











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Durante la sucesión secundaria de bosques ocurren cambios importantes en la composición, la estructura y la diversidad funcional de la vegetación (Lebrija-Trejos y Bongers 2008, Lohbeck et al. 2013), los cuales, en especial a nivel funcional, pueden estar relacionados con variaciones en las estrategias ecológicas de las especies (Lebrija-Trejos y Bongers 2008), incidiendo así en diferentes procesos ecosistémicos como el ciclaje de nutrientes o la acumulación de biomasa (Casanoves et al. 2011a, Lohbeck et al. 2015). Por ejemplo, se ha encontrado que los valores comunitarios de área foliar específica (AFE), una relación entre el área y el peso seco, están relacionados con las tasas de acumulación de biomasa durante la sucesión (e.g Lohbeck et al. 2015). A su vez, la acumulación de biomasa es un proceso directamente relacionado con la provisión de servicios ecosistémicos como la captura y almacenamiento de carbono (Casanoves et al. 2011a). Por lo tanto, estudiar los cambios en los rasgos funcionales durante la sucesión secundaria puede ser clave para entender tanto los procesos como los servicios ecosistémicos (Lavorel et al. 2011).
El Dióxido de Carbono (CO2) es el principal gas de efecto invernadero y el incremento de sus concentraciones está relacionado con el aumento de la temperatura a nivel global (Knee 2009); por lo que uno de los servicios ecosistémicos más importantes que actualmente ofrecen los bosques tropicales es la captura y almacenamiento del mismo (Wangming et al. 2014). La captura y almacenamiento de CO2 depende, entre otros aspectos, de las características funcionales de las hojas que determinan la capacidad que tienen estas de interceptar la radiación fotosintéticamente activa (RFA) (Warnock et al. 2006). Por ejemplo, en general, mayores valores de AFE se relacionan con una mayor capacidad fotosintética y en consecuencia, con mayores ganancias de carbono (Villar 2004), lo cual ha sido también identificado para el área foliar (AF) (Taiz y Zeiger 2006), a la par de mostrar una relación negativa con el contenido foliar de materia seca (CFMS) (De La Riva et al. 2014). Adicionalmente, la densidad de la madera (DM), entendida como la inversión o almacenamiento de carbono por unidad de volumen de tronco o rama (Chavé et al. 2009), es el resultado directo del carbono destinado a la producción de tejidos (Chavé 2002).
Los cambios en la diversidad funcional, y su impacto sobre los procesos y servicios ecosistémicos pueden ser analizados a partir del análisis de Tipos Funcionales de Plantas (TFPs) (Casanoves et al. 2011a) e índices de diversidad funcional como la media ponderada de la comunidad (MPC) (Petchey y Gaston 2006). Los TFP son grupos de especies que comparten valores similares de rasgos funcionales independientemente de su relación taxonómica, y que presentan por tanto respuestas y efectos similares en el ecosistema (Díaz y Cabido 1997). La abundancia de los TFPs puede cambiar a lo largo de la sucesión vegetal (Lavorel 1999), lo que a la postre puede influir en los procesos y servicios ecosistémicos que esta genere (Grime et al. 1997).
La MPC es una medida que representa el valor promedio de los rasgos de una comunidad en un momento determinado, y refleja, las respuestas de la comunidad al ambiente o su efecto sobre los procesos y servicios ecosistémicos (Lavorel y Garnier 2002). Se ha encontrado que la MPC de diferentes rasgos tales como la altura máxima, el área foliar específica (Carreño et al. 2012) y el área foliar (Anten y Selaya 2011) cambian a lo largo de la sucesión secundaria. En la sucesión de bosques secos tropicales, en particular, se han evidenciado cambios en la MPC de rasgos como la densidad de la madera (Poorter y Markesteijn 2008, Castellanos 2013) y el contenido foliar de materia seca (Lohbeck et al. 2013). En contraste, hasta donde se sabe, no hay estudios publicados sobre cambios en rasgos funcionales durante la sucesión de bs-T interandinos de Colombia, los cuales muestran grandes diferencias con respecto a bs-T de la zona del Caribe y los llanos orientales (Pizano et al. 2014).
