











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Palms are one of the most used plant groups by humans (Johnson 1998). These plants are characterized by great diversity and abundance, with approx. 1500 species and 200 genera worldwide (Henderson et al. 1995). Colombia has one of the highest palm species diversities in the world (Galeano and Bernal 2010). Attalea nucífera H.Karst. is an endemic palm of Colombia, with low-density across its area of distribution (Galeano and Bernal 2010). Its presence is known only in the mid- and low-valley of the Magdalena river, in Bolívar, Santander, Antioquia and Cundinamarca departments, between 200 - 1200 m (Galeano et al. 2015, Prada-Ríos and García 2019). According to Galeano and Bernal (2005) and Prada-Ríos and García (2019), this species has been characterized as Vulnerable (VU), as it is present in ten localities in which there was a decrease in habitat quality. Attalea nucífera grows in forest fragments along stream margins that are strongly altered by extensive cattle ranching and African oil palm (Elaeis guineensis Jacq.) cultivation (Prada-Ríos and García 2019). In the past, the inhabitants of the locality of Guaduas (Cundinamarca) consume the palm seeds, although, their consumption has decreased, and they rarely use the palm today (Prada-Ríos and García 2019).
Habitat loss has posted the highest threat to Colombian palms. Most of the Colombian species inhabit forest areas and have a low tolerance to deforestation. If the adult plants manage to survive and produce seed, then, the more vulnerable seedling cannot survive in open areas (Galeano and Bernal 2005). The primary cause of deforestation in the Andean valleys is the conversion of forest areas into cattle pastures (Pedraza Alvarez and Molina Portuguez 2007). Notably, African oil palm (Elaeis guineensis) crops have extended rapidly across the mid-Magdalena valley ecosystems, compromising wetlands and other areas that are strategic for wildlife conservation plans (Palacio-Lozano et al. 2008).
The study of population dynamics in natural habitats is essential for the conservation of rare or threatened species, and for establishing appropriate sustainable management plans (Sánchez-Velásquez et al. 2002, Chien et al. 2008, Martínez et al. 2010). In a large number of palms, the typical distribution of sizes has an inverted J shape characterized by a high number of seedlings and a decreasing number of juveniles and adults. Thus, a population with a different size distribution can be under stress because of the transformation of its habitat, or because of improper harvesting (Galeano et al. 2010).
Studies about the dynamics of populations are also an essential tool for the evaluation of the probability of growth of wild populations (Ticktin et al. 2002). Traditionally, the matrix population models and the elasticity analysis are the most widely used for this purpose (Caswell 2001, Schmidt et al. 2011). These models allow the projection of the population growth under different theoretical scenarios, to evaluate the contribution of different demographic processes occurring during the life cycle (Silvertown et al. 1993). For example, the analysis of elasticity can show which size categories are the most important for the maintenance of the population (Caswell 2000). Results of matrix population models have been used to assess the effects of harvesting in non-timber forest products or the effects of management measures in rare, endangered, or even invasive species (Schmidt et al. 2011). Other applications of demographic studies in conservation biology are the estimation of the probability of extinction for threatened species (Benton and Grant 1999).
In the present paper, we describe the results of a population study of A. nucifera in six localities of the valley of the Magdalena river, in Colombia. Specifically, we asked the following research questions: What is the structure and density of the A. nucifera populations? How is the population dynamics of A. nucifera under altered habitat conditions? What is the population behavior and what are the extinction probabilities under different management scenarios?
MATERIALS AND METHODS
Study species
Attalea nucifera is a palm with a unique and underground stem (acaulescent). The adults have 7-13 leaves of approx. 8-meter long, with 9-120 pines in each side, regularly distributed in the same plane of direction. The inflorescences are interfoliated with numerous rachillas that can reach 19 cm long. The male flowers are arranged in two rows in the exterior of the rachillas and can reach 2 cm long. The female inflorescences have the flowers arranged directly in the rachis. The fruits are elliptical to almost spherical (Galeano and Bernal 2010).
