Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El bosque seco tropical (bs-T) es un ecosistema altamente dinámico que se desarrolla en regiones cálidas entre 0-1000 m de altitud, con clima estacional que incluye un período de sequía de 4-6 meses, temperatura promedio >24°Cy precipitación anual entre 700-2000 mm (IAvH 1995). En el Neotrópico se localiza el 67 % de los remanentes de bs-T a nivel global, y en Colombia solo persiste cerca del 3,7 % de bs-T original conformado por áreas relictuales en parches y corredores riparios estrechos (Corzo y Delgado 2012). Además, contiene muchas especies con distribución severamente restringida, alto grado de endemismo (Portillo-Quintero y Sánchez-Azofeifa 2010) y procesos de especiación.
El bs-T es considerado único como reserva de variabilidad genética, con alto valor estratégico para la conservación de la diversidad biológica (Pizano y García 2014), y ofrece servicios ecosistémicos (SE) como captura de carbono (Andrade-Castañaeda et al. 2017), apoyo a la productividad del suelo, ciclaje de nutrientes, producción de madera y combustible, de los que se tiene poca comprensión (Portillo-Quintero y Sánchez-Azofeifa 2010, Casanoves et al. 2011). A pesar de la importancia ecológica, el bs-T enfrenta graves problemas de pérdida y fragmentación de hábitat, con alta tasa de deforestación que genera paisajes fragmentados con pequeños parches aislados de bosque secundario, rodeados de áreas de cultivos y ganadería extensiva (Collinge 2009, Ballesteros 2015).
En las regiones secas de Colombia el establecimiento de sistemas de ganadería extensiva Convencional (SC) y áreas de cultivos, han contribuido a la transformación del paisaje, a acelerar los procesos de erosión del suelo y el deterioro de su biodiversidad (Pizano y García 2014); factores que han convertido al bs-T en uno de los ecosistemas más degradados, amenazados y menos conservados del Neotrópico (Rodríguez et al. 2009). En tanto que, en los sistemas de ganadería extensiva bajo manejo silvopastoril (SSP), se integran procesos bióticos y abióticos dinámicos, en una modalidad de agroforestería con combinaciones de especies vegetales-animales y, patrones de uso y manejo agroecológico que permite incrementar la productividad de forma sostenible y conservar la biodiversidad.
En este escenario, el establecimiento de ganadería extensiva en SSP dotados de fragmentos relictuales de bs-T, podría ampliar la cobertura vegetal e incrementar la conectividad funcional, posibilitando la supervivencia de las especies sensibles a la deforestación y pérdida de hábitat (Barrance et al. 2009). En este trabajo, se compara el efecto del sistema silvopastoril y sistema convencional de ganadería extensiva sobre la composición florística y estructura de fragmentos de bs-T, aportando información clave para entender mejor la dinámica y estado de conservación del bs-T en paisajes ganaderos del Caribe colombiano.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en fragmentos bs-T asociados a dos sistemas de manejo: silvopastoril (SSP) y convencional (SC) de ganadería extensiva en Córdoba, región Caribe de Colombia (Fig. 1). Esta región hace parte del cinturón árido peri-caribeño colombiano, con clima cálido tropical; temperatura promedio anual de 28 °C, precipitación promedio anual de 1300 mm con un patrón de distribución unimodal, con una época seca de diciembre-marzo y otra lluviosa de abril-noviembre (Palencia et al. 2006). El principal uso de la tierra es agricultura y ganadería extensiva, siendo esta región uno de los centros ganaderos más importantes de Colombia.