En este estudio se analizaron los cambios en los rasgos funcionales relacionados con captura y almacenamiento de carbono en un gradiente sucesional de un bs-T interandino. Específicamente, se buscó identificar TFP asociados al secuestro de carbono y determinar cómo varían su abundancia y área basal a lo largo de la sucesión. Así mismo, se analizaron las posibles variaciones en la MPC de rasgos relacionados con el secuestro de carbono en el gradiente sucesional. Creemos que la MPC de rasgos funcionales asociados con mayor almacenamiento, y menor captura y liberación de carbono, aumentará a medida que avanza la sucesión; y, en consecuencia, la abundancia y dominancia de los TFPs que se asocian a estrategias conservativas aumentarán, mientras que los TFPs asociados con estrategias adquisitivas disminuirán durante la sucesión.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se llevó a cabo en el área de compensación ambiental y restauración ecológica de la central hidroeléctrica El Quimbo (75°41'44,84", 75°41'26,23" Oeste y 2°18'38,50'', 2°18'19,21'' Norte), localizada en el departamento de Huila (Colombia), en los municipios de El Agrado, Gigante y Garzón. La temperatura media anual es de 24 °C, la precipitación promedio anual es 1100 mm y la altitud varía entre 720 y 970 m. El área corresponde a un bs-T de 71,5 hectáreas, en el que dominan los pastizales abiertos y arbolados, arbustos cerrados y algunos relictos de bosque en pendiente fuerte y leve asociados a los drenajes (Avella-M. et al. 2019).
Se determinaron tres estados sucesionales a partir de la caracterización florística y estructural de la vegetación en el área de estudio (Avella-M. et al. 2019): (i) pastos arbolados con dominio de Guazuma ulmifolia Lam., (ii) arbustales cerrados, y (iii) bosques (Tabla 1); en los cuales se establecieron 14, 9 y 21 parcelas, respectivamente, en consideración a los estados sucesionales determinados por Avella-M. et al. (2019).
Tabla 1 Resumen de la caracterización ecológica de los tres estados sucesionales evaluados en un bosque seco tropical interandino
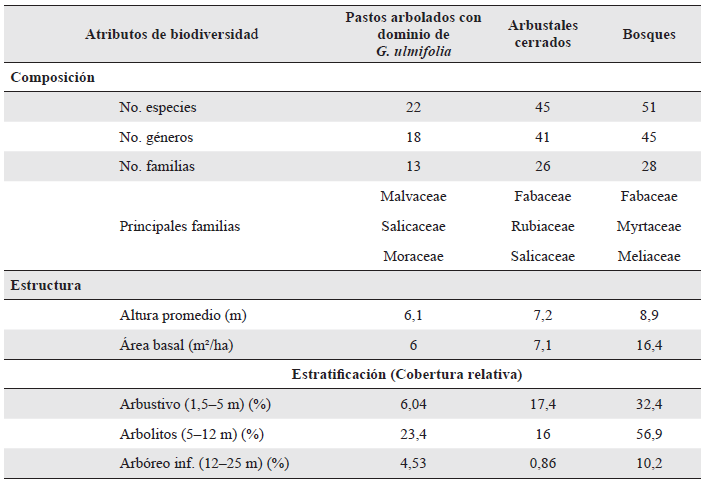
En la zona de estudio la mayoría de los potreros tienen más de 70 años de actividad, en éstos se hacían dos limpias al año con corte manual y en raras ocasiones se utilizaba fuego; sectores de bosques con grandes árboles fueron quemados para ampliar las áreas destinadas a la ganadería, y se mantuvieron algunos relictos para la provisión de recurso hídrico y madera. Producto de esta actividad agropecuaria la zona es un mosaico de pastizales y arbustales, con relictos boscosos asociados a drenajes o a sitios de difícil accesibilidad (Avella-M. et al. 2019)