Study area
The present study was executed in six localities of the mid-Magdalena valley (Colombia), in the departments of Cundinamarca and Santander (Table 1). The mid-valley of the Magdalena river is located between the Central and Eastern Andean Mountain Chains (Mojica and Franco 1990). These areas have a tropical humid climate with a bimodal rain pattern and an average yearly rainfall of 3000 mm (Mojica et al. 2006). The average temperature is between 27 and 29 °C; some natural forest relicts are present, mostly of the Tropical Humid type (Holdridge 1987). These forest relics have been transformed for agricultural and cattle raising activities (Pedraza Alvarez and Molina Portuguez 2007). The altitudinal sampling range varied from 150 - 1200 m.
Table 1 Description of localities studied for Attalea nucífera in its area of distribution in the Magdalena River valley, Colombia.

The census for the demographic study was carried out in the forest relicts of Cucharal Stream, within the small village of Cucharal (VC), located in the municipality of Guaduas (Cundinamarca). This locality (VC) has an undulating topography with extensive areas dedicated to cattle raising and small agriculture, and with some forest relicts in the small river banks. The locality of VC corresponds to Dry Tropical Forest (Holdridge 1987), with an average altitude of 900 m. It has pastures, secondary high vegetation, and gallery forest (IDEAM 2010).
Population sampling
In each locality (Table 1), we established sampling transects of 20 m x 5 m (0.01 ha), keeping a distance of 20 m between transects. Depending on the population size of each locality, between two and 28 transects were established (Table 1). Each transect was distributed longitudinally, following the direction of the river located in the patch. In each transect, the total number of individuals was registered. The following morphological and reproductive parameters were recorded: Type of leaf (bifid or pinnate), number of principal veins of the youngest leaves (in the right side of the rachis), and the presence or not of reproductive structures (inflorescences or infrutescences). According to these parameters, the population was classified into six size categories from seedling (P) to mature adult (A2) (Table 2).
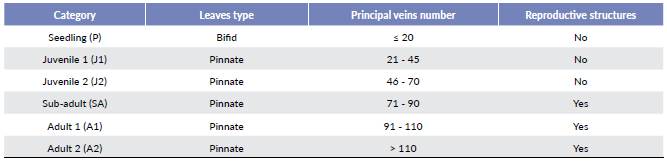
The census for the population dynamic study was done in February 2016 and February 2017 in the locality VC. A sample of 489 individuals of A. nucífera, 362 from the inner part of the transects, and 127 from the outer side, were marked according to the technique recommended by Galeano et al. (2010). We registered the number of principal veins in the youngest leaves (right side of the rachis from the opposite side), presence or absence of reproductive structures, number of dead individuals (mortality), and number of seedlings (recruitment) inside of the transects.
Data analysis
The population structure of the localities was analyzed considering the proportion of individuals in each size category, using the chi-square (X2) test. An evaluation of the similitude was performed using the analysis of conglomerates (Bray-Curtis distance with average linkage), based on the relative abundance per size categories.
We constructed a Lefkovitch matrix model based on the size categories (Table 3). The matrix was divided in six size categories, including processes of demographic growth (G), permanence (S) and fecundity (F). The growth process (G) represents the probability of individuals passing from one growth category to the next one: Gi = oi * yi, where oi corresponds to the annual survival probability of the category i, and yi is the probability that an individual pass from an i to an i+1 category. The value of yi was calculated as gi/ci, where gi is the annual growth rate measured as the increase in the number of principal veins and ci is the rank of the category i, that is, the number of principal veins of the category. The process of permanence (S) represents the probability of the individuals to survive and stay in the same size category: Si = σi - Gi. The process of fecundity represents the production of seedlings per individual in the adult size categories: Fi = σi * Prob{rep}i * fi, where Prob{rep}i is the probability that an individual in the i category can reproduce and fi is the effective annual reproduction; this means, the reproduction coefficient of recruited seedlings, during the monitored time (Vallejo et al. 2014).
