











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El iniciador de los estudios fitosociológicos en Cuba fue Veroslav Samek (Samek 1969, 1973a, b), con posterioridad Borhidi (1991, 1996) realizó el estudio más completo sobre este archipiélago respecto a esta disciplina geobotánica.
Sobre Cuba Oriental la que fue considerada como una microplaca separada (Iturralde-Vinent 2007) y que tiene la mayoría del endemismo cubano (López 2005) y con formaciones vegetales particulares (Reyes 2006, 2011-2012) se han realizado posteriormente avances en los estudios fitosociológicos (Reyes 2014, Reyes & Martínez 2005, Reyes & Acosta 2003, 2010a, b, 2011, 2012, 2013, 2014). Estudios análogos con una metodología similar tienen un gran desarrollo en Colombia (Pinto-Zarate & Rangel-Ch. 2010, Rangel-Ch. & Arellano-P. 2010, entre otros).
La pluvisilva o bosque lluvioso, es el tipo de vegetación con mayor desarrollo del archipiélago cubano y sobre el mismo se han realizado muy pocos análisis fitosociológicos (Borhidi 1996), otros estudios botánicos se han realizado por Reyes & Acosta (2005), Matos (2007), Reyes (2011-2012), Martínez Quesada (2011-2012) y Martínez Quesada & Fagilde (2015).
Las pluvisilvas se presentan casi exclusivamente en la región oriental donde se encuentran las siguientes subdivisiones: pluvisilva de baja altitud sobre complejo metamórfico, pluvisilva submontana sobre complejo metamórfico, pluvisilva de baja altitud y submontana sobre ofiolitas, pluvisilva submontana sobre suelo de mal drenaje y pluvisilva montana (Reyes 2011-2012).
Las cuatro primeras solo se observan en la subregión Sagua Baracoa, dos de ellas son exclusivas del parque (Fig. 1), principalmente sobre rocas ofiolíticas y metamórficas y en suelos ferríticos y ferralíticos.
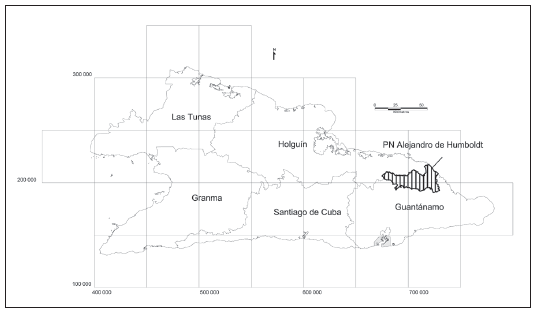
La pluvisilva es el más exuberante de los bosques cubanos y puede alcanzar hasta 45 m de altura; estructuralmente es muy compleja, pues a veces se observan hasta tres estratos arbóreos, como en la pluvisilva de baja altitud sobre complejo metamórfico de la parte media del río Jaguaní y una pequeña área de pluvisilva de baja altitud y submontana sobre ofiolitas en la parte alta de la cuenca del arroyo Angostura, muy cerca del nacimiento del río Toa.
Como ya se expuso, la fitosociología de las pluvisilvas está muy pobremente estudiada (Borhidi 1996, Reyes 2005), por ello, el objetivo de este trabajo es contribuir al conocimiento de las fitocenosis encontradas en las ofiolitas del Parque Nacional Alejandro de Humboldt.
MATERIALES Y MÉTODOS
Condiciones naturales en el área de las pluvisilvas de Sagua Baracoa.
Las zonas con pluvisilvas del macizo montañoso Sagua Baracoa (Núñez Jiménez & Viña Bayés 1989) se desarrollan desde la Sierra de Nipe al oeste, hasta el río Miel al este. Las principales rocas son las ofiolitas, principalmente ultramafitas y gabroides y las rocas metamórficas de las formaciones Santo Domingo y Sierra del Purial (Inturralde-Vinent 1990). Se encuentran también calizas fundamentalmente en las áreas periféricas.
Los suelos más extensos son los ferralíticos rojos lixiviados sobre las rocas metamórficas y los ferríticos rojo oscuros y los ferríticos amarillentos sobre las ofiolitas, también se hallan suelos pardos, sobre otras rocas.
La precipitación en el macizo Sagua Baracoa tiene características peculiares. La lluvia anual, varía desde alrededor de 1800 mm hasta poco más de 3600 mm (Montenegro 1991). El máximo absoluto en la región se produce desde las Alturas de Moa hasta el río Duaba, al este de la Planta, con más de 3000 mm; en algunos años se sobrepasan los 5000 mm. Las Sierras de Nipe y del Cristal, tienen valores anuales entre 1500 y 1600 mm al año.
Generalmente no se observan grandes diferencias en la lluvia caída entre los períodos seco, noviembre a abril y lluvioso, mayo a octubre; en la zona de máxima pluviosidad se presentan los mayores acumulados en los meses de noviembre a enero. Es común en las Sierras de Nipe y del Cristal, valores entre 120 y 150 días con lluvia al año. En las Alturas de Moa y las Cuchillas de Toa éstos aumentan, oscilando entre 150 y 180, a su vez en la zona de mayor precipitación llueve por encima de 200 días al año (Montenegro 1991).
La temperatura media anual varía entre 20 y 24°C, disminuyendo con el aumento en altitud, los menores valores se encuentran en Pico del Cristal, Pico del Toldo y Pico El Gato, todos con temperaturas medias ligeramente inferiores a los 20°C (Lapinel 1989). En general, las mínimas son en enero y las máximas en julio.
Metodología de muestreo
Los inventarios fitosociológicos con una unidad de muestreo de 25 x 25 m (625 m2) (Reyes 2005) se hicieron siguiendo los principios sigmatistas. La abundancia-dominancia de las especies, en cada estrato, se categorizó de la forma siguiente: cinco cuando cubre 75 % o más del área de la muestra; cuatro cubre 50 a 75 %; tres cubre 25 a 50 %; dos cubre 5 a 25 %; uno cubre menos del 5 %; "+" con pocos ejemplares aislados y con poca cobertura y "r" cuando eran dos o tres ejemplares aislados con poca importancia cenológica. La sociabilidad se anotó para cada especie, separada por un punto del valor de la escala combinada de abundancia-dominancia, los valores tienen los significados expuestos en Mueller-Dombois & Ellemberg (1974). Para la estructura vertical se consideraron los siguientes estratos (Samek 1973): arbóreo (E3) con más de 5 m de altura; arbustivo (E2) entre 2 y 5 m; herbáceo (E1) menos de 2 m; muscinal E0) hasta 5 cm; sinucia escandente (L- lianas) y sinusia epifítica (Ep). Se consideran subestratos cuando dentro de un estrato hay conjuntos de elementos que tienen alturas bien definidas y que son diferentes entre si.