Selección de especies e individuos
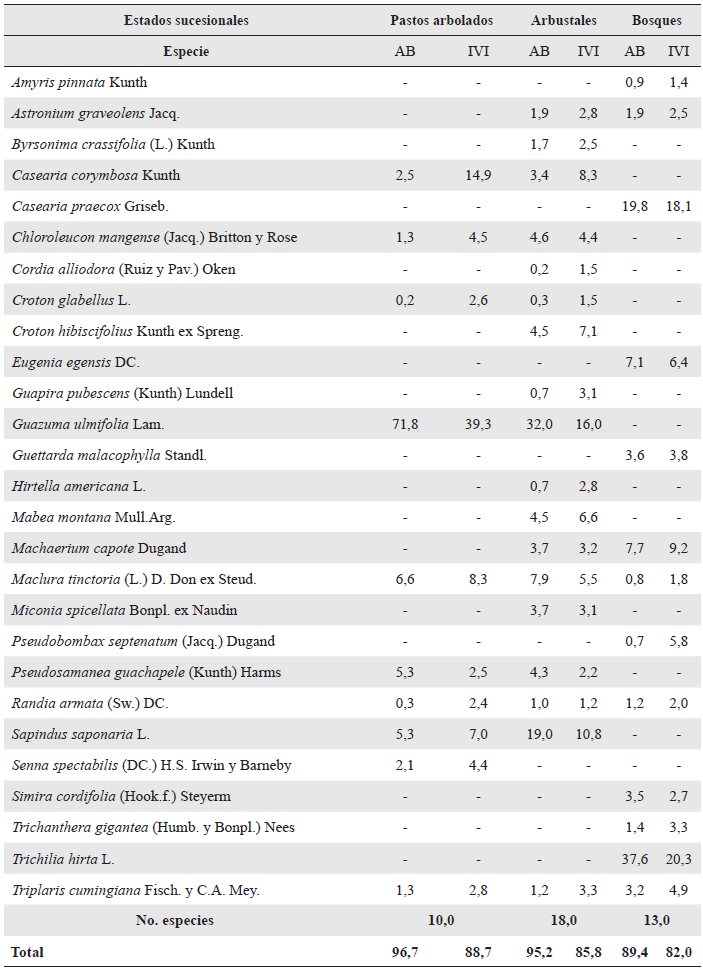
En cada estado sucesional se seleccionaron las especies dominantes que representaron más del 80 % del área basal y del Índice de Valor de Importancia (IVI) (Tabla 2). De acuerdo con la hipótesis de razón de biomasa (Grime et al. 1997), estas especies dominantes tendrían un papel fundamental en el secuestro de carbono (Pérez-Harguindeguy et al. 2013). En el cálculo del área basal se incluyeron individuos con DAP > 2,5 cm. Para la medición de rasgos funcionales se seleccionaron en cada estado sucesional cinco individuos por especie, con excepción de la altura máxima y forma de crecimiento, rasgos para los que se consideraron doce individuos por especie.
Medición de rasgos
Se midieron en total siete rasgos funcionales, cuatro foliares y tres vegetativos, comúnmente analizados en el estudio del secuestro de carbono (Polania et al. 2011). Los rasgos foliares seleccionados fueron: área foliar (AF- cm2), área foliar específica (AFE- cm2 mg-1), contenido foliar de materia seca (CFMS- mg g-1), y fenología foliar. El AF, AFE y el CFMS se midieron, para cada individuo, a partir de diez hojas jóvenes, bien expandidas y con un mínimo rastro de herbivoría o marchitez (Pérez-Harguindeguy et al. 2013). La medición de las áreas foliares se realizó por medio del Software ImageJ (Schneider et al. 2012).
Para la determinación de la fenología foliar se estimó a nivel de individuo la densidad de hojas utilizando la siguiente escala: (1) cuando se trató de individuos sin hojas (0-20 %); (2) para individuos con hojas escasas (20-40 %); (3) para una densidad intermedia de hojas (40-60 %); (4) para una alta densidad de hojas (60-80 %) y (5) para individuos con hojas en todas o casi todas las yemas foliares (80-100 %). Posteriormente, se analizaron las frecuencias con las que cada una de las categorías descritas aparecieron al evaluar los individuos de cada especie, con lo que se categorizó cada especie entre caducifolia, semicaducifolia y perenne, posteriormente se contrastaron dichas categorías con literatura especializada (Geilfus 1989, Figueroa-C. y Galeano 2007, Scholz et al. 2008, Rendón et al. 2013).
