











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Molting, an energy demanding activity, is one of the most important processes in birds’ life cycle (Howell et al. 2003, Echeverry-Galvis and Córdoba-Córdoba 2008). Learning about plumage variation, molting, sexing and aging in birds, permit us to understand their ecology and plumage evolution. Furthermore, with long-term monitoring, they will allow a greater understanding of species demography, contributing to more effective decisions on bird conservation (Gómez et al. 2012).
Molt strategies, especially for passerines, are well documented in temperate zones (see Jenni and Winkler 1994, Pyle 1997, 2008). However, patterns present in temperate zones do not completely coincide with those of Neotropical birds, varying in duration and seasonality (Wolf 1969, Ryder and Wolfe 2009). This could be partly due to differences in resource availability throughout the year between these regions (e.g.Miller 1961, Poulin et al. 1992, Echeverry-Galvis and Hau 2013). The new W-R-P system permits classifying tropical birds into age classes (Wolfe et al. 2010, Johnson et al. 2011) and eases the understanding of plumage and molts in tropical birds and leaves an open door for ornithological research (Hernandez 2012, Johnson and Wolfe 2014). Specifically, in Colombia, research about molt and age criteria is a growing research area (e.g.Botero-Delgadillo et al. 2012, Hernández 2012, Gómez et al. 2012, Moreno-Palacios et al. 2013).
The molt of Z. capensis (Müller, 1776) has been fairly well investigated with multiple studies in Colombia and Neotropics (Miller 1961, Wolf 1969, Davis 1971, King 1972, 1973, Zerda et al. 1986, Pyle et al. 2015). Because of this, comparisons can be made among locations, latitudes and populations. In Colombia, the most detailed research about plumage, molt and breeding of Z. capensis was carried out near Cali, Valle del Cauca (Miller 1961). This study established the molt strategy, a brief description of different plumages such as juvenal, formative, basic, and estimates the duration of molt and reproduction seasons for the species.
There exist various studies about plumage, molt and breeding in Z. capensis. The existing information and its broad distribution in Central and South America make of Z. capensis an excellent model to understand the variation of duration and timing in this process in the tropical region. Z. capensis is a widely distributed species and its molt strategy varies across its distribution, even between populations of the same subspecies. In this study, I documented both, strategy and extent of molt in Z. capensis in Bogotá, describing new external characteristics useful for age determination in Bogotá, as iris color. Also, I compared my results with previous investigations and propose the hypothesis of molt variation across latitudinal gradient.
MATERIAL AND METHODS
The study area is located in the Eastern Cordillera of the Colombian Andes, in the city of Bogotá D.C. (2560 m a.s.l.). I established a total of five sampling points, two in the Bogotá Botanical Garden-José Celestino Mutis (4°40’North, 74°6’West) and three in the campus of the Universidad Nacional de Colombia (4°38’North, 74°5’West). I conducted 65 mist net samplings using 12 m nets for a total of thirteen samplings per site, between 06:00 and 11:00 h, totaling 1883.5 net hours. Samplings occurred from the 1st Apr 2013 to the 5th Apr 2014. Each site was sampled every four weeks in zones with low vegetation and grasslands.
Molt strategy and pattern
Birds were banded, allowing for identification and tracking of individuals over time. I took detailed notes about plumage, body molt, flight feathers molt, molt limits, shape of external retrices, iris color categorized following Smithe´s color guide (1975) and skull ossification (Ralph et al. 1996, Pyle 1997). Ossification was determined following the methodology of Pyle (1997) assigning a percentage of the total area of the skull. In this methodology, 0 % means a skull with a single layer of bone and 100 % means a skull with two layers and complete pneumatization. The terminology used for plumage and molt follows Humphrey and Parkes (1959) modified by Howell et al. (2003).
Ageing, molt and plumage description
The bird's age was determined with criteria such as: the presence/absence of brown streaks on the chest of juveniles (Miller 1961), molt limits, shape of flight feathers, sharp in juvenal feathers, broad in individuals with basic feathers, growth bars, aligned in juvenal flight feathers and not aligned in basic feathers (Pyle 1997). Additionally, I took notes describing other useful age characteristics of Z. capensis like iris color and skull ossification. I used the mentioned criteria to assign every bird to an age category, following the molt cycle-based age-classification system - WRP (Wolfe et al. 2010, Johnson et al. 2011).