Además, se realizaron observaciones del ecótopo en el sitio de la muestra y sus alrededores y se midieron o estimaron particularidades del suelo como tipo, profundidad, pedregosidad o gravillosidad, erosión, signos de drenaje interno, además se caracterizaron: inclinación de la pendiente, exposición, altitud (msnm), relieve en general, así como el micro y el nano relieve. La ordenación de las listas de vegetación y la separación de las fitocenosis (sin-táxones, sintaxa) se efectuó por métodos fitosociológicos (Tüxen 1937, Scamoni & Passarge 1959). Una vez organizada la tabla y unificadas las fitocenosis, se realizó la comprobación de la homogeneidad de acuerdo con la ley de Raunkiaer (1934), la que expone que en una asociación homogénea las frecuencias de las constancias, o presencias, siguen una curva típica en forma de jota invertida, o sea, la relación de las constancias sigue la fórmula: I > II > III < = > IV < V. Para la combinación característica de la asociación se emplearon las especies con grados de presencia IV y V, y para las subasociaciones y variantes las combinaciones diferenciales (Mueller-Dombois & Ellemberg 1974, Reyes & Acosta 2011, 2013). En la designación de las especies características de las alianzas se utilizaron las recomendaciones de Braun Blanquet (1951) ratificadas por Mueller Dombois & Ellemberg (1974) al determinar las especies como: absolutamente estrictas (fiel), fuertemente asociadas y favorablemente asociadas.
En la categorización y la nomenclatura de los rangos de las fitocenosis se siguió el Principio II y el Artículo 11 del Código de Nomenclatura Fitosociológica (Weber et al. 2000).
En la descripción de los estratos y sinucias se establecieron las siguientes categorías de presencia de las especies (Reyes & Acosta 2011): constantes, presentes en el 80 % o más de los inventarios; frecuentes, presentes del 60 al 79 %; menos frecuentes, presentes del 30 al 59 % y ocasionales, presentes del 15 al 29 %.
También se midió el grosor de las capas del mantillo (capa L, parte superior sin descomponer; capa F o de fermentación y capa H o húmica), se registraron las raíces y raicillas, así como las características de la estera radical (Reyes & Fornaris 2011).
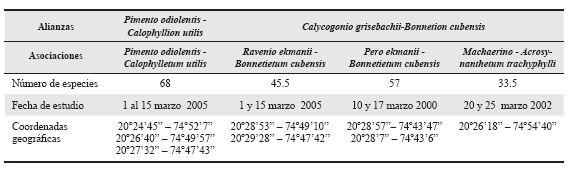
En diferentes áreas de cada formación vegetal estudiada se tomaron coordenadas, a partir de las cuales se delimitó un radio de 5 km en cuyo territorio se realizaron las unidades de muestreo con las características expuestas (Tabla 1).
Tabla 1 Coordenadas geográficas del centro de las áreas con 5 km de radio, número de especies y fecha de estudio de las pluvisilvas sobre ofiolitas en el Parque Nacional Alejandro de Humboldt.
Los nombres científicos completos, género, especie y autor, se establecen según Acevedo-Rodriguez & Strong (2012), complementado con otros estudios de la flora cubana (Mai 2005, Hagen 2007, Echavarría Cruz & Graham 2008, González Gutiérrez 2008, Mory 2010, Duno de Stefano & Angulo 2010, Rohwer 2014, Lepper & Gutierrez 2014).
RESULTADOS
Para caracterizar las dos alianzas, cuatro asociaciones y siete subasociaciones encontradas se realizaron 36 inventarios fitosociológicos, los que se integraron en las correspondientes tablas en las que se captaron las similitudes y diferencias tanto florístico - sociológicas como ecológicas. Para describir estas pluvisilvas se constituyó el siguiente arreglo fitosociológico:
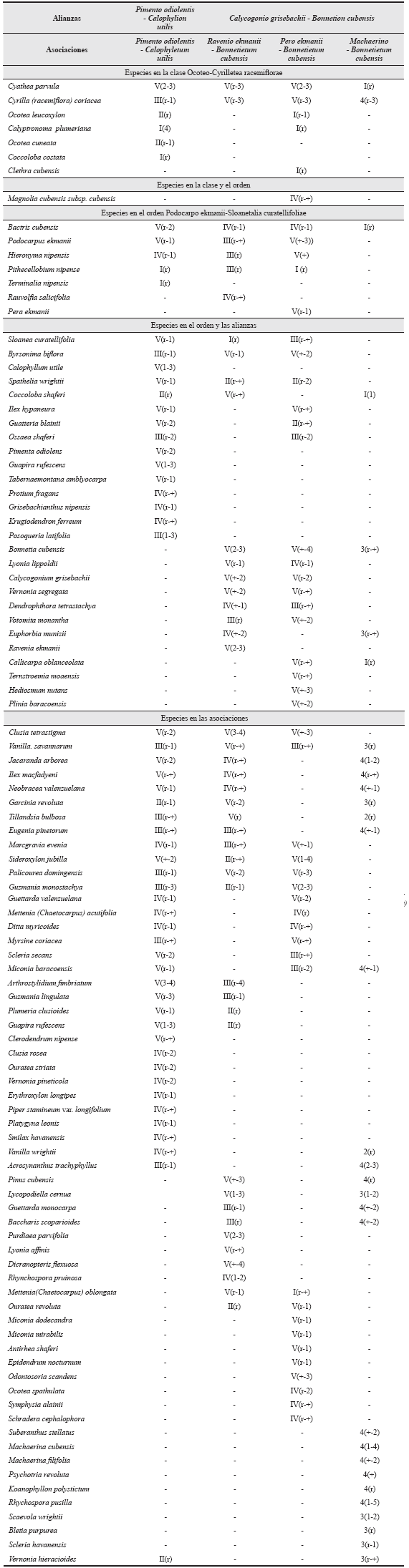
- Clase Ocoteo - Cyrilletea racemiflorae Borhidi 1996- Orden Podocarpo ekmanii - Sloanetalia curatellifoliae Borhidi & Muñíz 1996- Alianza Pimento odiolentis - Calo-phyllion utilis Reyes all. nov. En esta contribución.
Holotypus: Pimento odiolentis - Calophylletum utilis Reyes & Acosta ass. nov.
Composición florística - especies características. Absolutamente estrictas Pimenta odiolens, Buxus rotundifolia; fuertemente asociadas Protium fragans, Spathelia wrightii, Krugiodendron ferreum, Ossaea shaferi, Miconia moensis, Tabernaemontana amblyocarpa, Posoqueria latifolia; favorablemente asociadas Calophyllum utile, Guapira rufescens, Guatteria blainii, Sloanea curatellifolia, Ilex hypaneura y Grisebachianthus nipensis.
El estrato arbóreo fluctúa generalmente entre 10 y 20 m, con emergentes entre 25 y 35 m, excepcionalmente alcanza 45 m. La formación vegetal es la pluvisilva de baja altitud y submontana sobre ofiolitas (pluvisilva esclerófila), una descripción más exhaustiva se observa en la asociación correspondiente. Se presenta desde el nivel del mar hasta los 900 m snm en las Alturas de Moa, Cuchillas de Toa y fuera del Parque Nacional en la parte norte de las Cuchillas y Mesas de Baracoa-Imías, la Sierra del Cristal y la Altiplanicie de Nipe. Las rocas sobre las que se desarrollan pertenecen al complejo ofiolítico y los suelos son ferríticos rojo oscuros, muy pobres, ácidos y con un drenaje excelente. La lluvia varía entre 1700 y 3600 mm, sin época seca.
Asociaciones estudiadas:
Pimento odiolentis - Calophylletum utilis.
Buxo rotundifoliae - Calophylletum utilis (fuera de este parque nacional).
- Pimento odiolentis - Calophylletum utilis Reyes & Acosta ass. nov. En esta contribución.
Holotypus:Tabla 2, inv. 3.