Los rasgos vegetativos seleccionados fueron: densidad de la madera (DM- g cm-3), forma de crecimiento y altura máxima (Hmax-m). Para la determinación de la DM se tomaron muestras de ramas de al menos 1 cm de diámetro (Swenson y Enquist 2008). Después de remover la corteza, se determinó el volumen en fresco de cada muestra a partir del método de desplazamiento de agua (Pérez-Harguindeguy et al. 2013). Posteriormente, las muestras se secaron a 103 °C hasta alcanzar un peso constante (24-72 h) y se determinó su peso seco. La densidad de la madera de cada segmento se calculó como la relación entre peso seco y volumen fresco (Pérez-Harguindeguy et al. 2013). La forma de crecimiento se determinó a partir de la revisión de literatura (Geilfus 1989, CATIE 1994, Gómez y Toro 2008, Scholz et al. 2008, Rendón et al. 2013) y según la altura promedio de los individuos en el área de estudio. La altura máxima para cada una de las especies se determinó con base en la información registrada en levantamientos de vegetación en el área de estudio (García datos no publ.) y de acuerdo con lo recomendado por (Pérez-Harguindeguy et al. 2013).
Manejo de datos
Los TFPs se definieron a partir de un análisis jerárquico de conglomerados con los cinco rasgos cuantitativos, aplicando el método de encadenamiento de Ward y la medida de distancia euclídea. Para identificar diferencias significativas entre TFPs se llevó a cabo un análisis de varianza multivariado (MANOVA), utilizando la prueba de vectores medios de Hotelling, posteriormente se caracterizaron los TFPs mediante análisis de varianzas usando modelos lineales generales y mixtos (Di Rienzo et al. 2011 ) para los rasgos cuantitativos. De igual forma, se verificó la relación de los TFPs con los rasgos cualitativos mediante análisis de tablas de contingencia con pruebas de Chi cuadrado y análisis de correspondencias. Finalmente, se determinó si había diferencias significativas en las abundancias y dominancias relativas de TFPs en cada estado sucesional, por medio de un análisis de varianza a partir de modelos lineales mixtos. En cada estado sucesional, se calculó la media ponderada de la comunidad (MPC) (Garnier et al. 2004) de los rasgos cuantitativos a partir de las especies que representaron más del 80 % del área basal e Índice de Valor de Importancia (IVI). Por último, se determinó si había diferencias significativas en los valores de las MPC entre los estados sucesionales evaluados, a partir de modelos lineales mixtos. Los análisis estadísticos se hicieron en los programas estadísticos F-diversity (Casanoves et al. 2011b) e InfoStat (Di Rienzo et al. c2017) mediante las interfaces para R (R development Core Team c2017).
RESULTADOS
Especies seleccionadas para cada estado sucesional
Se seleccionó un total de 41 biotipos en todo el gradiente sucesional (diez en pastos arbolados, 18 en arbustales y trece en bosques), correspondientes a 26 especies, debido a que una misma especie se podía encontrar en diferentes estados sucesionales, así: tres especies se presentaron en los tres estados sucesionales evaluados, ocho en dos estados sucesionales, y 16 en un único estado sucesional (Tabla 2).
Determinación de Tipos Funcionales de Plantas (TFPs)
En el análisis jerárquico de conglomerados se identificaron cuatro tipos funcionales de plantas (Fig. 1). Al evaluar los rasgos distintivos de cada grupo funcional, primero de manera exploratoria a partir del análisis de componentes principales (Fig. 2), y luego de manera inferencial mediante modelos lineales mixtos, se observó que el TFP 2 mostró valores significativamente más bajos respecto a los demás grupos en la densidad de la madera (P « 0,05). De igual forma, el TFP 3 presentó un área foliar específica significativamente menor (P << 0,05) a la que presentaron los restantes tres TFPs.
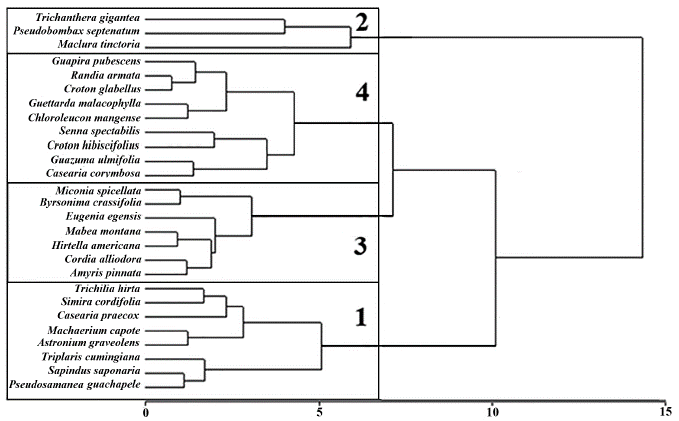
Figura 1 Tipos Funcionales de Plantas (TFPs), dendrograma obtenido mediante análisis jerárquico de conglomerados, usando método de Ward y distancia Euclídea, a partir de las variables (rasgos) área foliar, área foliar específica, contenido foliar de materia seca, densidad de la madera y altura máxima. Los números en las ramas señalan cada tipo funcional de planta.