RESULTS
Molt strategy and molt pattern
I banded 102 Rufous-collared Sparrows and recaptured 42 during the sampling year. I found that besides the basic plumages, Z. capensis counts with a formative plumage. Also, I did not find traces of alternate plumage, as it will be discussed later. Thus, the molt strategy of Z. capensis in Bogotá is a Complex Basic Strategy (CBS; sensuHowell et al. 2003). The prebasic molts present a complete molt pattern, i.e. molting all the feathers in the same molting process, and the formative plumage is the result of a partial molt in which flight feathers, primaries, secondaries, tertials, retrices and primary coverts, are not molted. Nevertheless, some captured individuals with a formative plumage had molted tertial feathers (30 %) or central retrices (17 %).
Ageing, molt and plumage description
In Bogotá, I categorized Z. capensis in eight age classes within the WRP system (Wolfe et al. 2010, Johnson et al. 2011). The first category, first-cycle prejuvenal molt (FPJ), occurs in the nest when the bird substitutes the natal down plumage for the juvenal. I did not sample nests and I do not have information about the FPJ. The first-cycle juvenal (FCJ) birds are the easiest to identify. They have a juvenal plumage which highly contrasts with posterior plumages. Juvenal plumage presents cryptic colors, brown streaks on stomach and breast and the absence of the specie's characteristic rufous collar (Fig. 1a). Moreover, 90 % of FCJ birds showed a skull ossification between 0 and 20 % and a raw amber iris color (Table 1; Fig. 2a).

Figure 1 Plumages of Rufous-collared Sparrow (Z. capensis costaricensis) in Bogotá, Colombia. a. Juvenal plumage, b. Formative plumage, and c. Basic plumage. Pictures by Diego Cueva.
Table 1 Distribution of the captured birds in the categories of iris color and skull pneumatization. Bold numbers highlight the higher percentage values at each age class. Some percentages may not reach 100 % because I could not conduct a proper inspection of some captured birds (n). i.e. molting shades may difficult the observation of the ossification.
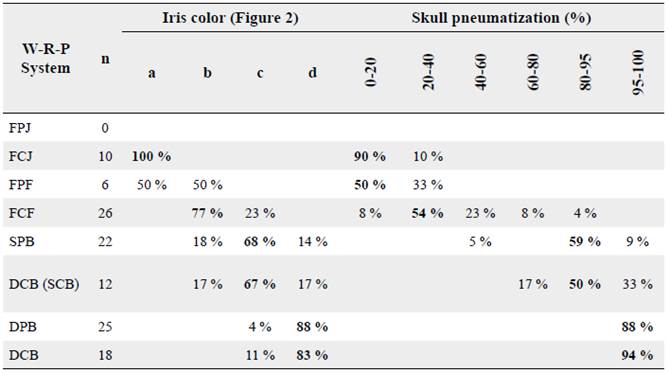

Figure 2 Iris color variation throughout different age stages. Colors according to Smithe (1975). a. Color 223 Raw umber, b. Color 32 Chestnut, c. Intermediate between 32 and 140 colors, and d. Color 140 Pratt’s Rufous. Pictures by Diego Cueva (a, b) and Julian Avila-Campos (c, d).
After FCJ, Z. capensis reaches a first-cycle preformative molt (FPF). The molt in FPF age does not include flight feathers. FPF birds presented molting body feathers, brown juvenal feathers on head and remaining streaks on chest, skull ossification under 40 % with a majority under 20 % (Table 1) and a raw amber/chestnut iris color (Fig. 2b). The acquired formative plumage named the next age class, first-cycle formative (FCF; Fig. 1b). As expected (Miller 1961, 1962), formative plumage is superficially similar to later basic plumages (Fig. 1c). Thus, body feathers became useless to classify individuals into an age class. FCF Rufous-collared Sparrow molted all secondary coverts but not primary coverts, they exhibited molt-limits between greater coverts and primaries coverts. Juvenal primary coverts had an external buffy (Fig. 3a) color that almost disappears in posterior moltings (Fig. 3b). Also, external retrices (juvenal) were narrower shaped, and flight feathers presented aligned growth bars. The preformative molt extent is similar to the one described for Z. capensis in other regions (Pyle et al. 2015) and for other Zonotrichia species (Pyle 1997). Furthermore, 77 % of captured FCF individuals had ossification between 20 and 60 %, and 54 % of the total FCF birds did not get over 40 % ossification. Finally, 77 % FCF individuals presented a chestnut colored iris, and 23 % presented an intermediate iris color, between chestnut and definitive Pratt's Rufous color (Table 1; Fig. 2b).

Figure 3 Color variation in primary coverts of Z. capensis in Bogotá. a. A FPF individual with buffy color on the external border (arrow) of the retained primary coverts and, b. a DCB individual without the buffy border after molting the same group of feathers.