Recolección de datos
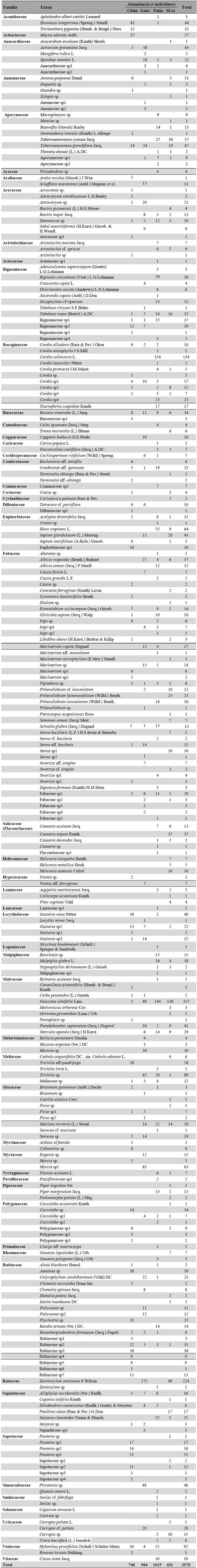
Durante agosto-octubre 2011 se realizaron muestreos en fragmentos de bs-T de cuatro localidades. Se evaluaron cuatro parcelas de 0,1 ha/localidad, distribuidas en forma aleatoria simple, siguiendo el método propuesto por Gentry (1982) con modificaciones de Villareal et al. (2004) para incluir plantas con DAP >1 cm. Se recolectaron muestras vegetales que fueron conservadas y procesadas. El material vegetal fue identificado mediante ayuda de claves taxonómicas, comparaciones con ejemplares depositados en herbarios y guías ilustradas de Mendoza et al. (2004) y las descripciones de Gentry (1995). Se censaron todos los individuos encontrados en cada unidad de muestreo (UP), y se midió: altura dosel, altura del fuste, altura y diámetro de la copa de los árboles, y estratificación vertical del bosque. Se registraron las características secundarias de las especies útiles en la identificación como presencia de flores, frutos, látex, exudado, olores y esencias vegetales. A los individuos censados con circunferencia a la altura del pecho (CAP) ≥ 3,14 cm, se les registraron los datos dasométricos de altura fustal, altura de copa y altura total, diámetro de copa y hábito de crecimiento (Mendoza-C. 1999). El material recolectado se depositó en el Herbario de la Universidad de Córdoba-Montería (HUC), y copias se enviaron al Herbario Joaquín Antonio Uribe, Medellín (JAUM) y al Herbario de la Pontificia Universidad Javeriana, Bogotá (HPUJ).
Análisis de la información
A partir de las medidas de CAP se determinó el DAP según Franco et al. (1997), y con los valores de DAP se calculó el área basal (AB) según Lema (2003). Se determinó la composición florística de la vegetación a partir del censo de los individuos con CAP ≥ 3,14 cm. Se construyeron curvas de acumulación de especies (Colwell et al. 2004), se calculó el índice de Shannon-Wiener y el inverso de Simpson (Magurran 2004). Se calculó la riqueza taxonómica mediante la diversidad alfa verdadera; y la tasa de recambio de especies entre fragmentos del SSP y SC se midió utilizando la diversidad beta verdadera (Jost 2006). En el análisis de la estructura vegetal se tuvieron en cuenta las especies de mayor área basal (AB). Se calculó el índice de valor de importancia de las especies (IVI), se evaluó importancia ecológica de las familias mediante el índice de valor de importancia de las familias (IVIF) según el método de Mori et al. (1983). Mediante el software PAST v. 2.17, se realizó un análisis de conglomerados para evidenciar el grado de disimilitud florística entre fragmentos asociados al SSP y SC, teniendo en cuenta el método jerarquico Unweighted Pair-Group Arithmetic Average (UPGMA) y el índice de Bray-Curtis (Magurran 2004). Se realizó un análisis inverso de Kaandorp (1986) para establecer la composición de las agrupaciones de cada localidad y determinar especies exclusivas que concentran el 100 % de su abundancia en un ensamblaje, especies características que concentran al menos el 65 % de su abundancia en un grupo y tiene una frecuencia mayor al 65 %, y especies generalistas que aparecen en todas las localidades o en la gran mayoría de ellos.
