











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Colombia es uno de los países con mayor diversidad biológica a nivel mundial (Colciencias 1999, UNEP-WCMC c2014). Los Andes colombianos albergan gran parte de esta diversidad, son importantes centros de especiación y alojan un alto número de especies endémicas (Myers et al. 2000, Kattán 2003), lo que convierte estos ecosistemas en hábitats prioritarios para la conservación biológica (Myers et al. 2000). Además, los bosques son fuente de servicios ecosistémicos fundamentales en el desarrollo de la vida humana (Kattán 2003, Tscharntke et al. 2005).
La importancia ecológica, social y cultural de los ecosistemas Andinos, viene acompañada de una fuerte presión antrópica que ha generado pérdida de hábitat, principalmente en los bosques premontanos entre los 1000 y 2400 m. En dichos paisajes predomina la agricultura y la ganadería, esta última con la mayor huella ecológica (Matson et al. 1997). Producto de esta fragmentación y del aislamiento a que conduce, se han confinado las especies silvestres en pequeños parches de bosque, con pérdidas irreversibles de especies nativas (Perfecto et al. 2009) amenazando la biodiversidad a largo plazo. La intensificación de la agricultura, responsable en parte de la crisis ecológica en paisajes rurales (IAASTD 2009), tuvo un impulso a partir de los años 40 y un pleno desarrollo entre los 60 - 70 (Perfecto et al. 2009). El término “intensificación” fue acuñado desde 1965, su definición ha sido muy amplia y se ha visto más relacionada con la productividad y las prácticas de manejo en las fincas. Desde otro enfoque, un poco más reciente y en el cual se fundamentó este estudio, se tuvieron en cuenta otras variables relacionadas con la medición de componentes estructurales de la vegetación, los cuales son fundamentales en la recuperación de la conectividad ecológica y servicios que podrían ofrecen estos paisajes (Ruiz-Martínez et al. 2015).
El café con sombra es uno de los agroecosistemas más emblemáticos debido a que tiene un gran potencial en la conservación biológica de árboles nativos (Perfecto et al. 2009), además, es de gran importancia para enfrentar el cambio climático (Jha et al. 2011). En Colombia se ha documentado que la pérdida de los árboles de sombrío conlleva a la pérdida de la biodiversidad de artrópodos como las hormigas (Armbrecht et al. 2005, Gallego-Ropero 2005, Urrutia-Escobar y Armbrecht 2013) y abejas, mostrando que este último grupo se favorece con la riqueza florística de los cafetales (Cepeda-Valencia et al. 2014). La diversidad de la flora asociada a los cafetales ha sido poco estudiada en Colombia, aunque cierta atención se le ha dado a las arvenses (Gómez-Aristizábal y Rivera-Posada 1987) y a los árboles (Sánchez-Clavijo et al. 2007).
En vista del grave estado en el que se encuentran los hábitats naturales, es necesario desarrollar prácticas de aprovechamiento que involucren el uso sostenible de los recursos naturales. La agroecología provee la posibilidad de un manejo sustentable con producción y conservación simultánea debido a que los procesos ecológicos en los agroecosistemas son entendidos y direccionados para mantener la multifuncionalidad (Armbrecht 2016). En este sentido, el presente estudio tuvo como objeto examinar la importancia de los cafetales como fuente de hábitat para la diversidad vegetal y aplicar un índice de intensificación agrícola (IA) de café para describir cómo responden las variables estructurales de la vegetación en cafetales de sombra, cafetales a libre exposición y parches de bosques (control positivo). Por otra parte, se pretende discutir el valor de los cafetales como refugio de especies, útiles en procesos de recuperación y reforestación en el suroccidente colombiano, particularmente, en la meseta de Popayán, Cauca.
MATERIALES Y MÉTODOS
Área de estudio
Los predios estudiados se encuentran distribuidos entre los municipios de Popayán y Cajibío, en la región de La Meseta Caucana que se extiende entre la cordillera occidental y central de los Andes Colombianos. Este altiplano presenta gran diversidad de hábitats por su variado relieve (Ramírez-Chaves et al. 2008). En esta área se establecieron 20 localidades experimentales conformadas por ocho cafetales a plena exposición o cafetales de sol, ocho cafetales con sombrío de árboles y cuatro bosques de referencia localizados entre los 1600-1750 m (Tabla 1). El estudio completo se realizó entre 2014-2017, concentrando los muestreos entre 2014-2016. Esta zona de vida pertenece al Bosque húmedo premontano (bh-pm), su topografía es ondulada, presentando abundantes vaguadas que escurren al río Palacé. La temperatura entre 12,3ºC y 25,7ºC, cuentan con régimen pluviométrico bimodal, nivel anual de lluvias medias de 2119 mm y promedio de días lluviosos al año entre 170 y 220, redundando en un régimen elevado de humedad relativa con valores que alcanzan hasta el 72 % (Alcázar et al. 2002).