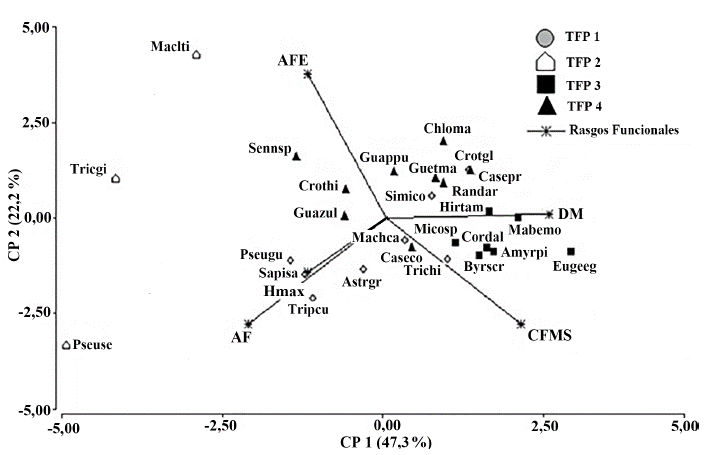
Figura 2 Análisis de Componentes Principales (ACP) para rasgos funcionales cuantitativos y TFPs, AF: Área Foliar. AFE: Área Foliar Específica, CFMS: Contenido Foliar de Materia Seca, DM: Densidad de la madera, Hmax: Altura Máxima. Especies (Ver Tabla 2): Maclti, M. tinctoria; Tricgi, T. gigantea; Pseuse, P. septenatum; Sennsp, S. spectabilis; Crothi, C. hibiscifolius. Guazul, G. ulmifolia; Psegu, P. guachalepe; Sapisa, S. saponaria; Tripcu, T. cumingiana; Astrgr, A. graveolens; Machca, M. capote; Guappu, G. pubescens; Simico, S. cordifolia, Caseco, C. corymbosa. Trichi, T. hirta; Micosp, M. spicellata; Guetma, G. malacophylla; Randar, R. armata; Chloma, C. mangense, Crotgl, C. glabellus; Casepr, C. praecox; Hirtam, H. americana; Mabemo, M. montana; Cordal, C. alliodora, Amyrpi, A. pinnata; Byrscr, B. crassifolia; Eugeeg, E. egensis.
Tanto el área foliar como la altura máxima presentaron valores significativamente más altos en los TFPs 1 y 2 respecto a los TFPs 3 y 4 (P << 0,05 y P << 0,01, respectivamente). Es importante resaltar que los valores de contenido foliar de materia seca fueron significativamente diferentes entre los cuatro grupos (P << 0,01), siendo el TFP 3 el que presentó los más altos. El rasgo que reflejó una mayor variación entre los grupos fue la densidad de la madera, mientras que el área foliar específica mostró una variación menor. Se encontró una asociación entre los TFP, las formas de crecimiento, y la fenología foliar. En este sentido, los TFPs 1 y 2 se asociaron con el hábito arbóreo (P << 0,01) y fenología semicaducifolia (P = 0,05), el TFP 3 al estrato arbustivo y la fenología perenne, el TFP 4 al estrato arbustivo y la fenología caducifolia.