RESULTADOS Y DISCUSIÓN
Composición florística
Se registró un total de 3378 individuos, con un esfuerzo de muestreo que alcanzó cerca del 84 % estimado (Fig. 2). Se identificaron 252 taxones de plantas de 173 géneros y 55 familias. Las familias que presentaron la mayor riqueza taxonómica fueron Fabaceae (45), Rubiaceae (19), Bignoniaceae (12), Boraginaceae (12) y Moraceae (10). En términos de abundancia relativa fueron dominantes Malvaceae (11,66 %), Fabaceae (9,14 %), Rubiaceae (7,46 %), Boraginaceae (6,98 %) y Rutaceae (6,66 %). El género Cordia presentó mayor diversidad de taxones (10), seguido de Machaerium (6), Senna (5), Pithecellobium (4) y Casearia (4). El 73 % de los géneros estuvo representado por un taxon, 14 % por dos taxones y 8,5 % por tres taxones. Cerca del 60 % del total de los taxones registrados presentaron abundancias menores a cuatro individuos. En los fragmentos de bs-T del SSP, las familias Fabaceae, Boraginaceae y Apocynaceae presentaron mayor riqueza taxonómica; en tanto que, en fragmentos del SC Fabaceae y Rubiaceae presentaron mayor riqueza. Estos resultados concuerdan con estudios realizados en otras áreas de bs-T (Gentry 1995, IAvH 1998, Mendoza-C. 1999, Carrillo-Fajardo et al. 2007, Carbonó-Delahoz y García-Q. 2010, Coelho et al. 2012, Rodríguez M. et al. 2012, Olascuaga-Vargas et al. 2016), donde estas familias muestran una alta consistencia taxonómica, con una composición vegetal característica de este tipo de ecosistemas en el Neotrópico y en Colombia.
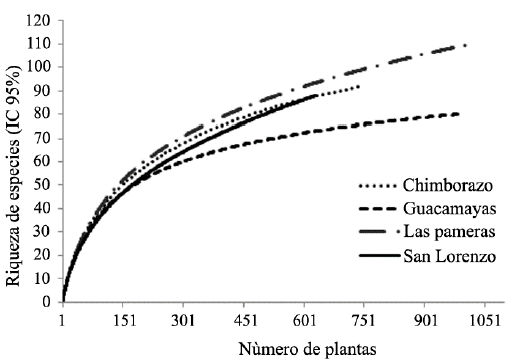
Figura 2 Curvas de acumulación de especies de plantas vasculares en los cuatro fragmentos de bs-T inmersos en matrices de ganadería extensiva en Córdoba, Colombia.
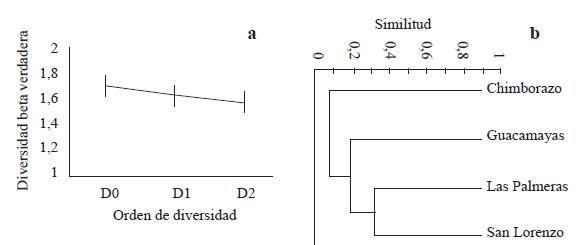
Figura 3 Análisis de diversidad y similitud taxonómica. a. Diversidad beta verdadera de vegetación entre fragmentos de bs-T en paisaje de ganadería extensiva bajo SSP y SC; se presentan tres medidas de diversidad beta con q (0, 1 y 2), con IC de 95 %; b. Análisis de agrupamiento con el coeficiente de similitud de Bray-Curtis entre los fragmentos asociados al SSP y SC de ganadería extensiva en Córdoba, Colombia.
En los fragmentos de bs-T del SSP, con 1646 registros se identificaron 161 taxones de plantas, donde las familias Malvaceae (19 %), Boraginaceae (12,3 %) y Fabaceae (10,8 %) fueron dominantes. En la localidad de Las Palmeras con 110 taxones registrados, Guazuma ulmifolia Lam., Cordia collococca L., Hura crepitans L. y Hybanthusprunifolius (Scult.) Schulze-Menz presentaron la mayor abundancia relativa; y 66 especies presentaron abundancias menores a cuatro individuos. En tanto que, en San Lorenzo con 88 taxones identificados, Guazuma ulmifolia, Zanthoxylum setulosum P. Wilson y Casearia arguta Kunth., registran la mayor abundancia relativa (Anexo en Material suplementario).
