











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Los bosques tropicales albergan la mayor diversidad de especies de plantas en el mundo (Gentry 1982, Balslev et al. 1998, Antonelli y Sanmartín 2011). El Chocó es una de las regiones con mayor riqueza de especies de plantas ('biodiversity hotspot' sensu Myers et al. 2000), alto grado de endemismo (Gentry 1986, 1993, McNeely et al. 1990, Antonelli y Sanmartín 2011) y tiene una gran fracción de plantas sin descubrir (Joppa et al. 2011).
Las palmas son un grupo icónico de los bosques tropicales. Su distribución abarca todo el trópico donde a menudo son muy diversas y abundantes. Las palmas presentan diferentes formas de vida, desde muy pequeñas en el sotobosque hasta muy altas alcanzando el dosel (Balslev et al. 2016). Las palmas están bien representadas en el Chocó con 122 especies de las cuales 32 son endémicas (Balslev et al. 2015). Son la segunda familia de plantas más utilizadas, después de Lauraceae, en la costa pacífica colombiana (Galeano 2000) y son de gran importancia económica para los habitantes del Chocó, ya que sus productos son comercializados en mercados regionales, nacionales e internacionales (Brokamp et al. 2011, Valois-Cuesta et al. 2013, Ledezma-Rentería y Galeano 2014, Cámara-Leret et al. 2016, Ledezma-Rentería et al. 2016). Por su riqueza, diversidad, importancia socio-económica y función ecológica, es esencial evaluar los patrones de riqueza de las palmas en el Chocó y analizar sus determinantes ambientales.
La región del Chocó se caracteriza por presentar una elevada precipitación, hasta 11 000 mm anuales en el centro del departamento del Chocó y disminuye hacia los extremos norte y sur, donde ronda los 2000-3000 mm anuales (Eslava 1992, Poveda et al. 2004).
Numerosos autores han confirmado que la diversidad de especies de plantas en los bosques tropicales aumenta con la precipitación y disminuye con la estacionalidad (Gentry 1982, 1988, Wright 1992, Specht y Specht 1993, Clinebell-H et al. 1995, Aplet et al. 1998). En una revisión de literatura se encontró que en 82 de 85 casos estudiados, la precipitación y la temperatura explican el 60 % de la variabilidad en la riqueza de especies, y la precipitación es el factor dominante en latitudes tropicales, mientras que la temperatura y la precipitación controlaban la riqueza de especies en zonas árticas y templadas (Francis y Currie 2003, Hawkins et al. 2003, Bjorholm et al. 2005).
En el trópico la relación entre la precipitación y la riqueza de especies de palmas ha sido mayormente estudiada a escalas continentales y con datos de presencia-ausencia. En América, África y Madagascar la precipitación es el factor que mejor explica la riqueza de palmas a escalas continentales (Bjorholm et al. 2005, Blach-Overgaard et al. 2010, Rako-toarinivo et al. 2013). En Tailandia, la precipitación es el segundo factor más importante después de la latitud en explicar la diversidad y distribución de especies de palmas (Tovaranonte et al. 2015). En la Península de Yucatán Mexicana la riqueza de palmas fue asociada positivamente con la precipitación, pero no con la abundancia (Alvarado-Segura et al. 2012). En el Chocó, Ramírez y Galeano (2011) estudiaron a escala regional la relación entre la precipitación y la riqueza de palmas, y plantearon la hipótesis que la riqueza aumenta hacia el centro de la región, donde la precipitación es mayor. En su investigación, muestrearon dos localidades (Tutunendo y Angostura) en el centro del departamento del Chocó colombiano, y registraron una mayor riqueza de palmas en Tutunendo (10 000 mm) que en Angostura (8000 mm).
Nuestro objetivo fue explorar la relación entre la riqueza y abundancia de palmas con la precipitación a escalas regionales en el Chocó biogeográfico. Abordamos este objetivo utilizando datos florísticos y ambientales obtenidos en 48 transectos en Colombia y Ecuador. A diferencia de investigaciones previas a escalas continentales y basadas en datos de potencial de especies, en este estudio exploramos los patrones a escalas regionales con datos de abundancia y riqueza medidos directamente en muestreos de campo. Mientras que estudios anteriores (e.g. Ramírez y Galeano 2011) incluyen una pequeña fracción del gradiente de precipitación (8000-10 000 mm) y de diversidad florística (33 especies), en este estudio abarcamos un gradiente de precipitación mayor (2000-10 000 mm) e incluimos 57 especies, aproximadamente 50 % de la flora de palmas del Chocó.