De acuerdo con lo anterior, los cuatro tipos funcionales de plantas se definieron así (Tabla 3): El TFP 1, caracterizado por árboles semicaducifolios de dosel, baja inversión en estructuras fotosintéticas y potenciales bajos valores de crecimiento relativo. El TFP 2 corresponde a árboles semicaducifolios de subdosel con alta inversión en estructuras fotosintéticas y tallos. El TFP 3 fue conformado por arbustos perennes con hojas pequeñas, baja inversión en estructuras fotosintéticas y potenciales bajos valores de crecimiento relativo, el TFP 4 compuesto por arbustos caducifolios, con hojas pequeñas y una alta inversión en estructuras fotosintéticas. En este sentido, los TFPs se agruparon en dos estrategias (Figura 2, Tabla 3): especies adquisitivas, con bajos valores de contenido foliar de materia seca, alta área foliar específica, con tendencia a la caducifolia y densidad de la madera de media a baja (TFPs 2 y 4); y especies conservativas con altos valores de contenido foliar de materia seca, baja área foliar específica y con tendencia a mantener las hojas durante el año (TFP 1 y 3).
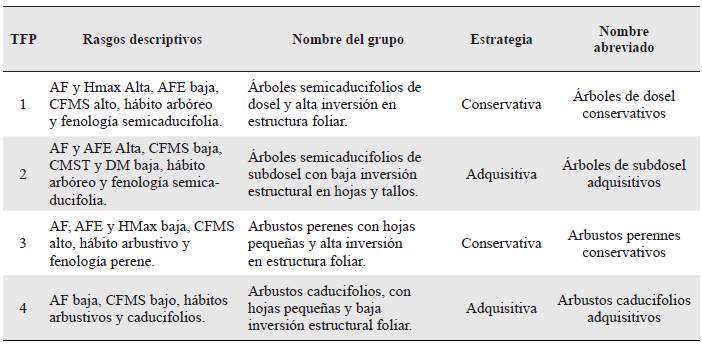
Variación en la dominancia de TFPs en el gradiente sucesional
En cada estado sucesional se determinó la dominancia de los TFPs en términos de abundancia y área basal, en los pastos arbolados con dominio de Guazuma ulmifolia el TFPs con mayor abundancia y área basal fue el 4 (arbustos caducifolios adquisitivos, P = 0,01); mientras que el TFP 3 presentó los valores más bajos (Fig. 3). Los arbustales fueron dominados por los TFPs 3 (arbustos perennes conservativos) y 4 (arbustos caducifolios adquisitivos, P = 0,05), los cuales presentaron una abundancia relativa significativamente mayor en este estado sucesional, que los TFPs 1 (árboles de dosel conservativos) y 2 (árboles de subdosel adquisitivos, Fig. 3). En los bosques, el TFP1 fue más abundante (P << 0,01) que los demás TFP, los cuales presentaron una abundancia similar entre sí (Fig. 3). Sin embargo, en términos de área basal relativa, en los bosques se encontró una dominancia estadísticamente mayor (P = 0,01) de los dos TFPs asociados a estrategias conservativas, es decir, árboles de dosel conservativos (TFP 1) y arbustos perennes conservativos (TFP 3), respecto a los dos TFPs asociados a estrategias adquisitivas (2 y 4).
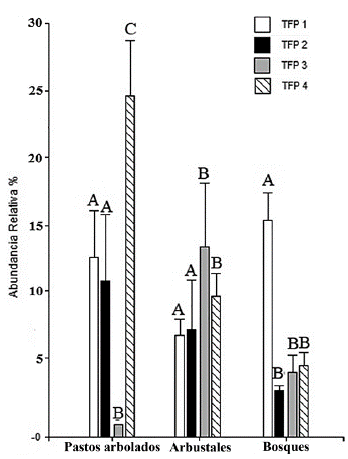
Figura 3 Abundancia relativa de Tipos Funcionales de Plantas (TFPs) en cada estado sucesional. Letras diferentes en cada estado sucesional significan diferencias significativas (Prueba de DGC, P < 0,05; n = 44).
En general, los patrones de variación de la abundancia y el área basal de los TFPs a lo largo de la sucesión fueron similares. La abundancia relativa de los TFPs 2 (árboles de subdosel adquisitivos) y 4 (arbustos caducifolios adquisitivos) disminuyó a lo largo de la sucesión (P << 0,01 y P = 0,01 respectivamente; Fig. 4), dicho patrón se mantuvo también para el área basal relativa del TFP 4 (P = 0,04), pero no para el TFP 2 (P = 0,18). En contraste, los patrones de abundancia y dominancia de los TFP 1 y 3, relacionados con estrategias conservativas, fueron variables. El TFP 1 (árboles de dosel conservativos), aumentó su área basal relativa de manera significativa en los bosques (P << 0,01), pero no mostró dicho aumento en su abundancia relativa de manera significativa (P = 0,07, Fig. 4). Por otro lado, el TFP 3 (arbustos perennes conservativos) mostró un aumento en su área basal relativa (P = 0,01) y abundancia relativa (P = 0,04, Fig. 4) en los arbustales y bosques, respecto a los pastos arbolados, pero redujo su abundancia relativa en el paso de arbustales a bosques.