En los fragmentos del SC, con 1732 registros se identificaron 144 taxones de plantas vasculares siendo dominados por las familias Rubiaceae (13,2 %), Rutaceae (10,1 %) y Fabaceae (7,6 %). En Chimborazo se registraron 93 taxones con mayor abundancia relativa de Trichilia aff. quadrijuga, Pouteria sp., y Bravaisia integerima (Spreng.) Standl., y 56 especies presentaron abundancias con menos de cuatro individuos, muchas de las cuales son especies pioneras, tal como lo reporta Vargas y Ramírez (2014) en bs-T. En Guacamayas con 80 taxones registrados, Zanthoxylum setulosum, Myrcia sp1., Trichilia sp., Picramnia sp., y Guazuma ulmifolia fueron más abundantes, y 36 especies presentaron abundancias con menos de cuatro individuos (Anexo en Material suplementario). La diversidad en la composición taxonómica entre fragmentos de bs-T asociados al SSP y SC fue significativamente diferente (Shannon test, P < 0,0001, df = 3092,1, t = 5,03). Esta diversidad y abundancia taxonómica es similar a otros fragmentos de bs-T en la Región Caribe colombiana (Marulanda et al. 2003, Rodríguez M. et al. 2012, Olascuaga-Vargas et al. 2016, Sanmartín-Sierra et al. 2016, Herazo Vitola et al. 2017), y lo encontrado puede ser resultado de la estructura horizontal de la vegetación en forma de mosaico en diferentes estados de sucesión ecológica, lo cual puede ser producto del tipo de manejo del sistema productivo, lo cual concuerda con el trabajo de García et al. (2014), que reportan también alta diversidad taxonómica y muchas especies raras.
La diversidad alfa verdadera (°D), fue significativamente mayor en los fragmentos del SSP con un nivel de completitud > 97 %, con una riqueza de 17 especies efectivas más que en los fragmentos del SC (Tabla 1).
Tabla 1 Diversidad alfa verdadera de plantas por localidades y sistemas de manejo en paisajes de ganadería extensiva en Córdoba. Se presentan tres medidas de diversidad: 0 D, 1 D, 2 D; y la completitud del muestreo, con un nivel de confianza del 95 %.
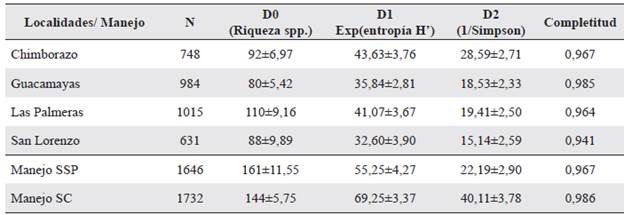
Estos índices de diversidad alfa verdadera son similares a los valores reportados por Lebrija-Trejos et al. (2008) para otras áreas de bosque seco. Tal diferencia puede explicarse por la formación de mosaicos paisajísticos con diferentes etapas de sucesión como consecuencia del tipo de manejo, grado de aislamiento, efecto de borde, frecuencia y duración del ganado dentro de los fragmentos, historia de uso de la tierra y el grado de conservación de los fragmentos (Ballesteros 2015), entre otros, características que no se evaluaron en este trabajo, pero que pueden estar incidiendo en la composición y estructura de la vegetación. La diversidad beta verdadera o tasa de recambio de especies entre fragmentos del SSP y fragmentos del SC fue del 84,5 % (0D = 1,69 comunidades efectivas), con un solapamiento de especies del 30,8 % (Fig. 3a). Una similitud del 20% en la composición taxonómica entre fragmentos del SSP y SC (Fig. 3b), sugieren que la composición vegetal entre fragmentos de bs-T es diferente, lo cual depende fundamentalmente de la historia de uso y manejo del sistema. Este resultado puede ser útil en el diseño de programas de conservación, en la medidad que cada fragmento alberga un importante componente de la diversidad vegetal, lo cual es importante conservar. Esta condición en la composición florística de los fragmentos de bs-T evaluados representa un aporte significativo para la conservación de la diversidad regional de la Región Caribe colombiana.