Table 3 Lefkovitch matrix of Attalea nucífera population in municipality of Guaduas, Cundinamarca, Colombia.
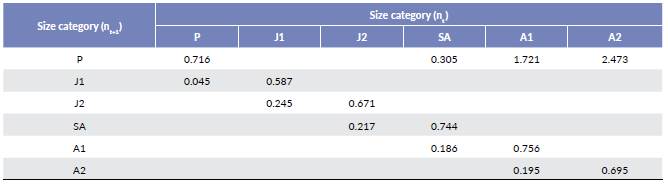
Regression models were used to calculate the transition probabilities for each vital rate (growth, survival, and reproduction) (Table 4). We studied growth through a linear regression model that related initial size (x) to final size (y). Survival and reproduction were studied through logistic regression models that related initial size(x) to survival probability and the probability of having offspring, respectively.
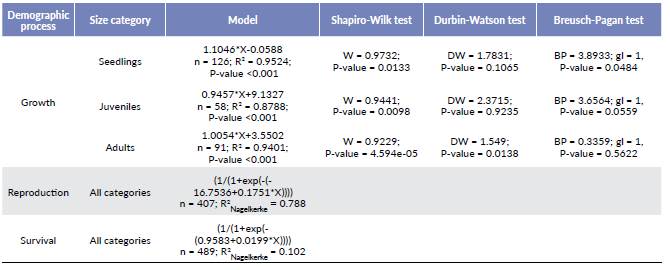
We use the following equation for the matrix model in order to analyze the population dynamics:
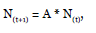
Where, N(t) and N(t+1) represent the population structure in the t and t+1 times, respectively, and A is the square matrix that contains all the transition probabilities among size categories during the study period (Caswell 2001). Thus, when A is multiplied by N(t) repeated times, the relative structure and growth rate of the population become stable. Likewise, when a stable size distribution is reached the dominating eigenvalue (λ), and the right eigenvector of the A matrix are equal to the finite population size, and the stable distribution of states, respectively (Chien et al. 2008). If the finite population growth rate (λ) is larger than one (λ > 1), the population will be growing, while if the rate is lower than one (λ < 1), the population will be decreasing. With the aim to statistically test if the finite population growth rate (λ) is different to one, we calculated the 95 % confidence intervals for the values using the series approximation methods described by Caswell (2001).
The prospective analysis (elasticity) was done to evaluate the relative contribution of different elements of the matrix to the finite population growth rate (λ). To provide information about the potential impacts that could happen in X, by doing small changes in the corresponding matrix (Caswell 2000). Before the elasticity analysis, we evaluated the significative differences in the stable population structure and the population structure observed in the field, using a chi-square (X2) test.
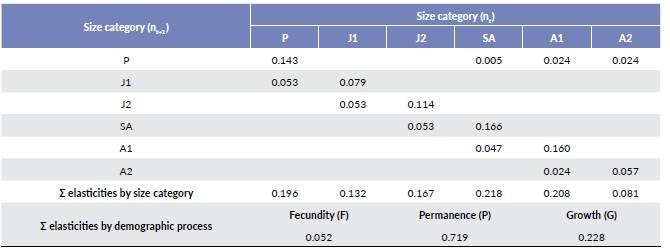
To measure how the population size can vary in the future, under the actual conditions and specific management scenarios, we projected the population structure found in the locality VC for 30 years using a transient analysis of the population dynamic (Groenendijk et al. 2011). We simulated scenarios of conservation and management of A. nucífera, using changes in the inputs of the Lefkovitch matrix, and evaluating their effects in the population size. The simulated effects in each of the scenarios were analyzed using different intensities (20 %, 40 %, 60 % y 80 %), which were related with potential situations, given the current reported conditions for the A. nucífera palm. The first scenario was the establishment of a conservation area, given an increase in the seedlings (P) and early juveniles (J1) survival. The second scenario corresponds to cattle lopping, which resulted in the decrease of the survival of the P and J1 categories (individuals with leaves of easy access for the cattle). The third scenario was the establishment of paddocks in the area, which resulted in a decrease in the survival of the plants in all size categories.