Cálculo del índice de intensificación agrícola (IA)
En la medición del IA se seleccionó el valor óptimo (mayor porcentaje de dosel, mayor densidad de cafetos, mayor riqueza de especies y así en cada caso) de cada variable y a partir de éste, se halló el aporte proporcional (en una escala de cero a uno) por variable en cada sitio experimental. El valor óptimo se consideró aquel del bosque más preservado, en otras palabras, el valor que tiene el ecosistema natural menos intervenido por el ser humano. En algunas variables, como la densidad de cafetos (la cual denota mayor intervención), el solo cociente fue directamente proporcional al valor de la intensificación. Por el contrario, para las variables como la cobertura de dosel o profundidad de la hojarasca, que a medida que aumentan reflejan una menor intensificación agrícola, es decir indican mejor estado del hábitat, se restó a uno al valor del cociente (Mas y Dietsch 2003).
El IA por cada variable oscila entre cero y uno, siendo uno el más intensificado y cero el más conservado. Por otro lado, el valor total del IA dependerá del total de variables estudiadas, como en este estudio se evaluaron trece variables estructurales de la vegetación, el valor máximo de IA será trece.
Medición del índice de Intensificación Agrícola (IA)
Los IA en sus orígenes fueron enfocados en la medición de variables relacionadas únicamente con el manejo agrícola del concepto de agricultura de revolución verde, por ejemplo variables como cantidad de nitrógeno/ha, o dinero invertido en la producción, o aplicación de agroquímicos (Ruiz-Martínez et al. 2015). Sin embargo, desde hace unos años se ha venido replanteando este índice en trabajos enfocados en medir la similitud de estos paisajes con hábitats naturales y como fuente de hábitat, ya que estos parámetros son de mayor relevancia en la evaluación de la conservación de la biodiversidad, la sostenibilidad de la producción, así como la recuperación de los servicios ecosistémicos (Ruiz-Martínez et al. 2015). El IA se determinó a partir del protocolo de Mas y Dietsch (2003) y seguido por Armbrecht (2003). Así, en cada finca donde se realizó la experimentación se establecieron dos parcelas circulares de 12 m de radio (P2) dejando 50 m entre ellas. En el centro de la P2 se delimitó una sub-parcela de 5m de radio (P1), así; la P2 incluyó la P1. Por otro lado, en la P2 se trazó un plano cartesiano con trece estaciones (En). En el centro se dispuso la estación cero (E0) y a partir de ésta, seis estaciones en cada eje separadas por cuatro metros entre sí. En dichas parcelas se realizó la medición y conteo de la vegetación, así como las variables de hábitat.
En este caso, las variables estuvieron enfocadas en la medición de la estructura de los agroecosistemas y en la valoración de la calidad del hábitat, las unidades de cada una dependerán de la naturaleza de la medición como: (1) profundidad de la hojarasca (PH), (2) cobertura de dosel (CD), (3) riqueza de árboles (RA), (4) diámetro de arbustos (DAPa), (5) altura de arbustos (Aa), (6) densidad de arbustos (Da), (7) diámetro de árboles (DAPA), (8) altura de dosel (AD), (9) densidad de árboles (DA), (10) densidad de cafetos (DC), (11) altura de cafetos (AC), (12) riqueza de epífitas (RE) y (13) densidad de epífitas (DE). Teniendo en cuenta que la biodiversidad en agroecosistemas se puede descomponer en dos componentes, biodiversidad planeada y la asociada (Swift et al. 1996), las variables aquí medidas dependen de un plan humano, de manera que la decisión y manejo que el agricultor dé a su agroecosistema afecta las condiciones del hábitat que se genera (Swift et al. 1996). Las arvenses no se incluyeron en este estudio dado su variabilidad por el recambio anual y la frecuencia y métodos variables con que son controladas por los diferentes caficultores.