Tabla 2 Pimento odiolentis-Calophylletum utilis en la pluvisilva de baja altitud y submontana sobre ofiolitas en las Alturas de Moa y Cuchillas de Toa. NE -noreste, SE - sureste, SSW - sursuroeste, NW - noroeste, NNW -nortenoroeste, ENE - estenoreste, ESE - estesureste. R: rocoso, mPp: muy poco profundo, Pp: poco profundo, Mp: medianamente profundo, P: profundo, MP: muy profundo.
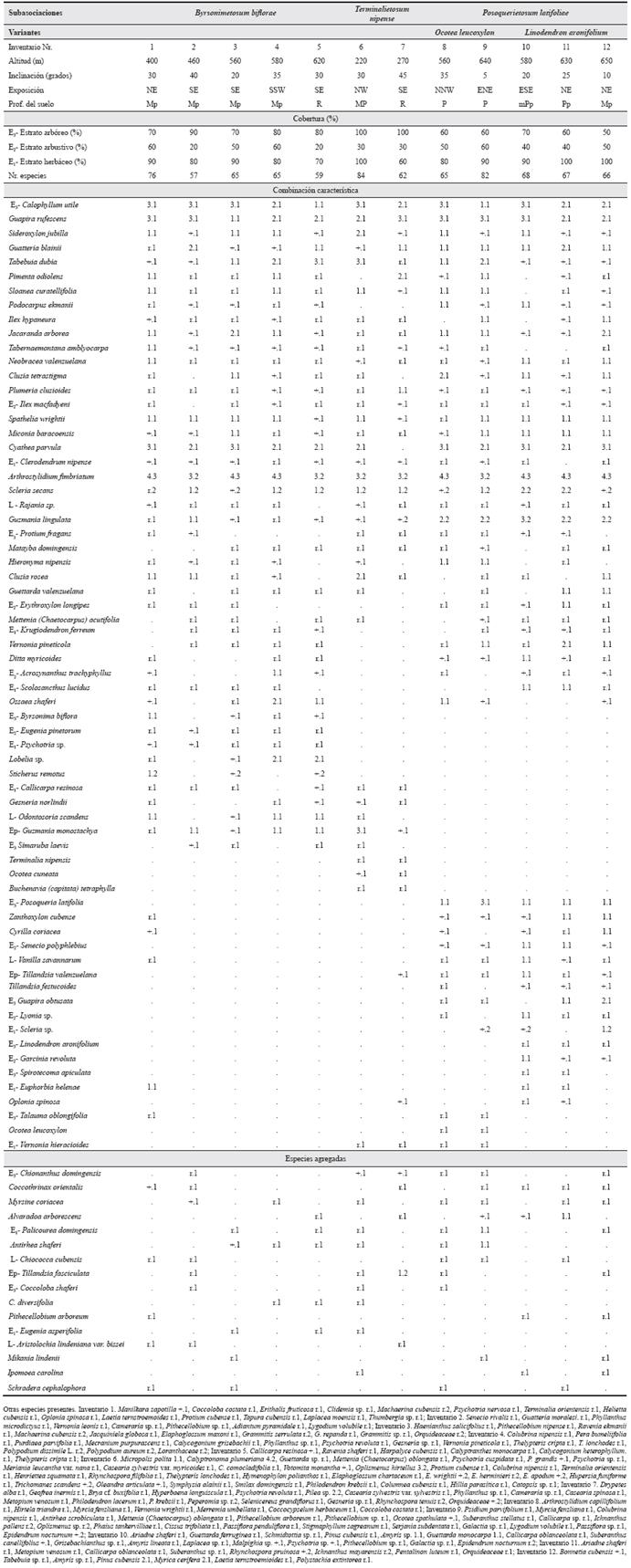
Esta pluvisilva de baja altitud y submontana sobre ofiolitas, presenta una fisionomía particular, predominando las especies esclerófilas, las hojas son notófilas y micrófilas, con un verde intenso; son frecuentes las flores rojas y las azules.
El estrato arbóreo es irregular en altura, generalmente fluctúa entre 10 y 20 m, con emergentes entre 25 y 35 m; la cobertura varía entre 60 y 100 %, excepcionalmente menos. Son constantes y abundantes Calophyllum utile Bisse, Guapira rufescens (Griseb.) Lund y Tabebuia dubia (Wr. ex Sauv.) Britt. ex Seibert.; son también constantes Sideroxylon jubilla (Ekm. ex Urb.) Gaertn., Sloanea curatellifolia Griseb., Pimenta odiolens (Urb.) Burret, Podocarpus ekmanii Urb., Tabernaemontana amblyocarpa Urb., Ilex hypaneura Loes, Guatteria blainii (Griseb.) Urb., Neobracea valenzuelana (A. Rich.) Urb., Clusia tetrastigma Vesque, Plumeria clusioides Griseb., Bactris cubensis Burret y Jacaranda arborea Urb., las dos últimas son localmente abundantes. Otras especies frecuentes, menos frecuentes y ocasionales se observan en la Tabla 2.
El estrato arbustivo es el más pobre en especies, de ellas el 53,5 % son juveniles de especies arbóreas. Su cobertura fluctúa entre 20 y 60 %. Las especies constantes son pocas, pues solo Spathelia wrightii M. Vict., Ilex macfadyeni (Walp.) Rehder, Miconia baracoensis Urb. y Cyathea párvula (Jenm.) Domin (abundante) se comportan como tal. El estrato herbáceo es denso, fluctúa entre 80 y 100 % de cobertura, ocasionalmente menos. Es relativamente rico en especies, de ellas el 32,7 % son plántulas arbóreas y el 47,2 % arbustivas. No obstante, debido a la gran constancia y cobertura de Arthrostylidium fimbriatum Griseb. y Cyathea parvula, las demás especies se hallan diseminadas; son también constantes Calophyllum utile, Spathelia wrightii, Clerodendrum nipense Urb. y Miconia baracoensis. Otras especies son observadas en la Tabla 2.
La sinusia de lianas y trepadoras es relativamente pobre en especies; solo Rajania sp. es constante. Son frecuentes Platygyna leonis Alain, Smilax havanensis Jacq., Marcgravia evenia Krug. & Urb. y Vanilla wrightii Rchb.
Como epífita constante se presenta Guzmania lingulata (L.) Mez. Son frecuentes Tillandsia valenzuelana A. Rich. y Guzmania monostachya (L.) Radlk.
Esta fitocenosis se extiende desde las Alturas de Moa hasta las Cuchillas de Toa, entre los 200 y cerca de los 900 m de altitud. La lluvia varía entre 1700 mm en Piedra la Vela y 3600 mm en La Melba, durante algunos años en esta última zona caen 5000 mm. En el primer territorio los meses más lluviosos son mayo-junio y septiembre-noviembre; en el segundo, en los meses menos lluviosos caen más de 200 mm, y los más lluviosos son mayo y de octubre a enero en que la precipitación sobrepasa los 300 mm mensuales. El macrorelieve es profundamente diseccionado, con un mesorelieve generalmente formado por pendientes abruptas, las más frecuentes son entre 20 y 35°. El suelo es ferrítico rojo oscuro, de poco profundo a muy profundo, a veces tiene algunas rocas en la superficie; es muy pobre y ácido; el drenaje es excelente, tanto interno como externo. La capa L fluctúa entre 1 y 3 cm, en ocasiones alcanza 5 cm; la F varía entre 0,5 y 3,5 cm con muchas raicillas que absorben directamente del material en descomposición. La capa H se constituye en una estera radical, embebida en una matriz de humus, entre 7 y 17 cm, más frecuentemente entre 7 y 10 cm; en ocasiones, cerca de algunos árboles llega hasta 40 cm. Dicha estera radical está integrada por una trama de raíces de diverso grosor y raicillas vitales.