MATERIALES Y MÉTODOS
Área de estudio
El Chocó biogeográfico se extiende desde el extremo sur de Panamá hasta el norte de Ecuador, y cubre aproximadamente 71 000 km2. Presenta una alta precipitación que alcanza ~11 000 mm en el centro del departamento del Chocó (Colombia) y disminuye hasta 2000-3000 mm en los extremos norte y sur. La temperatura media anual es de 27 °C y la humedad relativa es 90 % (Eslava 1992, Poveda et al. 2004). La investigación se realizó en Colombia en el departamento de Chocó en la región de Capurganá, donde la precipitación media anual oscila entre 2000-2983 mm, en Puerto Pervél (7127 mm), en el Valle del Cauca en la región de Bahía Málaga (7622 mm) y en Ecuador en la provincia de Santo Domingo de los Tsáchilas, en Santo Domingo (3340 mm) (Fig. 1).
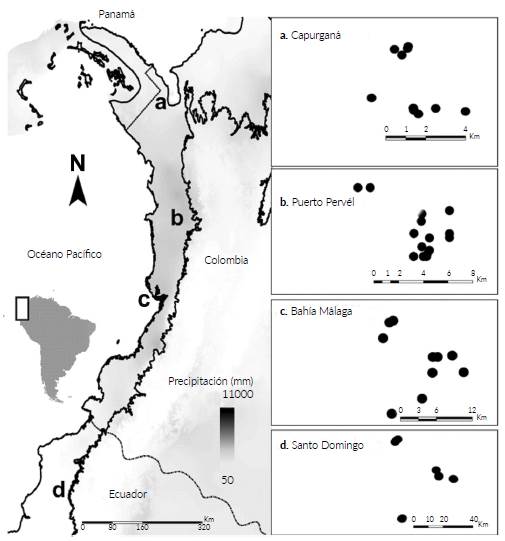
Figura 1 Localización de los 48 transectos muestreados en cuatro regiones del Chocó biogeográfico. Las cajillas a la derecha muestran la distancia entre cada uno de los transectos realizados en cada región.
Los datos de precipitación anual (en mm) fueron obtenidos de la base de datos de WorldClim Versión 2, a resolución espacial de 30 segundos (~ 1 km2) (Fick y Hijmans 2017).
Toma de Datos
Entre 2011-2017 se tomaron datos de distribución y abundancia de especies de palmas a lo largo del Chocó bio-geográfico (Fig. 1). Se utilizaron imágenes satelitales para establecer los 48 transectos en bosques de tierras bajas no inundables sin influencia humana. Cada transecto tuvo un tamaño de 5x500 m y fue dividido en 100 subunidades de 5x5 m. Los transectos fueron ubicados estratificadamente a una distancia de 500 m del borde de los caminos o de los ríos, siguiendo la metodología de Balslev et al. (2010). Se registró la localización geográfica en el punto medio de cada transecto con un GPS de mano según el sistema de coordenadas Universal Transversal Mercator (UTM). En cada subunidad se identificaron y contabilizaron todos los individuos presentes de todos los estados de crecimiento (plántulas, juveniles, subadultos y adultos). Se recolectaron muestras de algunas de las especies presentes dentro de los transectos y estas fueron depositadas en los herbarios del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia sede Bogotá (COL), Universidad Tecnológica del Chocó (CHOCO), Pontificia Universidad Católica del Ecuador (QCA) y de la Universidad Nacional de Colombia sede Medellín (MEDEL).
Análisis
Se cuantificó la riqueza de especies de palmas con el alfa de Fisher, definido por la fórmula:
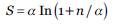
donde S es el número de especies, n el número de individuos y α el alfa de Fisher. Este índice es ampliamente utilizado porque es casi insensible al tamaño de muestra y discrimina bien entre comunidades (Magurran 2004, Colwell 2009). Se utilizó el programa PAST versión 3.1 (Hammer et al. 2001). Se realizó una regresión lineal simple para observar la relación entre la precipitación, el número de especies (S), número de individuos y diversidad de palmas.