Medición de la vegetación
En la parcela central uno (P1) se contaron los cafetos y se midió su altura. En la parcela dos (P2), que incluyó la P1, se hizo un conteo de arbustos (2,50 ≤ DAP ≤ 8,10) y árboles (DAP > 8,11 cm) a los cuales se les midió tanto el diámetro a la altura de pecho (DAP) como la altura. La altura se calculó con la ayuda de los tubos de una pértiga. La medición de las epífitas se hizo en todos los árboles con DAP > 8,11cm y hasta una altura máxima de 10 m.
En el conteo de epífitas se incluyeron todas aquellas con una altura superior a 20 cm. En dicho proceso se realizó un pre muestreo, donde las plantas se clasificaron hasta especie y morfo-especie (en el caso de las especies desconocidas). Luego de generar un listado base, se hizo un conteo con la ayuda de binoculares. En el caso de especies desconocidas o con inquietudes taxonómicas, se recolectó material botánico.
Medición de variables de hábitat
En cada zona experimental, así como en las inmediaciones, se hizo una descripción del hábitat y de los usos del suelo más relevantes.
En cada una de las 13 estaciones establecidas en la P2 se midieron la profundidad de la hojarasca (PH) y la cobertura de dosel (CD). Para calcular la PH se abrió un espacio entre la hojarasca hasta tocar suelo firme, seguido, con un pie de rey se midió el espesor de la hojarasca con la aguja de medir profundidades. En la medición de la CD se empleó un Densitómetro Forestal GRS. En cada estación (En) se realizaron cinco mediciones, una en el centro y a partir de ahí, cuatro mediciones formando una cruz a un metro de distancia. Con estos cinco datos, presencia y ausencia de dosel, se halló el porcentaje de dosel (CD).
Recolección e identificación de muestras vegetales
En cada localidad se recolectaron dos muestras por cada planta, tanto de flora arbórea como epífita. También se realizó recolección libre de especímenes con flores y frutos con la finalidad de completar el listado de especies, sobre todo de aquellas que se encontraban adyacentes a los sitios de muestreo.
Las muestras fueron prensadas, etiquetadas y permeadas con etanol al 70 %. Posteriormente el material se procesó en el herbario Luis Sigifredo Espinal, CUVC, de la Universidad del Valle, Cali, donde fue identificado por especialistas, especialmente en orquídeas. Por otro lado, se empleó la base de datos de Trópico-Home del Missouri Botanical Garden, Herbario-Colecciones Científicas en Línea de la Universidad Nacional de Colombia (COL) y Neotropical Herbarium Specimens - The Field Museum, para la identificación y actualización taxonómica.
Análisis de datos
Se registraron los aportes porcentuales por variable en cada localidad (sitio experimental) y la sumatoria de los mismos fue la que finalmente correspondió al valor del índice IA. Ya que se midieron trece variables, el valor máximo de intensificación fue trece. A partir de estos datos se hizo el Análisis de Componente Principales (ACP) en el software R versión 3.2.2. (R Development Core Team c2010), en el cual se usa una transformación ortogonal con el fin de convertir el conjunto de observaciones de variables correlacionadas en un conjunto de valores de variables linealmente no correlacionadas, es decir los componentes principales, con base en las variables estructurales involucradas en la medición de la IA. De este modo, se puede visualizar de una manera más sencilla la estructura subyacente que determina las tendencias en la intensificación agrícola del cultivo del café desde que el bosque es transformado.
RESULTADOS
En términos generales, los bosques presentaron los menores IA, donde el valor mínimo alcanzó 2,2 y el máximo 4,2. Los plantíos de café con sombra presentaron los menores valores de intensificación agrícola, con extremos entre 6,2 y 9,8; mientras que los cafetales a plena exposición o de sol obtuvieron los valores de IA superiores, con un mínimo de 10,1 y un máximo de 12,6 (Fig. 1, Tabla 2).
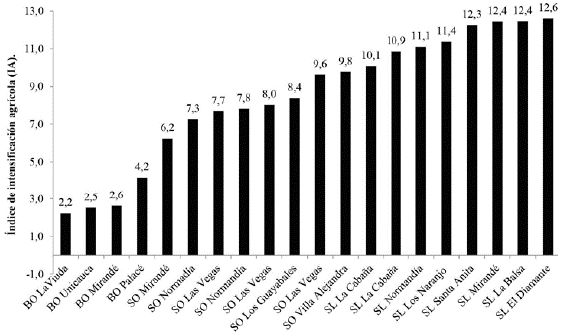
Figura 1 Índice intensificación agrícola en los 20 sitios de estudio. Se incluyen bosques (BO), cafetales de sol (SL) y de sombra (SO) ubicados en La Meseta Caucana.