Estructura de la vegetación
El índice de valor de importancia por especie (IVI) de plantas en los fragmentos del SSP fue mayor para Bactris major (40,8), Guazuma ulmifolia (29,4), Hura crepitans (11,9) y Cordia collococca (10,4), especies que presentan un IVI mayor que los reportados por Olacuaga et al. (2016) en áreas de bs-T en otras áreas de la Región Caribe. En los fragmentos del SC las especies Cavanillesia platanifolia (Humb. & Bonpl.) Kunth (31,9), Ceiba pentandra (L) Gaertn. (28), Astrocaryum sp. (16,5) y Lecithys minor Jacq. (11,7) tuvieron el mayor IVI (Fig. 4). El índice de valor de importancia para las familias (IVIF) en los fragmentos del SSP fue mayor para Fabaceae (57,6), Malvaceae (46,5), Malpighiaceae (20.8) y Boraginaceae (20,4); mientras que, en los fragmentos del SC, Fabaceae (42.9) , Arecaceae (23,8), Apocynaceae (18,8) y Euphorbiaceae (14,2) presentaron mayor IVIF (Fig. 5). Estos resultados son concordantes con los reportes de familias de plantas para ecosistemas secos tropicales (Gentry 1995, Mendoza-C. 1999, Albesiano y Rangel-Ch 2006, Carrillo-Fajardo et al. 2007, Carbonó-Delahoz y García-Q. 2010, Rodríguez M. et al. 2012, Berdugo-Lattke y Rangel-Ch 2015, Villanueva et al. 2015, Dryflor et al. 2016, Sanmartín-Sierra et al. 2016, Olascuaga-Vargas et al. 2016, Herazo Vitola et al. 2017).
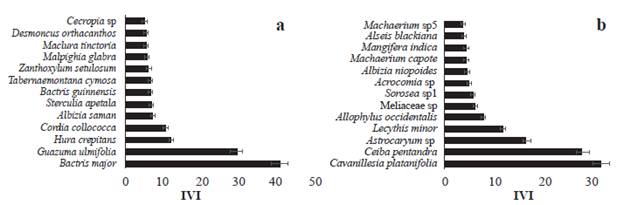
Figura 4 Índice de valor de importancia (IVI) por especies vegetales (DAP >1 cm, cuatro parcelas de 0,1 ha) en fragmentos de bs-T en matrices de ganadería extensiva con, a. sistema silvopastoril y b. sistema convencional.
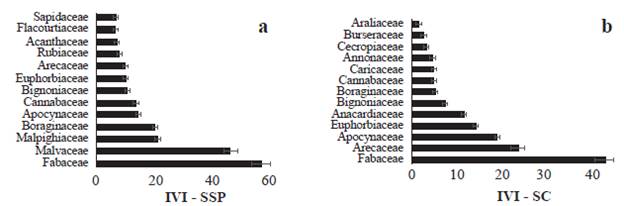
Figura 5 Índice de valor de importancia (IVIF) por familias (DAP >2,5 cm en cuatro parcelas de 0,1 ha) en fragmentos de bs-T en matrices de ganadería extensiva con a. sistema silvopastoril, y b. sistema convencional.
El análisis inverso de Kaandorp (Fig. 6) permitió identificar 158 especies exclusivas, distribuidas en cuatro grupos: I: Guacamayas (S = 28), II: Chimborazo (S = 40), III: Las Palmeras (S = 51), y IV: S. Lorenzo (S = 39). Se registraron siete especies generalistas para el conjunto I-II-III-IV donde Guazuma ulmifolia (Malvaceae) y Bursera simaruba (L.) Sarg. (Burseraceae) presentaron la mayor dominancia. El conjunto I-III-IV, presentó seis especies generalistas Maclura tinctoria (L.) Stend. (Moraceae) y Sterculia apetala (Jacq.) H. Karst. (Malvaceae) las más abundantes. De este último conjunto se desprende un subconjunto III-IV con 20 especies generalistas, donde Enterolobium cyclocarpum (Jacq.) Griseb. (Fabaceae), Cecropia sp. (Urticaceae) y Hura crepitans (Euphorbiaceae) fueron las especies más abundantes. Estos resultados confirman la alta heterogeneidad de los fragmentos evaluados, dado que solo comparten el 3,64 % de las especies registradas. Dada la pobre representación de los bs-T en áreas protegidas, cobra especial importancia la conservación de los fragmentos que persisten en las áreas de Reserva de la Sociedad Civil en los paisajes ganaderos de la región Caribe colombiana.