Las distancias geográficas fueron obtenidas en campo mediante un GPS de mano, y se calculó una matriz de distancias geográficas utilizando el índice euclidiano transformado en logaritmo en base 10. Realizamos un test de Mantel para evaluar si la disimilitud en la composición florística está correlacionada con la disimilitud en precipitación. Para controlar la autocorrelación espacial y así evitar inflar la importancia de las variables ambientales, realizamos un test parcial de Mantel, por el cual la correlación con las distancias geográficas transformadas logarítmicamente se controló antes de calcular la correlación entre las diferencias florísticas y ambientales. Las pruebas de Mantel y Mantel parcial se realizaron con datos de presencia-ausencia y con datos de abundancia. Para ambos, se utilizó la forma estandarizada de la estadística de Mantel (rM), que es equivalente al coeficiente de correlación de Pearson entre dos matrices de disimilitud. En todos los casos, se estableció la significancia estadística de cada correlación en P < 0,001 con una prueba de Monte Carlo con 999 permutaciones aleatorias. Se utilizó el paquete vegan (Oksanen et al. c2016), del programa estadístico R versión 3.3.2. (R Core Team c2017).
RESULTADOS
Riqueza
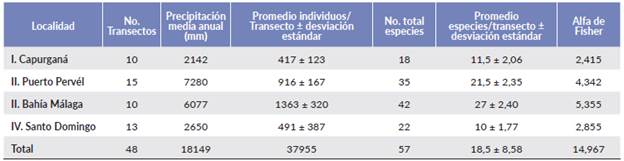
Se registraron 57 especies y 37 955 individuos de palmas en los 48 transectos que cubren en total doce hectáreas (Tabla 1). Las regiones del Chocó central presentaron un mayor número de especies y mayor riqueza (S) que las regiones de los extremos norte y sur (Tabla 1).
Abundancia
Bahía Málaga fue la región con mayor abundancia de palmas (promedio = 1363 individuos por transecto), seguida de Puerto Pervél (916 individuos), correspondiendo a las regiones centrales del Chocó (Tabla 1). Las especies más abundantes fueron Wettinia quinaria (O.F. Cook & Doyle) Burret (124 ± 162), Oenocarpus bataua Mart. (123 ± 207), Welfia regia H. Wendl. ex Andres (87 ± 217), Euterpe oleracea Mart. (43 ± 177) y Synechanthus warscewiczianus H.Wendl. (40 ± 90) (Tabla 2). Socratea exorrhiza H. Wendl. fue la especie más frecuente (45 de los 48 transectos), seguida de Geonoma cuneata H. Wendl. ex Spruce (43), Oenocarpus bataua y Wettinia quinaria (ambas 30).
Las palmas más abundantes son representativas de las especies más altas como es el caso de Oenocarpus bataua y Socratea exorrhiza (Fig. 2), pero también están representadas por algunas palmas pequeñas de sotobosque como Geonoma cuneata (Fig. 3).

Figura 2 Algunas de las especies más abundantes del dosel de los bosques del Chocó a. Wettina quinaria; b. Oenocarpus bataua; c. Welfia regia; d. Euterpe oleracea, e. Socratea exorrhiza y f. Attalea colenda.

Figura 3 Algunas de las especies más abundantes de sotobosque a. Calyptrogyne costatifrons; b. Aiphanes macroloba; c. Synechanthus warscewiczianus; d. Chamaedorea deneversiana; e. Bactris chocoensis (Bernal et al. 2014) y g. Geonoma cuneata.
Relación entre precipitación, riqueza y abundancia
Se encontró una correlación positiva entre la precipitación y la diversidad de especies (r2 = 0,84; P < 0,001; n = 48), (Fig. 4a). De igual forma, la precipitación presentó una fuerte correlación positiva con la abundancia de palmas (r2 = 0,53; P < 0,001; n = 48) (Fig. 4b). Al igual que la diversidad y la abundancia, encontramos una mayor riqueza (alfa de Fisher) en la zona central del Chocó donde la precipitación es mayor (r2 = 0,78; P < 0,001; n = 48) (Fig. 4c). Según la prueba de Mantel, existen correlaciones positivas entre la composición florística y la precipitación tanto al analizar datos de presencia-ausencia (rM = 0,88; P = 0,001; n = 48) como de abundancia (rM = 0,79; P = 0,001; n = 48). Se encontró una correlación positiva entre la distancia geográfica y la composición de las comunidades de palmas en el Chocó, para datos de presencia-ausencia (rM = 0,63; P = 0,001; n = 48) como para datos de abundancia (rM = 0,57; P = 0,001; n = 48). Después de controlar el efecto de la distancia geográfica con la prueba de Mantel parcial, todas las correlaciones entre la composición florística y la precipitación se mantuvieron estadísticamente significativas tanto para datos de presencia-ausencia (rM = 0,80; P = 0,001; n = 48) como de abundancia (rM = 0,67; P = 0,001; n = 48).