Índices de Diversidad Funcional
La MPC de los rasgos foliares cambió en los estados sucesionales (Fig. 5). Por un lado, la MPC del área foliar fue mayor en los bosques (P << 0,01). Por el contrario, la media ponderada de la comunidad del contenido foliar de materia seca fue menor en los pastos arbolados (P << 0,01), pero no mostró diferencias significativas en el gradiente sucesional para el área foliar específica (P=0,25). En cuanto a los rasgos vegetativos, tanto los valores de MPC de la densidad de la madera como de la altura máxima fueron mayores en los bosques (P << 0,01 y P = 0,01 respectivamente). A diferencia de los demás rasgos, la MPC de la densidad de madera aumentó significativamente a lo largo de los tres estados sucesionales.
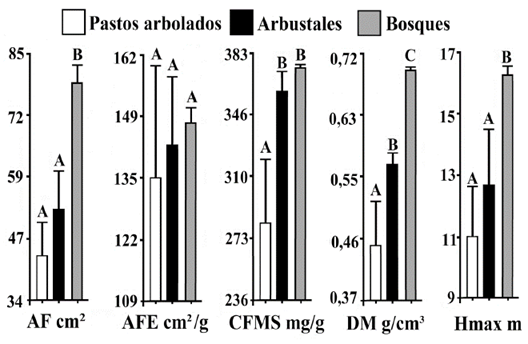
Figura 5 Comparación de Media Ponderada de la Comunidad (MPC) entre estados sucesionales. AF: Área Foliar en cm2. AFE: Área Foliar Específica en cm2/g. CFMS: Contenido de Foliar de Materia Seca en mg/g. DM: Densidad de la madera en g/cm3. Hmax: Altura Máxima en m. Medias con una letra común no son significativamente diferentes (Prueba de DGC, P < 0,05; n=44).
DISCUSIÓN
Tipos Funcionales de Plantas asociadas a secuestro de carbono en bs-T
Los grupos asociados a estrategias adquisitivas (TFPs 2 y 4), se caracterizan por presentar valores de rasgos que favorecen la adquisición rápida de recursos, altas tasas de fotosíntesis, y una rápida retención y liberación de carbono y otros elementos (Grime et al. 1997). En contraste, los dos TFPs restantes (1 y 3) se relacionaron con estrategias conservativas, las cuales se asocian a tasas fotosintéticas y de captura de carbono reducidas (Salgado-Negret et al. 2016), con una mayor retención y almacenamiento del carbono en sus estructuras (Wright et al. 2004).
Esta diferenciación se ha reportado en otros ecosistemas secos del caribe colombiano por Montes et al. (2017), quienes encontraron un TFP asociado a estrategias conservativas y otro a especies adquisitivas, utilizando también como principales variables para su diferenciación el área foliar específica y el contenido foliar de materia seca. Adicional a estos rasgos, la forma de crecimiento fue un parámetro diferenciador, puesto que los TFP 3-4 están asociados a hábitos arbustivos y los TFP 1-2 al hábito arbóreo. Patrón evidenciado también al analizar un gradiente climático en China por Jiang y Dong (2000).
Variación de TFPs asociados al secuestro de carbono en el gradiente sucesional
Los arbustos caducifolios adquisitivos (TFP 4) fueron los más abundantes en los pastos arbolados con dominio de G. ulmifolia. Este patrón podría obedecer a una mayor capacidad de adaptación a condiciones que exigen alta eficiencia hídrica, expresada en rasgos que contribuyen a tolerar o evadir la sequía (Aranda et al. 2014), en este caso la presencia de hojas caducifolias (Salgado-Negret y Paz 2016) o a una menor área foliar, lo que contribuye a disminuir la respiración y la pérdida de agua (Kucharik et al. 1998). Por lo tanto, es posible inferir que el principal filtro ambiental que configura la composición de rasgos en la comunidad es la disponibilidad de agua (Ceccon y Hernández 2009), seguido por el tipo de perturbaciones antropogénicas (Foley et al. 2005) debido a que el área de estudio estuvo sometida a una ganadería tradicional extensiva (Avella et al. datos no publ.) en la que era frecuente dejar algunos árboles de G. ulmifolia para favorecer el sombrío y forrajeo del ganado.