Tabla2. Índice de intensificación agrícola (IA) en 20 localidades de la Meseta caucana de cada una de las 13 variables de estructura vegetal analizadas.
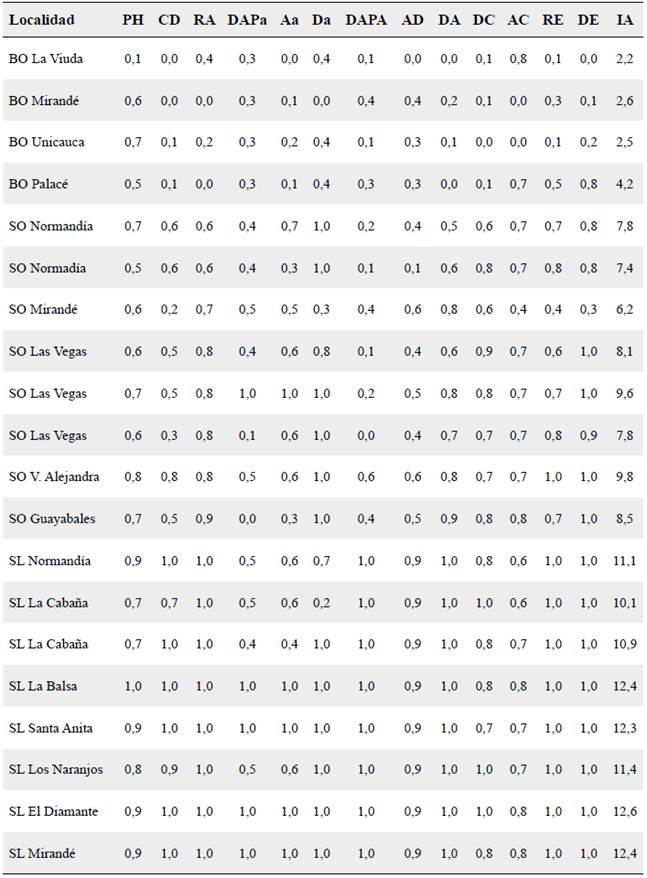
PH = Profundidad de la hojarasca, CD = porcentaje de la cobertura de dosel, RA = riqueza de árboles, DAPa = diámetro de arbustos, Aa = altura de arbustos, Da = densidad de arbustos, DAPA = diámetro de árboles, AD = altura de dosel, DA = densidad de árboles, DC = densidad de cafetos, AC = altura de cafetos, RE = riqueza de epífitas y DE = densidad de epífitas.
El análisis de componentes principales (ACP) (Fig. 2) muestra que la mayoría de las variables estructurales estudiadas estuvieron representadas en el factor uno el cual reunió el 68,16% de la variabilidad, con excepción de la variable altura de cafetos (AC) que estuvo representada en el factor dos, el cual alcanzó 11,53 % de la variabilidad. Por otro lado, la relación de la densidad arbustiva (Da) con los dos componentes no fue evidente, presentando sesgo hacia el componente dos.
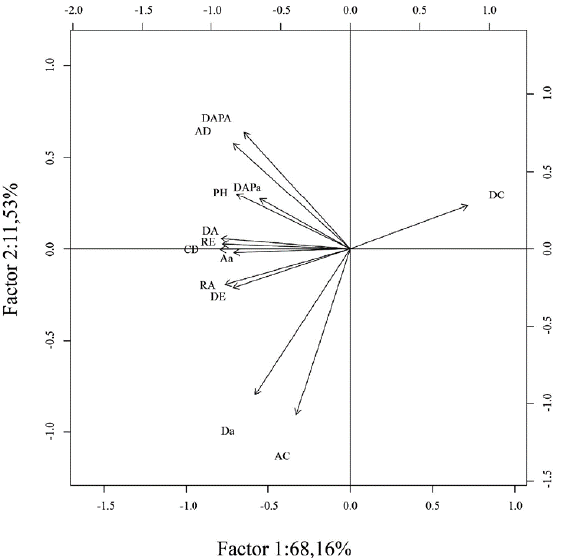
Figura 2 Correlación del Análisis de Componentes Principales (ACP) de las variables de intensificación y los aportes de correlación en cada factor. La desviación estándar del factor 1 es de 2,97 y del factor 2 es de 1,22. Para convenciones ver Tabla 1.