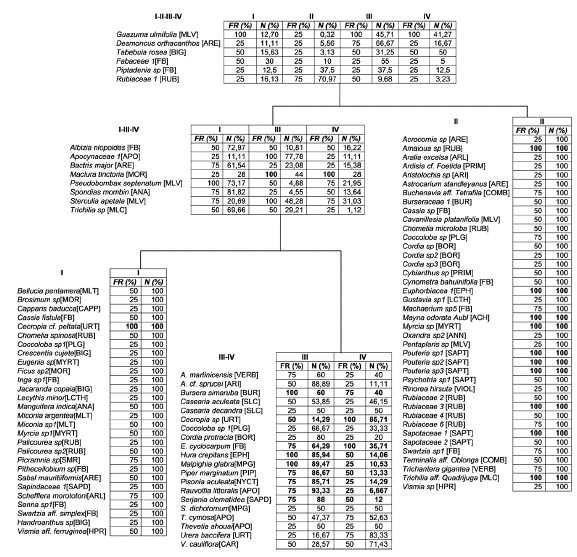
Figura 6 Análisis inverso de Kaandorp de los ensamblajes plantas vasculares en los fragmentos de bs-T Guacamayas (I), Chimborazo (II), Palmeras (III) y San Lorenzo (IV), Porcentaje del promedio de abundancia total (% N) y porcentaje de la frecuencia por ensamblaje (% Fre). Achariaceae (ACH), Acanthaceae (AC), Anacardiaceae (ANA), Annonaceae (ANN), Apocynaceae (APO), Araceae (ARA), Araliaceae (ARL), Arecaceae (ARE), Aristolochiaceae (ARI), Asteraceae (AST), Bignoniaceae (BIG), Boraginaceae(BORA), Burseraceae (BUR), Cannabaceae (CANN), Capparaceae (CAPP), Caricaceae (CAR), Cochiospermaceae (CCHL), Combretaceae (COMB), Connaraceae (CNN), Costaceae (CST), Cyclanthaceae (CYCLAN), Dilleniaceae (DLL), Euphorbiaceae (EPH), Fabaceae (FB), Flacourtiaceae (FLAC), Heliconiaceae (HCL), Hypericaceae (HPR), Lamiaceae (LMC), Lauraceae (LRC), Lecythidaceae (LCTH), Logoniaceae (LOG), Malpighiaceae (MPG), Malvaceae (MLV), Melastomataceae (MLT), Meliaceae (MLC), Moraceae (MOR), Myrsinaceae (MRS), Myrtaceae (MYRT), Nyctaginaceae (NYCT), Passifloraceae (PSS), Piperaceae (PIP), Polygonaceae (PLG), Primulaceae (PRIM), Rhamnaceae (RHN), Rubiaceae (RUB), Rutaceae (RUT), Sapindaceae (SAPD), Sapotaceae (SAPT), Simaroubaceae (SMR), Smilacaceae (SMLC), Solanaceae (SOL), Urticaceae (URT), Vervenaceae (VERV), Violaceae (VIOL), Vitaceae (VIT).
CONCLUSIONES
La alta diversidad de taxones de plantas encontradas y porcentaje de especies exclusivas, indican la gran importancia que tienen estos fragmentos para la conservación de la biodiversidad regional. Los fragmentos de bs-T en matrices SSP de ganadería extensiva presentan mayor diversidad vegetal frente a los fragmentos del SC, y la composición de especies depende de las actividades antrópicas y la manera como usan estos ecosistemas.