Extinction time
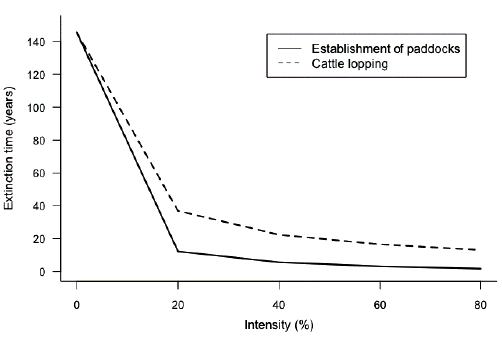
The estimated extinction time (te) was calculated with the methodology proposed by Valverde and Silvertown (1997). In this methodology, it is considered that a species is extinct when the number of individuals reaches 5 % of the original size (extinction threshold) according to the following equation:
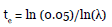
Similarly, we obtained the estimated variation in the times to extinction for the population of A. nucífera in the VC locality, projected for the different intensities of anthropic usage that were modeled. All the statistical analyses were done using the package R version 1.1.383 (R Development Core Team c2009), and for the demographic analysis, the Popbio package (Stubben and Milligan 2007) was used.
RESULTS
Population structure
The localities and populations studied are compared in Table 1 and 5, respectively. The highest total density (ind/ha) was observed in the locality of CS (n = 2143/ ha), followed by the localities of VC (n = 1404/ha), CV (n = 1240/ha), LA (n = 875/ha), GK (n = 520/ha) and CY (n = 450/ha). Although the highest density was observed in CS, this was the locality with the smallest sampling area (0.07 ha). The structure of the populations was found to vary significantly among localities (X2 = 1817; gl = 25; P-value < 0.05). In general, most individuals are concentrated in the seedling size category (P), while the lowest proportion of individuals is distributed among juveniles (J1 and J2) and adults (SA, A1 and A2); except in the CY locality where no seedlings (P) and early juveniles (J1) were reported. Also, in the locality CV, the pattern is different because a higher density of sub-adults (SA) is observed (Table 5, Fig. 1).
Table 5 Population density (ind./ha) of Attalea nucifera in six localities of the Magdalena River valley, Colombia.
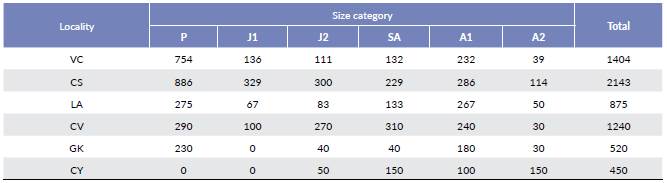
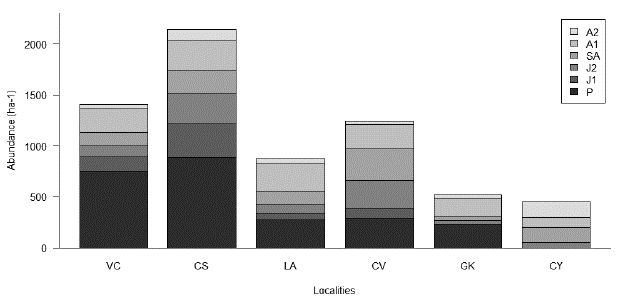
Figure 1 Population structure of Attalea nucífera in the Magdalena River valley, Colombia. See abbreviations in Table 2.