Esta asociacion sigue bien la ley de Raunkiaer (1934). La combinación característica se observa en la Tabla 2; son especies características con el nivel de fuertemente asociadas: Pimenta odiolens, Tabernaemontana amblyocarpa, Protium fragans, Krugiodendron ferreum y Vernonia pineticola.
Se diferencian tres subasociaciones que son:
Pimento odiolentis - Calophylletum utilis byrsonimetosum biflorae Reyes subass. nov.
Pimento odiolentis - Calophylletum utilis terminalietosum nipensis Reyes subass. nov.
Pimento odiolentis - Calophylletum utilis posoquerietosum latifoliae Reyes subass. nov.
Las diferencias son fundamentalmente geográficas y de altitud.
La primera subasociación se estudió en las cercanias del poblado de La Melba. En los lugares cercanos a los arroyos las rocas, piedras y árboles caídos se cubren de musgos, incluso los troncos de los árboles vitales hasta cinco metros. En la combinación diferencial se observan Eugenia pinetorum Urb., Psychotria sp., Lobelia sp., Sticherus remotus (Kaulf.) Spreng. y Byrsonima biflora Griseb. (Typus: Tabla 2, inv. 3).
La segunda se presenta al oeste del poblado de La Melba. Se encuentra a menor altitud que la fitocenosis anteriormente descrita. En los lugares más húmedos, los troncos de los árboles hasta los cinco metros de altura, a veces el arbol entero, se encuentran llenos de helechos, musgos y bromeliaceas epífitas. Se caracteriza sobre todo negativamente, pues faltan muchas especies que se nombran en las otras subasociaciones. La combinación diferencial es muy pobre y forman parte de ella solamente Ocotea cuneata Griseb., Buchenavia (capitata) tetraphylla (Aubl.) R.A. Howard y Terminalia nipensis Alain (Typus: Tabla 2, inv. 6).
La tercera subasociación se observa en Piedra la Vela, en la parte suroeste de este macizo ofiolítico. La lluvia es menor que en las fitocenosis anteriores, con alrededor de 1700 mm. Es decir, se halla separada de las otras subasociaciones no solo geográficamente, sino también por la menor cantidad de lluvia. El estrato muscinal está bien desarrollado pues los musgos cubren entre 40 y 80 % de la superficie, localmente menos; las principales especies son Leucobryum giganteum Mull. Hal. y Bryohumbertia filifolia (Hornsch.) Frahm var. filifolia. Tiene una rica combinación diferencial (Typus: Tabla 2, inv. 9). Se presentan dos variantes: Linodendron aronifolium y Ocotea leucoxylon. La primera ocupa los edátopos menos profundos y tiene como combinación diferencial a Garcinia revoluta Urb., Linodendron aronifolium Griseb. y con menor presencia a Spirotecoma apiculata (Britt.) Alain, Euphorbia helenae Urb. y Oplonia spinosa (L.) Raf. La segunda se halla en suelos profundos y las especies que la separan de la otra variante las comparte generalmente con las demás subasociaciones.
- Alianza Calycogonio grisebachii -Bonnetion cubensis Reyes all. nov. En esta contribución.
Holotypus: Ravenio ekmanii - Bonnetietum cubensis Reyes & Acosta ass. nov.
Especies características: absolutamente estrictas: Bonnetia cubensis, Ravenia ekmanii, Laplacea moensis, Lyonia lippoldii, Calycogonium grisebachii, Vernonia segregata, Callicarpa oblanceolata, Hedyosmum nutans, Euphorbia munizii, Votomita monantha, Plinia baracoensis, Ternstroemia moaensis y Dendrophthora tetrastachya; favorablemente asociadas: Coccoloba shaferi y Byrsonima biflora.
El estrato arbóreo alcanza entre 20 y 25 m de altura, localmente algunos emergentes llegan hasta 30 m. La formación vegetal es la pluvisilva submontana sobre suelos de mal drenaje. Esta alianza se presenta en las altiplanicies antiguas de las Alturas de Moa. Geológicamente se corresponde con el complejo ofiolítico y los suelos son ferríticos amarillentos lixiviados, pobres, ácidos, muy plásticos y con drenaje deficiente. La lluvia es de alrededor de 3000 mm, sin período seco.
Las asociaciones que conforman esta alianza son:
Ravenio ekmanii - Bonnetietum cubensis.
Pero ekmanii-Bonnetietum cubensis.
Machaerino-Acrosynanthetum trachyphylli.
Podocarpodo ekmanii - Bonnetietum cubensisBorhidi & Muñíz 1996.
- Ravenio ekmanii-Bonnetietum cubensis Reyes & Acosta ass. nov. En esta contribución.
Holotypus:Tabla 3, inv. 3.
Tabla 3 Ravenio ekmanii-Bonnetietum cubensis en la altiplanicie entre La Melba y Moa. SE - sureste, SSE - sursureste, S - sur, WSW - oestesuroeste, W - oeste.
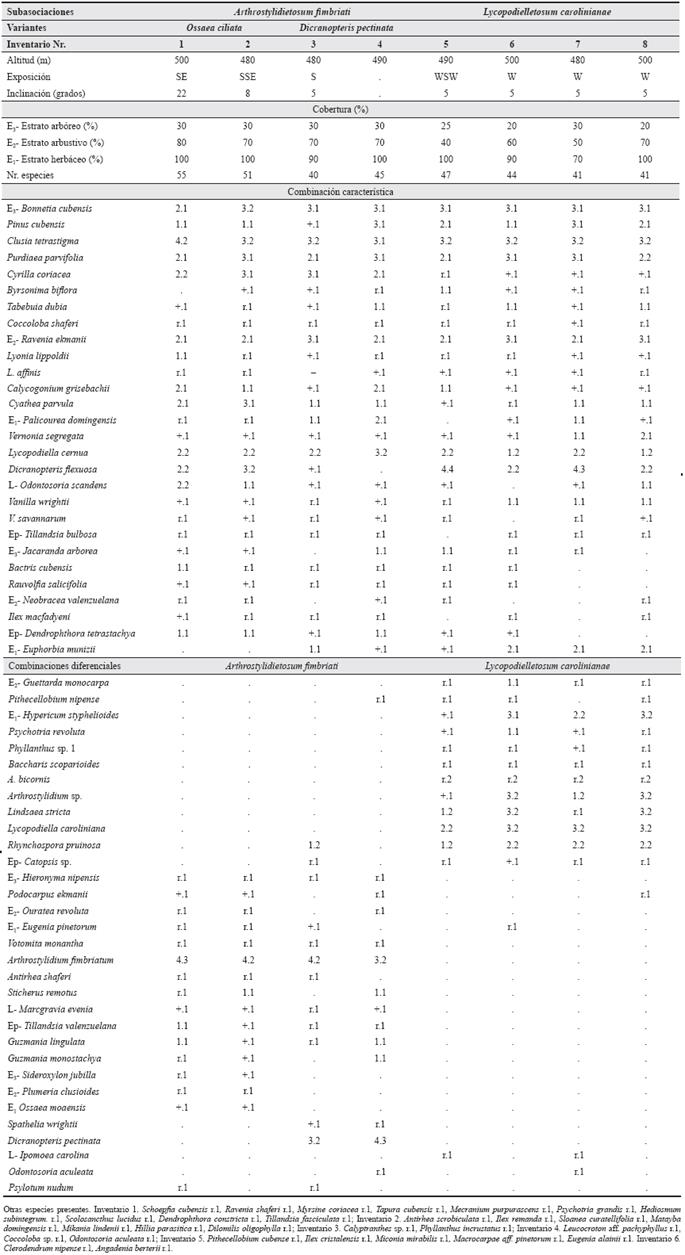
Esta conspícua fitocenosis forma parte de las pluvisilvas submontanas sobre suelos de mal drenaje y es producto de una evolución florística particular en las ofiolitas. El estrato arbóreo (E3) tiene entre 12 y 20 m de altura y una cobertura actual, debido a la explotación forestal, entre 20 y 30 %. A su vez, dicho estrato es muy pobre en especies, de éstas las constantes y más abundantes son Bonnetia cubensis (Britt.) Howard y una ecomorfosis ofiolítica del Pinus cubensis Griseb., en que las agujas son más duras y arracimadas que el fenotipo típico; son frecuentes también Byrsonima biflora y Clusia tetrastigma.
