión vegetal
El incremento global en la temperatura presenta una relación positiva para la aparición y dominancia de Distichia en cada uno de los núcleos. Hecho contrario al comportamiento encontrado para los macrofósiles de musgos y restos vegetales no identificados, los cuales se ven afectados negativamente por dicho aumento. Un caso particular, es la aparición simultánea de Distichia en los dos núcleos a partir del año 2000, hecho relacionado con el incremento en 1°C de la temperatura para los últimos diez años. La colonización y máxima aparición de musgos para el periodo comprendido desde 1970 hasta el año 1980 se encuentra asociada con un incremento en las variables climáticas de temperatura y precipitación, con un valor de aproximadamente 14.23°C y 2600 milímetros por año (Fig. 4 e-f).
DISCUSIÓN
El calentamiento dado para sitios de alta elevación resulta en un cambio detectable en la composición de especies a lo largo de los últimos 200 años (Fig. 4). Los cambios en la composición de especies en los humedales está también relacionado con los cambios en la balance hídrico local, que a su vez puede estar relacionado con la temperatura por el retroceso glaciar (Poveda & Pineda 2009). De hecho, estos ecosistemas de turbera son alimentados por los minerales desprendidos de la roca con el paso del agua, haciendo de ésta un sitio rico en nutrientes y fuentes hídricas. Por tanto, se hace abundante en Distichia muscoides (Cooper et al. 2015). La ligera acidez (pH>5.5) de las turberas de D. muscoides (Tabla 1) indica la fuerte influencia de materiales del suelo y la roca de origen sedimentario marino que son arrastrados por la lluvia o escorrentía superficial proveniente de los glaciares la cual aumenta la alcalinidad de turberas que son normalmente ácidas (Gonzalez & Flint 1965, Benavides et al. 2013).
El comportamiento de D. muscoides en "Laguna de Los Patos" ubicada a 4600 m de elevación se explica a partir del asentamiento o la formación temprana de la turbera como consecuencia del retroceso glaciar (Kuhry et al. 1993). Este fenómeno, produce fluctuaciones hídricas, las cuales son un factor altamente determinante en la formación de turberas y la colonización de plantas. Es así, como el posible desarrollo temprano de dicho humedal trae consigo una dominancia de una vegetación diferente a Distichia y el aumento en la abundancia de material vegetal no identificado para esta localidad en el periodo recuperado por el núcleo de 1900 a 1990. Los restos encontrados en la parte más antigua del núcleo COC2, muy posiblemente están asociados a la cercanía de sedimentos del fondo de un sistema lacustre o iniciación de la turbera con dominancia de plantas menos resistentes a la descomposición como Werneria pygmaea Gillies ex Hook. & Arn. (Cleef 1981). Distichia muscoides es una planta resistente a la descomposición y se puede preservar durante un largo periodo en el cual se logra percibir aún las vainas de las hojas cuando se compara con cualquier otra especie de humedal sobre los 4.000 m (Cooper et al. 2015).
Durante el periodo comprendido entre 2000 y 2005 D. muscoides desaparece en el núcleo "El Valle de los Cojines" (4300 m) y aumenta de manera considerable en "La Laguna de los Patos". Este cambio en la composición de la turba está en sincronía con el evento del Niño más intenso reportado a la fecha (Martínez et al. 2011). La sincronía entre ambos eventos indica una posible relación causa efecto entre las condiciones climáticas y el desarrollo de los humedales de alta elevación. Las sequías producidas por el evento del Niño 1997-1998 muy posiblemente tienen un efecto diferente sobre ambas localidades. La mortalidad de D. muscoides, la invasión de musgos en el "Valle de los Cojines" y un crecimiento de D. muscoides en "La Laguna de los Patos" ocurre justo después de este periodo. En el "Valle de los Cojines" la colonización de musgos diferentes a Sphagnum y el aumento de fragmentos no reconocibles de plantas indica altas tasas de mortalidad y colonización por especies de cojines más un incremento de la acidez. En tanto el rápido crecimiento de D. muscoides en "La Laguna de los Patos" indica condiciones hidrológicas estables (Benavides et al. 2013, Cooper et al. 2015). "La Laguna de los Patos" se encuentra bajo la influencia directa del cuerpo glaciar del pico Ritacuba Blanco y las altas temperaturas y radiación asociadas al Niño incrementan sus tasas de deshielo alimentando las turberas en sus laderas. Al mismo tiempo, el retroceso glaciar desconecta las turberas más alejadas como "El Valle de los Cojines" del sistema hidrológico glaciar y las hace más dependientes de la precipitación (Ponette-González et al. 2014).
Los glaciares de la Sierra Nevada del Cocuy eran relativamente extensos y constituían seis masas principales con movimiento evidente en la década 1960-1970 (Gonzalez & Flint 1965, Van der Hammen et al. 1980). La extensión de las seis masas glaciares se ha reducido y actualmente ninguna muestra evidencia de avance o acumulación neta, solo de retroceso (Poveda & Pineda 2009). Estudios palinológicos de la Laguna Ciega a 3500 m en cercanías de Güicán indican la influencia de los periodos de avance y retroceso glaciar durante el Holoceno (Van der Hammen et al. 1980). La resolución temporal de la paleoecología de Laguna Ciega es muy gruesa y los análisis empiezan por debajo de los primeros 50 cm de turba lo que limita su comparación con el presente estudio (Van der Hammen et al. 1980).
La prevalencia de los musgos en la "Laguna de los Patos" durante el periodo entre 1970 y 1980, aún con alta temperatura, puede deberse a la regulación hidrológica de la masa glaciar y a la transición de un sistema de humedal con un nivel de agua alto, evidenciado por la materia orgánica más profunda (Yu et al. 2003). La iniciación de turba requiere una combinación de características únicas como son un nivel de agua estable, una baja concentración de nutrientes y la colonización de especies formadoras de turba (Kuhry et al. 1993). La acidez creada por la colonización de musgos permite el desarrollo de una capa de vegetación que previene la descomposición y facilita la colonización posterior de D. muscoides (Kuhry 1988).
CONCLUSIONES
El incremento en la temperatura ha creado nuevas dinámicas en la ecología de las turberas (Turetsky et al. 2002, Benavides et al. 2013). El pasado climático de la Sierra Nevada del Cocuy ciertamente era más frío y la vegetación existente respondía a una conectividad hidrológica entre el sistema glaciar, la precipitación, y la escorrentía superficial como se evidencia en la estrecha relación entre desarrollo de la turbera de D. muscoides y precipitación reportado en este estudio. La intensidad de la anomalía de temperaturas del Niño tiene efectos sobre la variabilidad hidrológica que a su vez afecta el desarrollo de la vegetación y son el impulso final en el cambio en la composición y función de los ecosistemas. Se espera que la frecuencia e intensidad de periodos de anomalía de temperaturas en el pacífico se intensifiquen en los próximos años como producto del cambio climático (Camargo & Sobel 2005). El aumento en la frecuencia de eventos climáticos extremos podrá tener efectos impredecibles en los ecosistemas de alta montaña (Alley et al. 2003, Bradley et al. 2006). Nuestro estudio muestra solo una de las trayectorias posibles luego de una perturbación de alta intensidad en un sistema de humedales particulares. Nuestros resultados resaltan la importancia de entender cuáles son los factores que determinan que existan ecosistemas de turbera de alta elevación en los trópicos, la permanencia de estos sistemas en el tiempo y el control de servicios ecosistémicos importantes para la sociedad como la acumulación de carbono (Gorham 1991, Yu et al. 2011).