La proyección de las localidades distribuidas en los dos componentes evidenció un gradiente de manejo en función del componente uno (Fig. 3). Los bosques estuvieron agrupados hacia la izquierda del componente uno, al igual que las variables que denotaban calidad del hábitat. A diferencia de esto, los cafetales de sol estuvieron opuestos a los bosques, justo donde se localizó la variable que midió la densidad de cafetos (DC), una variable que da cuenta de la intensificación agrícola en las plantaciones de café, ya que el estrato es principalmente arbustivo. Entre tanto, los cafetales con sombra estuvieron en la mitad de estos dos paisajes, evidenciándose así un gradiente de intensificación agrícola en función de la variación estructural del estrato arbóreo, arbustivo y epífito (Fig. 3).
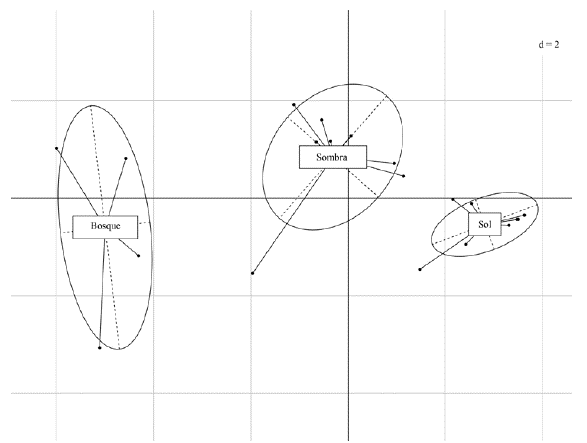
Figura 3 Representación gráfica de los tipos de agroecosistemas sobre los dos primeros componentes principales obtenidos en el ACP que muestra las variables locales evaluadas y que explican el 79,69 de la varianza del sistema (ver Figura 2).
DISCUSIÓN
Las variables estudiadas en esta investigación que mejor describen la intervención antropogénica, medida como la IA, se relacionan con la pérdida estructural de las comunidades vegetales en los agroecosistemas de café a medida que se intensifica su manejo eliminando la vegetación arbustiva y arbórea. Así, el índice de intensificación agrícola fue sensible a la riqueza arbórea (RA), densidad arbórea (DA) y cobertura de dosel (CD), así como la riqueza y densidad de epífitas (RE y DE respectivamente) y la altura de los arbustos (Aa), todas estas relacionadas con la calidad del hábitat. Así pues, la estructura y la flora de los ecosistemas naturales y agroecosistemas, no solo es hábitat para otros organismos, sino que dicha flora sustenta y provee recursos a la fauna asociada (Philpott y Bichier 2012, Valencia et al. 2014, Armbrecht et al. 2005). El índice de intensificación aplicado a diferentes estrategias de manejo en cafetales puede constituirse en una herramienta útil para examinar científicamente otros efectos ecológicos o agronómicos de interés. Por ejemplo, se podría poner a prueba si, relacionado con el índice, ocurre pérdida o ganancia de productividad a largo plazo, erosión, regulación hídrica, cambios en la diversidad de otros grupos biológicos como vertebrados o invertebrados, entre otros, que podrían ser utilizados como variables de respuestas, considerando el IA como el tratamiento o variable independiente ya que constituye una medida objetiva del hábitat.
Por otro lado, la configuración del agroecosistema de café determina las características de dicho hábitat, así; la diversidad y composición de la flora que el caficultor incorpora en sus plantíos se encuentra directamente relacionada con la estructura y con la calidad del hábitat en un lugar (Valencia et al. 2014). Por lo anterior, la medición de la IA en función de las variables estructurales estudiadas, se convierte en una poderosa herramienta para demostrar objetivamente cómo el manejo de los agroecosistemas puede afectar su valor como hábitat para la conservación biológica (Ruiz-Martínez et al. 2015). La densidad de cafetos (DC) afectó de gran manera el IA, resultado consistente con Armbrecht (2003) en Risaralda, situada en el centro-occidente de Colombia. Esta variable presenta mayor relación con directa la intervención humana al momento de diseñar el agroecosistema y que afecta claramente la estructura de los agroecosistemas de café. Dicha variable se comportó contrariamente a variables como la riqueza y estructura arbórea y arbustiva y se relaciona directamente con la tecnificación de la caficultura que favorece especialmente una dimensión de los agroecosistemas, la producción.