Entre los estratos arbustivo (E2) y herbáceo (E1) falta localmente una diferenciación aguda. El estrato arbustivo es denso y cubre entre 50 y 80 %, excepcionalmente menos; de las especies presentes el 42.8 % son plántulas de árboles. Son constantes y abundantes Bonnetia cubensis, Clusia tetrastigma y Purdiaea parvifolia (M. Vict.) Thomas; aunque son también constantes Garcinia revoluta Urb. y Calycogonium grisebachii Triana. Otras especies frecuentes, menos frecuentes y ocasionales se encuentran en la Tabla 3.
El estrato herbáceo presenta siempre una cobertura elevada, entre 90 y 100 %. De las especies observadas el 20,6 % son arbóreas, el 42,2 % arbustivas y el 17,2 % helechos. Las especies constantes y más abundantes son Bonnetia cubensis, Clusia tetrastigma, Lycopodiella cernua (L.) Pichi-Sermoli y Dicranopteris flexuosa (Shrad.) Und.; son también constantes Tabebuia dubia y Cyathea parvula. Cenológicamente importantes se observan en este estrato Ravenia ekmanii, Euphorbia munizii Borhidi y Calycogonium grisebachii. El estrato muscinal (Eo) cubre generalmente entre 10 y 30 % y está compuesto de musgos.
Entre las lianas y trepadoras son constantes Vanilla wrightii Reichb., V. savannarum Britt. y Odontosoria scandens (Desv.) C. Christ. Las epífitas no son abundantes, solo es constante Tillandsia bulbosa Hook y como frecuentes se observan la hemiparásita Dendrophtora tetrastachya (Griseb.) Urb. y Catopsis sp.
Ravenio ekmanii-Bonnetietum cubensis se desarrolla en las altiplanicies submontanas de las Alturas de Moa, al norte del poblado de la Melba y su altitud varía desde 400 hasta 800 m.
El microrelieve es generalmente plano, en los parteaguas secundarios la convexidad comúnmente no supera los cinco grados. La zona ocupada por esta fitocenosis se encuentra en la parte alta de las montañas que hacen elevar los vientos alisios cargados de humedad, produciendo las mayores precipitaciones del archipiélago cubano, por ello se estima que caen alrededor de 3000 mm de lluvia con una distribución regular. Además, las temperaturas son relativamente frescas, entre 20 y 22°C y las nieblas y nubes bajas cubren el área más de la mitad de los días del año. El suelo es ferrítico amarillento lixiviado, pobre, ácido, muy plástico debido a las condiciones de drenaje deficiente. En las áreas con relieve positivo se presenta erosión laminar, la que algunas veces es intensa, al igual que en zonas estrechas depresionales donde el agua superior se canaliza.
Esta asociación sigue bien la ley de Raunkiaer (1934). La combinación característica es relativamente rica (Tabla 3). En general, Bonnetia cubensis, Pinus cubensis, Clu-sia tetrastigma, Purdiaea parvifolia, Cyrilla coriacea, Ravenia ekmanii y los helechos son los más abundantes y por tanto, le imprimen a la vegetación los rasgos anteriormente expuestos. Constituyen especies características como: absolutamente estrictas Ravenia ekmanii, Purdiaea parvifolia, Lyonia affinis y como fuertemente asociadas Rhynchospora pruinosa y Rauvolfia salicifolia.
En esta fitocenosis se diferencian dos sub-asociaciones, que son:
Ravenio ekmanii - Bonnetietum cubensis arthrostylidietosum fimbriati Reyes subass. nov.
Ravenio ekmanii - Bonnetietum cubensis lycopodielletosum carolinianae Reyes subass. nov.
Entre ambas subasociaciones hay diferencias topográficas que dan lugar a disimilitudes en su funcionamiento.
La primera subasociación se encuentra en las faldas de las ligeras ondulaciones, las exposiciones son hacia el sur. La capa L tiene entre 2 y 2,5 cm y la F de 0,5 a 1 cm. La capa H, que conforma una estera radical embebida en una matriz de humus que alcanza entre 10 y 20 cm; por ello, al andar se tiene la sensación de caminar sobre un colchón. Las raíces y raicillas absorben de este material, por lo que la casi totalidad del ciclo de nutrientes se realiza en dicha estera radical. Aquí se halla la mayor cantidad de plántulas de las especies arbóreas en el estrato herbáceo, así como de arbustos y epífitas. La combinación diferencial es rica (Typus: Tabla 3, inv. 3). Arthrostylidium fimbriatum tiene una gran cobertura, por lo que también puede ser fácilmente reconocida la subasociación.
Se presentan dos variantes con pequeñas diferencias: Ossaea moaensis y Dicranopteris pectinata. Ossaea moaensis se encuentra en exposiciones hacia el sureste y las inclinaciones son poco mayores; la combinación diferencial está constituida por Sideroxylon jubilla, Plumeria clusioides y Ossaea moaensis Alain; faltan aquí Euphorbia munizii, Rhynchospora pruinosa Griseb. y Catopsis sp. que forman parte de la combinacion característica. La variante Dicranopteris pectinata que tiene como combinación diferencial a Spathelia wrightii y Dicranopteris pectinata (Willd.) Under. se halla donde la capa de hojarasca es menor, o es variable por el arrastre de las corrientes de agua.
La segunda subasociación se presenta en los parteaguas secundarios, ligeramente convexos, con microrelieve plano y pequeñas inclinaciones hacia el oeste; no obstante, se observa la erosión laminar. Debido al arrastre de la hojarasca por el agua no se establece una estera radical, sino dicha hojarasca está localizada en ocasiones debajo de Bonnetia cubensis que es el principal productor de este ecosistema. La combinación diferencial es también rica (Typus: Tabla 3, inv. 5).