After FCF, Z. capensis turned into a second-cycle prebasic molt (SPB) individual. At this stage, 64 % of the individuals still have skull windows, most of them with skull ossification between 80 and 95 %. Also, 68 % of the birds presented an intermediate iris color between chestnut and Pratt's Rufous (Fig. 2c). Finally, juvenal primaries, secondaries, retrices and primary coverts can be identified before they are completely molted, according to the descriptions of the FCF birds. After second prebasic molt, Z. capensis enters into the second-cycle basic (SCB) age class. Birds in this stage did not show differences with subsequent plumages. Nevertheless, some individuals, presumably at the beginning of the SCB, presented skull windows and iris that did not reach a Pratt's Rufous coloration yet (Fig. 2c) permitting age identification. However, at this stage, Z. capensis develops the final iris color, Pratt's Rufous, and completes skull ossification, making birds indistinguishable from birds in definitive-cycle basic (DCB). This forces categorization as after first cycle (FAJ), following Wolfe et al. (2010) and Johnson et al. (2011).
Finally, definitive prebasic molt (DPB) birds differ from previous molt stages because the primaries, secondaries and primaries coverts are not juvenal and do not present the characteristics described in the FCF section. Furthermore, the individuals have a Pratt's Rufous iris (Fig. 2d) and a completely ossified skull. The definitive-cycle basic (DCB) birds have a plumage that will not change in posterior cycles. Moreover, they present a Pratt's Rufous iris color and 100 % skull ossification.
Additionally, I captured several times an individual 7.4 years old, first banded by the Ornithology Group of Universidad Nacional de Colombia (GOUN) on the 20th Sep 2006 and last time captured on the 16th Feb 2014. This bird allowed me to confirm the characteristics of DCB. Also, outstanding captures were two FCF females with high feather wear, category three over four, and developed brood patch. Moreover, seven males with suspended molt (P2-3), all with a cloacal protuberance. These captures highlight that in Bogotá Z. capensis reach sexual maturity and can reproduce at an early age class, during the FCF. Finally, the seven males are evidence of molt suspension due to overlapping with reproduction. Similar cases were found in Tucumán, Argentina (King 1972, 2009).
DISCUSSION
In Bogotá Z. capensis has a Complex Basic Strategy. All prebasic molts, including pre juvenal molt, are complete molts. Only the preformative molt is a partial molt. The strategy and extent of the molts in Bogotá do not differ from the populations around Cali, Colombia (~3° North; Miller 1961). Furthermore, both populations did not show evidence of alternate plumage, differently from several studies occurred at higher latitudes such as Costa Rica (~10° North, Wolf 1969), Peru (~12° South, Davis 1971) and Chile (~30° South, Pyle et al. 2015), where an alternate plumage was present. However, studied populations of Z. capensis in Tucuman, Argentina (King 1972, 2009) did not show prealternate plumage either.
A second important fact about molt of Z. capensis is that reported by Miller (1961) indicating individuals going through two complete prebasic molts in a single calendar year. Is it possible that the incomplete alternate molt reported in Peru by Davis (1971), which included 2-6 primaries, became a complete molt in tropical latitudes? If it is a complete prealternate molt, there is no way to differentiate it from a complete prebasic molt. Even if two molting peaks occur in Cali and Bogotá during the same year (Miller 1961, Zerda et al.1986), neither of them may be associated with prebasic or prealternate molt, due to the existence of complete molts all year around (Miller 1961, Zerda et al. 1986). Differently to temperate areas, where most birds respond to an annual climate cycle (Pyle 1997), in the Neotropics Z. capensis is subjected to a two-cycle climate year (Miller 1961, Zerda et al. 1986). Here, energy consuming activities such as reproduction have two seasons per year, and a single individual can be part of both (Miller 1961, 1962). I think, there are reasons to believe that Z. capensis has two complete prebasic molts, instead of one prebasic and one prealternate molts. Nevertheless, the discussion remains open.
Finally, I want to underline that females are sexually mature at the FCF age. In temperate zones, for most birds, there is only one favorable season for reproduction, when several resources become available. Thus, hatching individuals wait for a complete year to the next reproduction season. Contrastingly, the Neotropics have two climate cycles per year and some species, as Z. capensis, have two reproductive seasons (Miller 1962). This way, hatched birds concur with a reproductive peak that occurs approximately six months later, and if they reach sexual maturity, they will start to reproduce. However, subsequent molts will be delayed due to the high energy demand during reproduction and this will lead to a trade-off between good quality plumage and early reproduction.
There is not clarity regarding which environmental factors in the Neotropics trigger the physiological process that leads to seasonal reproduction. However, Wikelski et al. (2000) presented evidence that tropical birds can respond to photoperiod variation and used it to synchronize reproduction.