According to the conglomerate study of the plant density by size categories, we observed three different population groups, with similarities between the localities VC and CS, and among GK, LA and CV (Fig. 2). The first group (VC-CS) corresponds to localities where cattle raising and small-scale agriculture are in practice, using the Guaduero river bank (municipality of Guaduas). For the second group (GK, LA, CV), only the localities of LA and CV share the characteristic of having an environmental protection. The third group (CY) is not related to any particular characteristic and shows the most significant differences with the other localities.
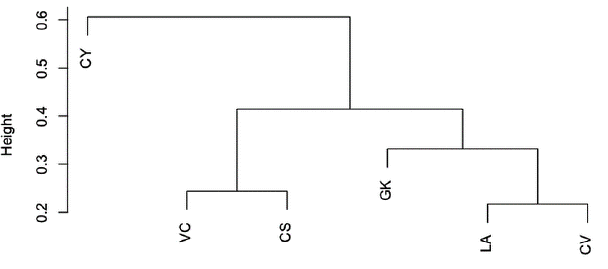
Figure 2 Cluster dendrogram illustrating the similarities among Attalea nucifera populations in six localities of the Magdalena River valley, Colombia. See abbreviations in Table 1.
Population dynamics
The population in the locality CV (municipality of Guaduas) is decreasing in size (λ = 0.979; CI95 % = 0.962-0.997). Likewise, the stable population structure obtained presents no significant differences with the population structure observed in the field (X2 = 30; gl = 25; P-value > 0.05). The elasticity analysis shows that the demographic process that contributes the most to λ is the permanence (71.9 %), followed by growth (22.8 %) and fecundity (5.2 %) (Table 6). Equally, the elasticity matrix shows that the highest value corresponds to the SA size category, followed by Al, P, J2, J1 and A2 (21.8 %, 20.8 %, 19.6 %, 16.7 %, 13.2 % and 0.81 %, respectively).
Transient dynamic and scenario simulations
The transient dynamic projected to 30 years, under the current conditions observed in the VC locality, shows that the population size slightly increase until year six. Then, the population decreases until reaching a lower abundance than the initial one. Under the scenario where a conservation area is established, the population size increases significantly, mainly after 40 % of seedling (P) and early juveniles (J1) survival have been positively affected (Fig. 3a). For the scenario where paddocks and cattle lopping are used, the population size decreases drastically (Fig. 3b and 3c).
Extinction time
Under the conditions studied in the locality of VC, the population will reach the extinction threshold in 145 years. However, this situation can become more critical in scenarios where there is an intensive anthropic effect, such as the establishment of paddock areas and by cattle lopping. For the paddock and cattle lopping scenarios, the extinction is achieved in less than 40 years, once the 20 % of the intensity of the modeled effect has been reached (Fig. 4).
DISCUSSION
The population structure results show that the density distribution per size categories can be a consequence of different anthropic pressures, as determined by different land use categories and the main transformation factor, similarly as it has been observed for other species of plants (Helm and Witkowski 2012, Cousins et al. 2014) and palms (Lozada and Moraes 2013, Melito et al. 2014). Statistically, the structure of the A. nucifera populations is different among all localities. However, the conglomerates' analysis of the size category densities shows that the localities of VC and CS of the Guaduas municipality, have the most similar population structures, given that both are located in areas with similar anthropic land use (cattle raising and small-scale agriculture). Studies of the population structure in palms have shown that the presence of cattle in nearby areas, directly affects the density of plants in the different size categories. This effect can be positive, such as seed dispersion and old padlocks colonization (Uribe et al. 2001, Lozada and Moraes 2013), or negative such as an increase in mortality (Lopez-Toledo et al. 2011).