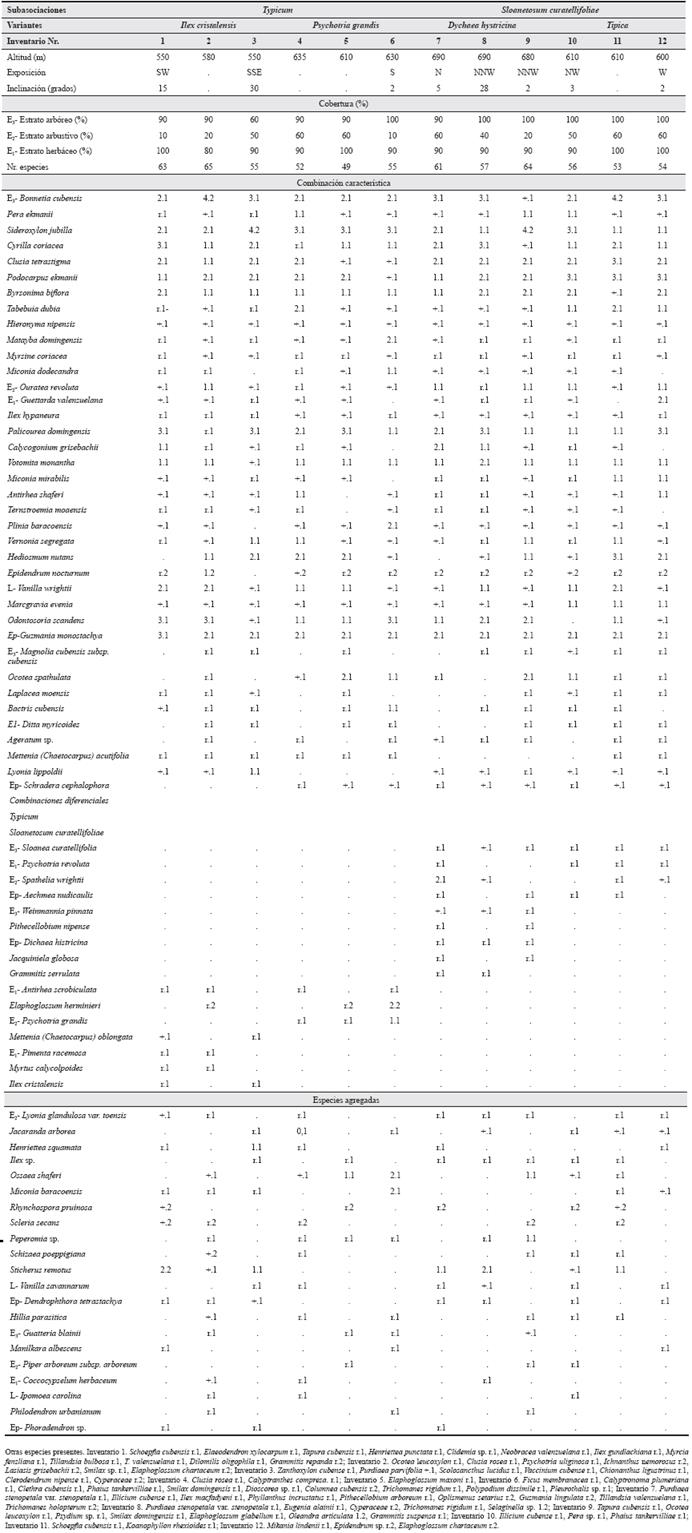
- Pero ekmanii - Bonnetietum cubensis Reyes & Acosta ass. nov. En esta contribución.
Holotypus:Tabla 4, inv. 2.
Tabla 4 Pero ekmanii-Bonnetietum cubensis en Monte Iberia. SW - suroeste, SSE - sursureste, S - sur, N - norte, NNW - nortenoroeste, NW - noroeste, W - oeste.
Esta asociación se corresponde con las pluvisilvas submontanas sobre suelos de mal drenaje. El interior de esta pluvisilva tiene características que la distingue de las demás. Generalmente todos los árboles y arbustos están cubiertos de musgos y hepáticas epifítos hasta alrededor de los 6 a 10 m de altura; a veces, los árboles están tapizados por capas de Herberthus pensilis (Tayl.) Spruce de hasta 6 a 8 cm de espesor, en ocasiones pueden llegar hasta 15 cm. En arbustos inclinados cuelgan grupos de musgos y hepáticas de hasta 10 cm de largo, principalmente de Phyllogonium fulgens (Hedw.) Brid. La superficie de los árboles en descomposición está recubierta de briófitos, y a veces con helechos epífitos, lo mismo si permanece erecto como si está en el suelo. Odontosoria scandens forma una densa red entre los árboles y sobre algunos arbustos que alcanza hasta dos metros de altura; ella forma una estructura que semeja una telaraña, e imprime (junto con los briófitos epífitos) a esta fitocenosis un aspecto particular.
El estrato arbóreo (E3) es muy denso, con una cobertura entre 90 y 100 %, excepcionalmente menos. Su altura es irregular, fluctuando entre 8 y 25 m, algunos ejemplares llegan hasta 30 m. En las fotos aéreas se observa una estructura de grano grueso, formada por los emergentes, que son principalmente de Sideroxylon jubilla. La estructura interna del bosque es generalmente continua. En ocasiones se encuentran dos subestratos, el superior de alrededor de 20 m y el inferior de 8 a 10 m. Este estrato es relativamente rico en especies. Las constantes y más abundantes, que a su vez definen la fisionomía arbórea, son Bonnetia cubensis, Sideroxylon jubilla, Cyrilla coriácea, Clusia tetrastigma, Podocorpus ekmanii y Byrsonima biflora; son también constantes Tabebuia dubia, Hieronyma nipensis Urb., Matayba domingensis (P. DC.) Radlk., Myrsine coriacea (Sw.) R. Br., Miconia dodecandra (Desv.) Cong. y Pera ekmanii Urb. (Tabla 4).
La cobertura del estrato arbustivo fluctúa entre 10 y 60 %, siendo más usuales los valores entre 40 y 60 %. De las especies presentes el 50.6 % son plántulas de especies arbóreas. Este estrato es relativamente pobre en especies constantes, pues solo Podocarpus ekmanii, Hieronyma nipensis y Cyathea parvula se comportan como tal.
El estrato herbáceo es muy denso, pues fluctúa de 90 a 100 %, excepcionalmente 80 %. Es el estrato más rico en especies, de ellas, el 28,8 % son arbóreas y el 54,6 % arbustivas. Las especies constantes y más abundantes son Podocarpus ekmanii, Clusia tetrastigma, Palicourea domingensis (Jacq.) A. DC. y Cyathea parvula; son también constantes Sideroxylon jubilla, Matayba domingensis, Hieronyma nipensis, Ouratea revoluta (Wr.) Engl., Myrsine coriacea, Psychotria sp., Vernonia segregata Gleason y Pimenta racemosa (Mill.) J.W. Moore (Tabla 4).
De todas las pluvisilvas, en esta fitocenosis es donde se observa el estrato muscinal (E0) más desarrollado, pues varía entre 20 y 80 % de la superficie, excepcionalmente menos. La especie predominante es Sphagnum meridense (Hampe) Müll. Hal., que forma manchas que parecen una alfombra sobre la hojarasca; a veces, se hallan parches de S. macrophyllum Bernh. ex Brid., S. perichaetiale Hampe, Leucobryum gigan-teum y Bryohumbertia filifolia var. filifolia.