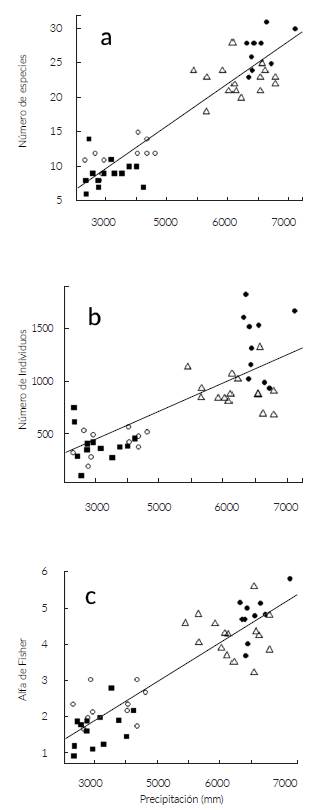
Figura 4 Relación entre precipitación y a. número de especies; b. abundancia y c. diversidad de palmas (Alfa de Fisher) en 48 transectos en el Chocó. Círculos ,ellenos = Bahía Málaga (Chocó central), triángulos = Puerto Pervél (Chocó central), círculos sin relleno = Capurganá (Chocó noreste), cuadros rellenos = Santo Domingo (Chocó sur).
DISCUSIÓN
Riqueza
Los resultados de esta investigación (57 especies en doce hectáreas) corroboran que el Chocó es una de las regiones más ricas en especies de palmas (Balslev et al. 2015). A escala regional, Ramírez y Galeano (2011) registraron 33 especies de palmas en 2,5 ha, Gentry (1986) registró 17 especies en 0,1 ha y Browne y Karubian (2016) registraron 23 especies en 5,5 ha. En nuestro estudio, los géneros con mayor número de especies corresponden a palmas de sotobosque (Geonoma con diez especies y Bactris con ocho) que son también el grupo más rico en especies en la serranía del Darién (Cano et al. 2017) y en el occidente Amazónico (Balslev et al. 2017, Cámara-Leret et al. 2017a).
La alta riqueza de especies de palmas a escalas continentales y regionales está muy ligada a factores ambientales (Eiserhardt et al. 2011). La correlación positiva encontrada entre diversidad de palmas y precipitación a escala regional corrobora lo encontrado en estudios anteriores en América, África y Madagascar (Bjorholm et al. 2005, Blach-Overgaard et al. 2010, Eiserhardt et al. 2011, Alvarado-Segura et al. 2012, Rakotoarinivo et al. 2013, Tovaranonte et al. 2015). Esto confirma que la riqueza de especies de palmas incrementa con la precipitación a escalas continentales y regionales.
Abundancia
La abundancia de palmas disminuyó desde el centro del Chocó hacia los extremos y estuvo correlacionada con la precipitación. Una explicación a este patrón podría ser que las especies más abundantes (Euterpe oleracea, Euterpe precatoria, Oenocarpus bataua, Socratea exhorriza, Wettinia quinaria y Welfia regia) son especies medianas-altas adaptadas para sobrevivir en ambientes muy húmedos que presentan menor densidad en lugares con menor precipitación. Cuando estas especies se excluyen de los análisis los valores en la regresión lineal cambian (r 2 = 0,08; P = 0,04; n = 48) (Tabla 2).