Nuestra hipótesis inicial planteó un recambio de especies a lo largo del gradiente sucesional, en el que aumentan la abundancia y dominancia de TFPs asociados a estrategias conservativas, mientras disminuyen las de adquisitivas, en efecto, en el gradiente sucesional disminuyó la abundancia de los TFPs adquisitivos (2 y 4) que podrían presentar rápido crecimiento producto de una baja inversión estructural en hojas (Salgado-Negret et al. 2016) y altas tasas fotosintéticas (Wright y Westoby 2002), así como madera blanda (TFP 2) y hojas caducifolias (TFP 4); así mismo, se presentó un aumento en la dominancia de los árboles de dosel conservativos (TFP 1) relacionados con bajas tasas fotosintéticas (Pérez-Harguindeguy et al. 2013, Salgado-Negret et al. 2016), cuya área basal aumentó en el gradiente sucesional.
Estos cambios a lo largo de la sucesión han sido evidenciados mediante métodos destructivos por Vaccaro et al. (2003) en ecosistemas subtropicales húmedos en Venezuela, por Hughes et al. (2000) en bosques tropicales de México y Peña et al. (2011) mediante ecuaciones alométricas en bosques húmedos tropicales y muy húmedos premontanos en Colombia. Con lo que, considerando esta correspondencia, la abundancia y dominancia relativas de TFPs, y sus cambios a lo largo de la sucesión, podrían servir para inferir cambios en el secuestro biológico de carbono en ecosistemas en recuperación.
Comportamiento de la MPC de los rasgos del gradiente sucesional
La media ponderada de la comunidad (MPC) del área foliar específica no mostró variación a lo largo de la sucesión; mientras que para el área foliar y contenido foliar de materia seca se presentaron variaciones estadísticamente significativas entre dos estados sucesionales. En contraste, la MPC de la densidad de madera mostró un aumento consistente a lo largo del gradiente, adicional a una variabilidad reducida al interior de cada uno de los estados sucesionales evaluados, lo que indicaría que a nivel comunitario se presenta un cambio en la capacidad de almacenamiento de carbono por unidad de volumen a medida que se avanza en el gradiente. Esta tendencia fue también encontrada por Castellanos (2013) en los bosques secos tropicales del Caribe colombiano.
Las variaciones de este índice en ecosistemas secos, y su estrecha relación con el carbono almacenado, han llevado al incremento de estudios en los últimos años, en los que se ha encontrado que la MPC de la densidad de la madera se ha mostrado como un buen predictor del almacenamiento de carbono, tanto en metaanálisis (Van der Sande et al. 2017), como en sitios específicos como los bosques semiáridos del Chaco Argentino (Conti y Díaz 2013); por lo que se cuenta con evidencia para plantear que la densidad de la madera podría servir como un predictor de la capacidad de la vegetación de secuestrar carbono.
CONCLUSIONES
Este estudio buscó generar elementos para el análisis de la sucesión vegetal desde un enfoque funcional, particularmente ecosistemas con limitada investigación hasta el momento, como los bosques secos tropicales interandinos. Los TFPs asociados con estrategias adquisitivas (2 y 4) presentaron una disminución en su abundancia relativa a lo largo del gradiente sucesional; mientras el TFP 1, asociado a estrategias conservativas, aumentó su área basal pero no su abundancia. Esto, sumado al aumento de la MPC de la densidad de la madera, muestra un vínculo entre la variación funcional durante la sucesión y la disminución en la captura de carbono, acompañada de un mayor almacenamiento.
Se sugiere que futuros estudios analicen en detalle las relaciones encontradas entre la densidad de madera y la abundancia/ dominancia de TFPs con respecto al proceso de almacenamiento y captura de carbono.