Esta fitocenosis es relativamente pobre en lianas y trepadoras. La única constante y abundante es Odontosoria scandens, son también constantes Vanilla wrightii, localmente abundante, y Marcgravia evenia. Las epífitas y hemiparásitas son también pobres en especies. Como constante y abundante se halla solo Guzmania monos-tachya y como constante Epidendrum nocturnum Jacq. Menos frecuentemente se observan Hillia parasitica Jacq., Aechmea nudicaulis Griseb. y Dendrophthora tetrastachia.
El altiplano de Monte Iberia ocupa la parte superior del macizo ultramáfico entre los valles inferiores de los ríos Santa María y Macaguanigua al Norte y los ríos Jiguaní, Nibujón y Jaguaní al oeste, este y sur respectivamente. El relieve es ligeramente ondulado, con menos de 100 m de diferencia entre la red de drenaje y el plano principal de la altiplanicie, tiene amplias zonas llanas, donde localmente el drenaje externo es pobre. Desagua principalmente hacia el río Jiguaní; no obstante, la parte este drena hacia el río Nibujón. Monte Iberia es una montaña extraordinariamente abrupta en su vertiente norte; al entrar los vientos alisios cargados de humedad por los valles de los ríos Santa María y Nibujón, éstos se elevan brúscamente, por lo que se enfrían rápidamente, aumentando notablemente las precipitaciones. Al encontrarse esta fitocenosis en la parte superior de esta montaña, recibe alrededor de 3,000 mm regularmente distribuidos; es decir, pertenece a la zona de mayor pluviosidad del archipiélago cubano. Las temperaturas son relativamente frescas, entre 20 y 22°C. Una parte importante de los días del año están cubiertas por las nieblas y nubes bajas; además, como la humedad ambiental es muy elevada y la evaporación es pequeña, se crean condiciones ecológicas favorables para el desarrollo del epifitismo. El suelo es ferrítico amarillento lixiviado, con muy mal drenaje, tanto interno como externo, pobre y ácido, con signos de reducción en todo el perfil. En casi toda la altiplanicie se observa sobre el suelo los restos de un hardpan de perdigones consolidados, es decir, un horizonte petroférrico, el cual está desmembrado en pedazos más o menos grandes y sobre los cuales se camina. La hojarasca está muy desarrollada. La capa L alcanza entre 2,5 y 3,5 cm compuesta principalmente de hojas de Bonnetia cubensis; la F fluctúa de 2 a 3,5 cm, generalmente con raicillas absorbiendo del material en descomposición. La capa H se constituye en una estera radical, embebida en una matriz de humus, de color marrón, y varía entre 15 y 35 cm, dicha capa está completamente llena de raices y raicillas vitales. El enraizamiento de los árboles es superficial, a veces se observa que parte del mismo se encuentra superpuesto sobre el suelo.
Esta fitocenosis sigue bien la ley de Raunkiaer (1934). Las especies características como fuertemente asociadas son: Pera ekmanii, Ouratea revoluta, Callicarpa oblanceolata, Ternstroemia moaensis, Hedyosmum nutans, Miconia mirabilis y Plinia baracoensis.
Se encuentran dos subasociaciones que son:
Pero ekmanii-Bonnetietum cubensis typicum Reyes subass. nov.
Pero ekmanii-Bonnetietum cubensis sloanetosum curatellifoliae Reyes subass. nov.
Las principales diferencias son de exposición y de inclinación de la pendiente.
La subasociación typicum abarca la parte central de la altiplanicie, al norte del principal afluente del río Jiguaní en la misma. Generalmente ocupa la menor altitud y la mayor inclinación del área estudiada; la exposición es hacia el sur, por lo que recibe mayor insolación. En comparación con la otra subasociación, hay más arbustos y especies en general en el estrato herbáceo. Esta fitocenosis se caracteriza porque faltan muchas especies que serán nombradas más adelante (Typus: Tabla 4, inv. 2). Se observan dos variantes:
La primera es más común en las exposiciones hacia el sur y en las inclinaciones entre 15 y 30 grados. Se caracteriza por Mettenia (Chaetocarpus) oblongata Alain, Pimenta racemosa, Myrtus calycolpoides (Griseb.) Burret e Ilex cristalensis Loes; la segunda por Psychotria grandis Sw.
La segunda subasociación se encuentra en áreas llanas o de muy pequeña inclinación, sobre todo en exposiciones hacia el norte y en altitudes superiores a los 600 m. La combinación diferencial (Typus: Tabla 4, inv. 7) está compuesta por Sloanea curatellifolia, Psychotria revoluta Urb., Spathelia wrightii y Aechmea nudicaulis. Se presentan dos variantes con pocas diferencias florísticas y ecológicas, ellas son:
La variante Dichaea hystricina ocupa las áreas más elevadas de la asociación y en exposiciones hacia el norte. Geográficamente se ubica cerca del borde sur de la meseta. Tiene como combinación diferencial a Weinmannia pinnata L., Dichaea hystricina Rchb. y con menor constancia Pithecellobium nipense Britt., Jacquiniela globosa (Jacq.) Schltr. y Grammitis serrulata (Sw.) Sw. La variante Típica ocupa las áreas llanas o ligeramente onduladas de la parte este del territorio. En ella faltan muchas especies.
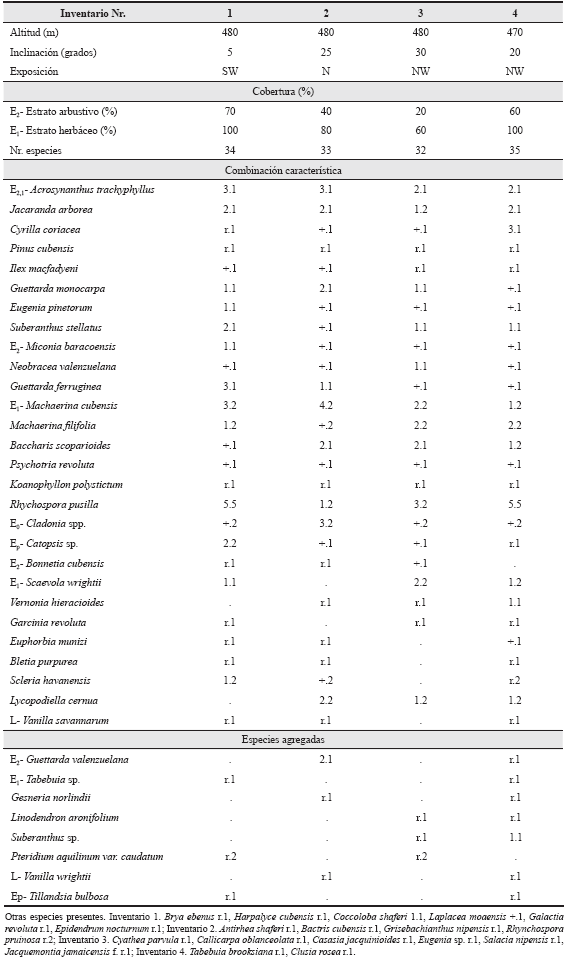
- Machaerino - Acrosynanthetum trachy-phylli Reyes & Acosta ass. nov. En esta contribución.
Holotypus:Tabla 5, inv. 4.
Tabla 5 Machaerino-Acrosynanthetum trachyphylli en la parte sur del macizo El Toldo. SW - suroeste, N - norte, NW - noroeste.