Tabla 2 Especies registradas en los 48 transectos y sus abundancias en las cuatro regiones estudiadas del Chocó
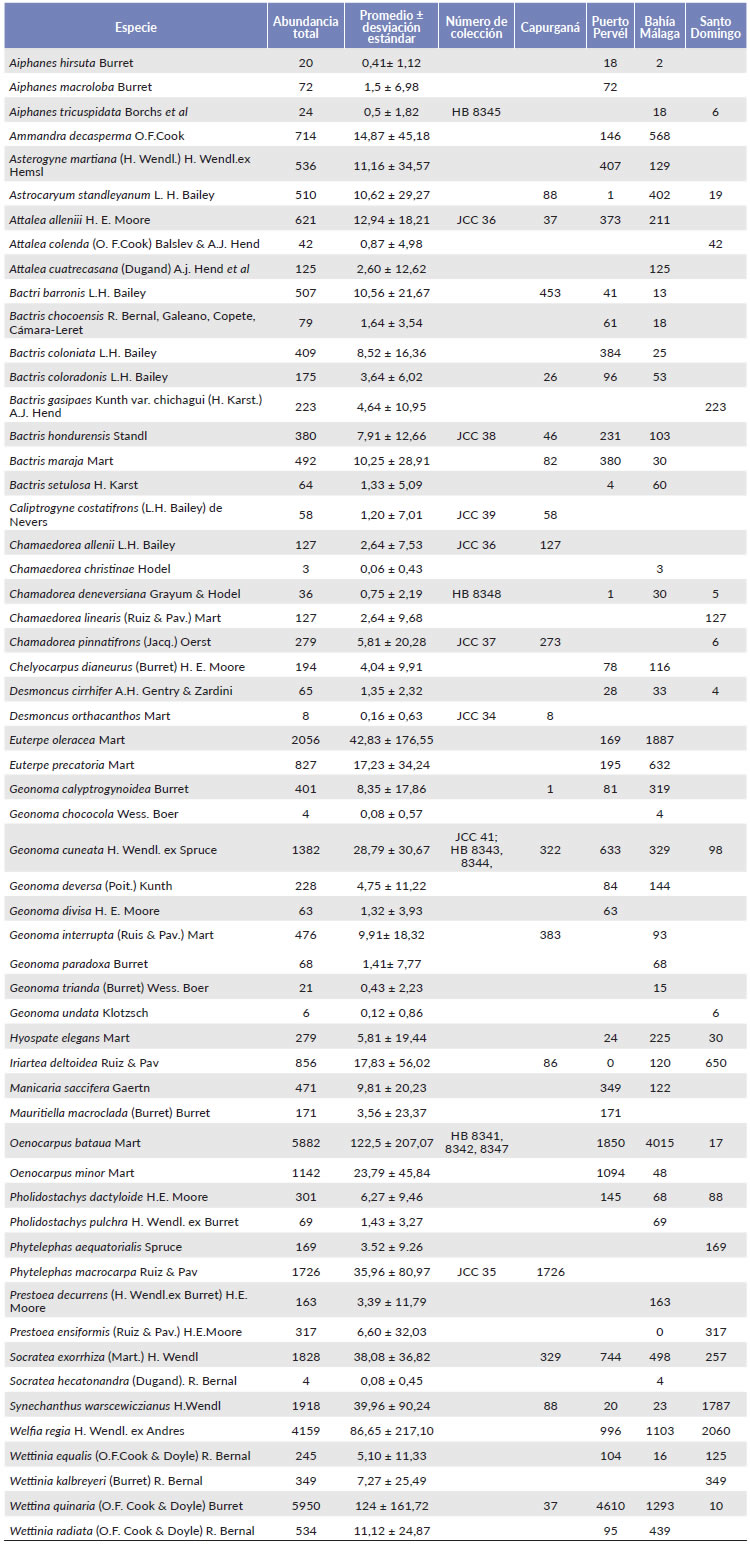
Las especies de palmas más abundantes en el Chocó central son diferentes a las encontradas en los extremos norte y sur. Por ejemplo, Wettinia quinaria es más abundante en el centro que en el norte/sur del Chocó posiblemente porque es una especie más tolerante a los altos niveles de precipitación. En cambio, las especies de Chamaedorea son más abundantes en el extremo norte que en el centro y que en el sur. Esto puede ser explicado por la historia biogeográfica del género, cuyo centro de diversificación es Centroamérica (Hodel 1992, Galeano y Bernal 2010). Las especies de palmas más abundantes en este trabajo (Wettinia quinaria y Oenocarpus bataua) son similares a las registradas en otros inventarios en el Chocó (Faber-Langendoen y Gentry 1991, Galeano 2001, Asprilla et al. 2003, Rangel 2004, Mosquera et al. 2007, Ramírez y Galeano 2011, Torres-Torres et al. 2016) y se destacan por su importancia etnobotánica en el noroeste suramerica-no (Cámara-Leret et al. 2017b). En el Chocó, los tallos de W. quinaria se usan ampliamente en la construcción de casas, mientras que los frutos de O. bataua se valoran para la alimentación humana (Cámara-Leret et al. 2016). Futuros estudios enfocados a mejorar nuestra comprensión sobre el nicho climático-edáfico de las especies de palmas del Chocó, su respuesta a los cambios climáticos (Loarie et al. 2009), y la relación entre la abundancia y la influencia humana (Levis et al. 2017) serán importantes para desarrollar estrategias para minimizar la pérdida de biodiversidad en este punto caliente de biodiversidad.