Las variables arbóreas fueron fundamentales en la valoración del índice IA, de manera que las fincas que presentaron mayor tecnificación fueron las mismas que tuvieron menor riqueza de árboles, arbustos y epífitas, mayor densidad de cafetos y menor altura de éstos. Características típicas de cafetales de sol o con sombríos mono-genéricos. Así mismo, tales plantaciones fueron muy similares en el valor total del índice de IA, ya que su estructura fue homogénea.
En la finca Normandía, con dos cafetales con sombra diversificada producto de su composición arbórea, estos posiblemente se establecieron mediante la entresaca de madera, tala del sotobosque, incorporación de algunas maderables y siembra de café. Así mismo, presentaron un IA menor y se destacaron por contener especies como Cecropia telealba Cuatrec. (yarumo), Inga edulis Mart. (guamo rabo de mico), Quercus humboldtii Bonpl. (roble), Myrcia popayanensis Hieron. (arrayán), Nectandra lineatifolia (Ruiz & Pav.) Mez. (jigua), Cinnamomum triplinerve (Ruiz & Pav.) Kosterm. (aguacatillo), Alchornea latifolia Sw. (escobo), Meriania speciosa (Bonpl.) Naudin. (flor de mayo), Erythroxylum citrifolum A. St.-Hill.(coca de monte), Viburnum cornifolium Killip & A.C. Sm. y Pinus patula Schiede ex Schltdl.& Cham. (pino pátula) (introducida). Las especies registradas en estas dos localidades estuvieron en la mayoría de los bosques y aunque la riqueza arbórea fue similar, la densidad y abundancia de las mismas fue diferente. En otros cafetales de sombra con menor IA, después de los bosques, como Mirandé, se observó Citrus x limon (L.) Osbeck. (limón mandarino), Citrus reticulata Blanco (mandarina), I. edulis (guamo rabo de mico), Inga sp. (guamo) y Quararibea cordata (Bonpl) Vischer. (sapote). También especies forestales como Jacaranda caucana Pittier. (gualanday) y Samanea saman (Jacq.) Merr. (samán), esta última típica de bosque seco. En contraposición, se destaca la presencia de Ricinus communis L. (higuerilla), una especie muy abundante en el cafetal con sombra de Mirandé, la cual contribuyó a incrementar la estructura arbórea en esta localidad, sin embargo, esta especie presenta alto riesgo de invasión dada su alta diseminación geográfica y su plasticidad en la colonización de hábitats perturbados (Cárdenas et al. 2010).
Otra fuente de diversificación de los sombríos fueron las especies frutales y de importancia agrícola (policultivos), las cuales tuvieron efectos positivos sobre el incremento de la heterogeneidad y riqueza arbórea. Esto acompañado de la vegetación nativa afecta positivamente la estructura del agroecosistema que conlleva a una mejora de las funciones del hábitat, convirtiendo a estos en excelentes amortiguadores del efecto de borde producto de la fragmentación de los bosques (Perfecto et al. 2007, Cepeda-Valencia et al. 2014). Además, pueden funcionar como corredores en paisajes transformados (Pimentel et al.1992, Philpott y Bichier 2012).
De este estudio se destaca que la presencia de árboles de sombra no es un indicativo a priori del valor de conservación en un determinado paisaje agrícola. La mayor parte de los cafetales con sombra de árboles mono-genéricos estuvieron lejos de representar un bosque natural, pues la riqueza arbórea fue muy baja y predominaron especies introducidas como cítricos y guamos (e.g. Inga edulis, o guamo rabo de mico, Inga cf. acreana Harms o guamo machete, I. punctata o guama entre otras). Aún está por estudiarse con mayor detenimiento cuáles son los servicios ecológicos que provee este tipo de sombrío y por qué en el conocimiento tradicional popular se les considera tan valiosos acompañantes en los cafetales en Colombia (Armbrecht, obs. pers. 2002-2018).