Otras especies presentes. Inventario 1. Brya ebenus r.1, Harpalyce cubensis r.1, Coccoloba shaferi 1.1, Laplacea moaensis +.1, Galactia revoluta r.1, Epidendrum nocturnum r.1; Inventario 2. Antirhea shaferi r.1, Bactris cubensis r.1, Grisebachianthus nipensis r.1, Rhynchospora pruinosa r.2; Inventario 3. Cyathea parvula r.1, Callicarpa oblanceolata r.1, Casasia jacquinioides r.1, Eugenia sp. r.1, Salacia nipensis r.1, Jacquemontia jamaicensis f. r.1; Inventario 4. Tabebuia brooksiana r.1, Clusia rosea r.1.
El estrato superior alcanza entre 3 y 6 m, con una cobertura de 20 a 70 %, principalmente más del 40 %; las especies más abundantes son Acrosynanthus trachyphyllus Standl., Jacaranda arborea y a veces Suberanthus stellatus (Griseb.) Borh. & Fernández, Guettarda monocarpa Urb., G. ferruginea Wr. ex Griseb. y es escasa G. valenzuelana A. Rich.. Frecuentemente se observan emergentes aislados, de 8 a 10 m, sin sobrepasar el 5 %; generalmente son de Pinus cubensis (ecomorfosis), Bonnetia cubensis y Cyrilla coriacea.
El estrato herbáceo cubre entre 60 y 100 % y las especies más abundantes son Rhynchospora pusilla (Sw.) Griseb., Machaerina cubensis (Kük.) T. Koyama, M. filifolia Griseb., Scaevola wrightii (Griseb.) Maza, Baccharis scoparioides Griseb., Lycopodiella cernua, Jacaranda arborea y Suberanthus stellatus (Tabla 5).
Las lianas y las epífitas son escasas, de las primeras las más frecuentes son Vanilla savannarum y V. wrightii y de las segundas Catopsis sp.
Las especies características al nivel de favorablemente asociadas son: Suberanthus stellatus, Machaerina cubensis, M. filifolia y Rhychospora pusilla.
Se desarrolla sobre rocas gabroides, pertenecientes al complejo ofiolítico, de donde se forma una corteza de meteorización amarillenta, profunda, plástica, con muy mal drenaje interno y en cuyos intersticios se mantiene el agua. El suelo es pardo grisáceo, muy poco profundo. La lluvia es alrededor de 2000 mm. La altitud es cercana a los 500 m, se presenta en la parte superior de una colina que se encuentra en el extremo sur del macizo El Toldo, en la confluencia de los ríos Jaguaní y Yarey, al norte de la loma El Mulo. Esta fitocenosis secundaria constituye un estadío sucesional avanzado. Se considera que fue un fragmento de pluvisilva submontana sobre suelos de mal drenaje que fue destruida por un huracán, observándose árboles derribados, fue además afectado por un fuego intenso. Al quedar el suelo descubierto, se erosionaron en algunas áreas más de 10 cm, formando a veces cárcavas de 30 a 40 cm de profundidad; se conservan principalmente las zonas protegidas por las plantas herbáceas que forman macolla como Rhynchospora y Machaerina.
En general, en las ofiolitas del Parque Nacional Alejandro de Humboldt se han caracterizado dos alianzas y cuatro asociaciones (Tabla 6) que refuerzan los valores del área como Patimonio de la Humanidad y área núcleo de la Reserva de la Biosfera Cuchillas del Toa. Cuando se comparan estos valores fitocenóticos con otros macizos ofiolíticos con pluvisilvas del mismo tipo se comprueba que en la Sierra de Nipe hay una alianza y dos asociaciones, en la Sierra del Cristal una de cada categoría y en la parte norte de las Alturas de Moa solo una asociación (Reyes et al., datos no publ.).
DISCUSIÓN
De los cinco tipos de pluvisilvas encontrados por Reyes (2011-2012) dos se analizan en este trabajo y tienen aquí fitocenosis únicas y muy particulares. Comparando esta unicidad fitocenótica con la de los pinares y charrascales del parque (Reyes & Acosta 2012, Capote et al. 2000) se observan en ellos igual peculiaridad sintaxonómica.
Son interesantes los inconfundibles rasgos fisionómicos de Ravenio ekmanii - Bonnetietum cubensis, fitocenosis desarrollada en suelos ferríticos amarillentos lixiviados, de mal drenaje y extremadamente pobres (Ruíz 1988), en que las hojas no forman planos horizontales sino tienen una inclinación con dicho plano de alrededor de 45 grados. Estas características ya fueron señaladas por Leigh (1975) en otros bosques montanos.
En Pero ekmanii - Bonnetietum cubensis faltan, o están muy pobremente representadas, varias especies que son comunes en las otras altiplanicies con el mismo tipo de pluvisilva. Este bosque está muy bien conservado, presentando una gran densidad en su estrato arbóreo; se considera que por su estructura y cobertura, no permitió que entraran dichas especies. Además, el desarrollo de este bosque, bajo condiciones extremas de mal drenaje, realizó una selección de las especies. Es decir, en la presencia o ausencia de las mismas actuaron factores ecológicos y de competencia; algunas de ellas son Pinus cubensis, Clusia rosea, Clerodendrum nipense, Baccharis scoparioides, Euphorbia munizii, Andropogon spp., Calyptronoma plumeriana, Philodendron lacerun, Ilex macfadyeni, entre otras.
Es también de destacar, que las plantas observadas en fitocenosis sobre ofiolitas son fundamentalmente micrófilas, esclerófilas y muchas de ellas con hojas gruesas, lo que fue anteriormente observado por Borhidi (1992, 1996). Se considera también que las condiciones edáficas tan difíciles: oligotrofía excepcional, acidez, anaerobiosis, compuestos reducidos (Ruíz 1988), en que se encuentran Ravenio ekmanii-Bonnetietum cubensis y Pero ekmanii-Bonnetietum cubensis tiene como resultado un edátopo extremo, que causa la elevada microfília, esclerofília y reciclaje de nutrientes que aquí se presenta. A su vez, la interacción de estos factores con el climátopo origina las particularidades florísticas y morfológicas de estas fitocenosis.
Respecto a la sintaxonomía en estas formaciones vegetales, Borhidi (1996) extendió la alianza Podocarpo ekmanii-Byrsonimion orientensis Borhidi & Muñíz 1996 a esta zona con la asociación Podocarpo ekmanii-Bonnetietum cubensis Borhidi & Muñíz 1996. Se considera que dicha alianza está restringida a las Sierras de Nipe y Cristal y por tanto inexistente en las Alturas de Moa y Cuchillas de Toa, donde se presenta el Parque Nacional Alejandro de Humboldt. Ambas zonas están aisladas por rocas diferentes de las ofiolitas. Como sintaxa equivalentes se describen en este territorio las alianzas Pimento odiolentis-Calophyllion utilis y Calycogonio grisebachii-Bonnetion cubensis, las que difieren fundamentalmente por el tipo de suelo y una evolución bajo condiciones óptimas de drenaje la primera y con insuficiencias en este aspecto la segunda. Por tanto, se transfiere para esta última la asociación Podocarpo ekmanii-Bonnetietum cubensis Borhidi & Muñíz 1996.
PARTICIPACIÓN DE LOS AUTORES
Ambos autores realizaron el trabajo de campo de inventario fitosociológico, determinación de las especies y valoración de los índices. FAC determinó las especies desconocidas. Ambos autores participaron en la elaboración de las tablas fitocenológicas y revisaron el trabajo elaborado. OJR redactó el trabajo.