Entre tanto, mediante el Análisis de Componentes Principales se visualizó la importancia de la presencia de árboles, la cobertura del dosel, su altura, pero además, la riqueza y densidad de árboles y epífitas como componentes determinantes en la configuración del hábitat en agroecosistemas cafeteros. De hecho, sobre estas últimas se ha demostrado que en cafetales poli-género de manejo orgánico, la identidad y riqueza de epífitas es similar a la del bosque mientras que disminuye en cafetales con mayor intensificación agrícola (Moorhead et al. 2010). El estrato arbóreo constituye la mayor parte de la estructura del hábitat y también provee de hojarasca al suelo, amortigua los cambios extremos en la temperatura, además de ser fuente de hábitat para otros organismos como aves locales (Mas y Dietsch 2004, López-Gómez et al. 2008) y migratorias (Bakermans et al. 2009), epífitas (Goodall et al. 2014), líquenes (Meneses y Escandón, datos no publicados), insectos (Perfecto y Armbrecht 2002, Cepeda-Valencia et al. 2014), entre otros.
Flora en la zona de estudio
Las especies reportadas en la zona de estudio, con un listado de más de 240 especies vegetales (listado disponible con los autores) corresponden al gremio de las heliófilas (HE) o pioneras, las cuales soportan alta luminosidad y colonizan hábitats transformados (Sánchez et al. 2007), hallazgo consistente con lo reportado para México por Valencia y colegas (2014). En este grupo se incluyen especies con un alto valor en la recuperación o enriquecimiento temprano de espacios degradados como Montanoa quadrangularis Sch. Bip. (arboloco) (Tamayo-Rincón et al. 2010) y Baccharis pedunculata (Mill.) Cabrera (chilca), taxones propios de zonas con alto grado de disturbio y suelos ácidos erosionados, esta última estuvo presente en la mayoría de las zonas de estudio. Otras HE encontradas fueron Ochroma pyramidale (Cav. ex Lam.) Urb. (balso negro) (Louman et al. 2001) y Heliocarpus cf. americanus L. (balso blanco) al interior de los bosques y en claros contiguos a los cafetales y Trema micrantha (L) Blume (cagalero), una especie típica de zonas disturbadas con defensas químicas contra herbivoría (Muñiz 2008) y Cecropia telealba, relacionada con hormigas que la protegen de la herbivoría (Holldobler y Wilson 1990). Vismia baccifera (L.) Planch y Triana (aguacatillo), especie de borde, importante como fuente de frutos para las aves, es considerada como una especie que se adapta fácilmente y de rápido crecimiento, ideal para enriquecimiento en etapas tempranas de la sucesión y en el establecimiento de cercas vivas (Higuita et al. 2014). Se resalta la presencia de especies arbustivas como Piper hispidum Sw. (cordoncillo), Piper cf. aduncum L (cordoncillo), Solanum umbellatum Mill. (pepo), Solanum cf. deflexiflorum Bitter., Siparuna aspera (Ruiz y Pav.) A. DC., Clidemia ciliata Pav. ex D. Don (mortiño), Bocconia frutescens L. (trompeto), especies frecuentes en zonas abiertas con alta incidencia de luz, borde de bosque, en vaguadas y bosques de galería. Se encontraron otras pioneras importantes para regeneración temprana como Myrsine guianensis (Aubl.) Kuntze (Primulaceae) y Myrsine latifolia (Ruiz y Pav.) Spreng. (ambas conocidas como cucharo) (Esquivel y Calle 2002) y Myrsine coriaceae (Sw.) R.Br. ex Roem y Schult (espadero) que no ha sido reportada en otros estudios, es evidente el potencial de esta especie en la colonización de espacios con algún grado de disturbio.
Otro grupo perteneciente a este gremio son las heliófilas de larga duración (HD), las cuales se caracterizan por requerir sombra al inicio de su ciclo de vida y alta luminosidad en las últimas etapas de desarrollo, su estructura basal superiores a la de las HE, son especies de dosel o emergente y pueden regenerarse en claros más pequeños al interior del bosque. La mayoría de las especies maderables de uso comercial pertenecen a este grupo (Finegan 1996). Entre estas, se resaltan árboles como Saurauia cf. brachybotrys Turcz (dulumoco), asociada a los bosques de galería y riachuelos, importante fuente de frutos para la avifauna. Oreopanax sp. Decne y Planch (manodioso) con una alta presencia en los bordes de los bosques y rara vez al interior. Hedyosmum cf. bonplandianum Kunt (silbador), una especie de interior y rara vez en borde de bosque, importante en bosques maduros, se caracteriza por colonizar claros al interior de bosque, con alturas superiores a 15 m, al igual que Alchornea latifolia Sw., que se encontró en la mayoría de las localidades y al interior de bosque. Otras especies muy importantes fueron Toxicodendron striatum (Ruiz & Pav.) Kuntze (manzanillo), que se establece fácilmente en suelos deficientes de nutrientes, tiende a formar poblaciones homogéneas y pueden afectar a las personas causando alergia. Ladenbergia oblongifolia (Humb ex Mutis) L. Andersson (cascarillo), Banara guianensis Aubl y Lacistema aggregatum (P.J. Bergius) Rusby (café de monte) fueron árboles muy frecuentes en borde de bosque, carreteras, en plantaciones de café con sombra, con abundantes frutos que son consumidos por aves, especialmente L. aggregatum. Este fue el caso de N. lineatifolia, una de las especies con mayor presencia en bosques y cafetales con sombra diversificada. Esquivel y Calle (2002) mencionan que esta especie es importante en la generación de micro hábitats para el establecimiento de otras especies y presenta gran potencial en la restauración de pastizales, además, es una especie con un importante aporte de frutos para aves grandes y mamíferos pequeños. Por otro lado, C. trinerve y Persea cf. caerulea (Ruiz y Pav.) Mez, especies de borde y bosques ribereños, importantes productoras de frutos pequeños y medianos.
Entre tanto, especies como Syzygium jambos (L.) Alston. (pomarroso) y M. popayanensis fueron muy representativas, S. jambos fue dominante en los bosques de Mirandé y Unicauca, mientras que M. popayanensis estuvo presente en la mayoría de los bosques y cafetales con sombrío. Esta última, se caracterizó por ofertar una gran cantidad de frutos para aves. Otras especies, no tan frecuentes, importantes por su aporte en frutos y de origen autóctono, fueron Tetrorchidium cf. euryphyllum Standl y Calliandra trinervia Benth (carbonero rojo) que estuvieron solo en bosque, así como E. citrifolium que además estuvo en un cafetal con sombra.
Finalmente, se registró un rodal de Q. humboltii en el bosque uno de Normandía y un individuo de dicha especie en el cafetal con sombra diversificada uno de la misma localidad. Esta especie pertenece al gremio de las esciófitas, las cuales son intolerantes a la luz en sus primeras etapas de desarrollo y con altos requerimientos en la culminación de su ciclo de vida ya que son especies de dosel, de maderas duras y finas dado su lento desarrollo, consideradas como especies de bosque maduro (Louman et al. 2001). Q. humboldtii es casi exclusiva de Colombia, se distribuye a lo largo de las regiones montañosas, entre los 1000 y 3000m de altitud. Su madera es muy fina, es empleada en la ebanistería, enchapados, su uso data desde el siglo XIX. Ha sido categorizada en el libro rojo de árboles maderables como vulnerable (VU A2cd) pues sus poblaciones se han visto reducidas en más del 42 %. Actualmente existen poblaciones naturales en distintos parques y santuarios naturales del país (Cárdenas y Salinas 2007).
Flora epífita
Se registró un amplio número de especies de orquídeas, una familia emblemática en el mundo por su uso en la ornamentación. La riqueza de este grupo estuvo concentrada en los bosques mejor estructurados y con menor intervención. Se destacan Bifrenaria racemosa (Hook.). Lindl., R. granadensis y Rodriguezia lanceolata Ruiz y Pav., reportadas por UICN en categoría de preocupación menor (LC), Epidendrum peperomia Rchb.f. y otras especies del género Pleurothallis R. Br., Epidendrum L. y Stelis S.w. Entre las bromelias se reportaron Tillandsia fendleri Griseb, Tillandsia elongata Kunth., Tillandsia racemosa Urb. y Aechmea angustifolia Poepp. y Endl. Así mismo, se reportó un alto número de especies pertenecientes a Polypodium y a Peperomia.
Este trabajo es relevante por cuanto encontró evidencia que los agroecosistemas cafeteros pueden mantener flora nativa y de valor para la conservación como la mencionada; además el trabajo señala que no todo cafetal de sombra, por definición, contribuye igual a conservar la biodiversidad vegetal. Por otro lado, este trabajo documenta el uso de diferentes variables estructurales de la vegetación en la medición de la intensificación agrícola (IA). Se evidenció que estas variables se encuentran estrechamente relacionadas con la medición de la intervención antropogénica, en función de la calidad del hábitat. Esto, convierte este análisis en una herramienta importante en la medición de la degradación estructural de los agroecosistemas. El IA podría usarse en futuros estudios para evaluar la respuesta de otras variables de interés con miras a una caficultura sustentable y amigable con la biodiversidad.