Another population group was identified across the localities of GK, LA and CV. This group covers localities that seem geographically different, but have some characteristics in common. LA and CV are located in environmentally protected and ecotourism controlled areas, where populations are recovering. GK is not a protected area, but palms grow densely inside of small forest relicts surrounded by cattle raising and African oil palm crops. We, thus, infer that the historic land use and environmental characteristics can affect the structure of the A. nucifera populations. Likewise, the level of perturbation or conservation of the area can affect the population structure. Lopez-Toledo et al. (2011) and Melito et al. (2014) stated that the level of environmental protection of an area could produce structural differences in the Brahea aculeata (Brandegee) H.E. Moore and Euterpe edulis Mart. populations. We think this can explain the similitude found between the populations of the LA and CV localities, despite their spatial differences. On the other hand, the population sampled in the locality of CY exhibits a structure that is different from the other ones. This population has a small number of individuals restricted to a forest relict located within a paddock area.
Generally, almost all studied populations, except the ones in the CV and CY localities, have a higher number of seedlings, while the lower numbers are for juveniles and adults. The localities of CS, VC, LA and GK probably have self-regenerative populations with an inverted J-shape form (Thompson et al. 2009). These populations are characterized by having individuals of almost all growth stages, where death individuals are replaced by younger ones (Galeano et al. 2010). Given that for a significant number of palms the typical distribution of sizes has an inverted J-shape form, it is considered that the populations in the CV and CY localities have conditions that are atypical and particular of each locality. The CV locality has a population with regeneration characteristics, which is located in the Cabildo Verde Natural Reserve, an environmentally protected private area since 1994 (ONG Cabildo Verde pers. com.). This protected area is surrounded by ample extensions of African oil palms crops with extensive cattle raising activities. The population in the locality of CY is a small relict of forest that has been profoundly affected by cattle activities in the surrounding areas. Negative (disturbances) and positives (natural reserves) anthropic modifications in the surrounding landscape, and the landscape context, as described in Idohou et al. (2016), can have the ability to affect the structure and the distribution of the Hyphaene thebaica (L.) Mart. palm in its natural habitat. We believe that the landscape context is determining a similarly low proportion of seedlings in populations that have had a different history of use.
The incidence of the surroundings over the study areas, allow us to infer, that cattle, African oil palm, small-scale agriculture, among other factors, are affecting in different ways the distribution of the size categories of the populations studied. Extensive cattle raising and African oil palm plantations require the complete removal of the original vegetation. In contrast, low scale activities cause only local and small-scale disturbances, which can coexist with the original vegetation. This coexistence allows that the cattle enter forest patches inhabited by A. nucífera affecting only individuals that are of easy access or from the size categories that are most palatable. The susceptibility of seedlings to cattle lopping when they are located in natural habitats near paddocks has been described by Mandle and Ticktin (2012), Andrade-Erazo and Galeano (2016), and García et al. (2017) in other palm species.
The results from the matrix model show that the population of A. nucifera in the locality of VC (Guaduas municipality) has an annual negative population growth rate of 2.1 %. According to the elasticity analysis, if the conservation goal is to increase the finite population growth it is necessary to design management actions that guarantee survival of seedlings, sub-adults and young adults palms. According to our simulation results, establishing a conservation area that guarantees an increase in the survival of the most susceptible size categories to cattle lopping (P and J1) by 20 % produce a significant population growth (Fig. 3a). Thus, the most effective conservation plan for A. nucifera is the increase in the number or extension of protected areas where it is distributed, as currently only two protected areas are reported (Prada-Ríos and García 2019). Such measurements are needed to guarantee the long-term survival of the palm, since our results show that under paddock land use with cattle lopping scenarios populations decrease in size to the point of potentially becoming extinct in less than 40 years. The conversion of natural vegetation to active pastures produces significant effects on the demography of the plants, inhibiting the establishment of seedlings and therefore regeneration (Morales-Romero et al. 2012). Although some authors report that cattle browsing does not significantly affects the finite rate of population growth (Sánchez-Velásquez and Pineda-López 2010, Aschero et al. 2016), Nazareno and dos Reis (2014) indicate that populations of palm Butia eriospatha (Mart. ex Drude) Becc. are severely affected in its structure because of such action.
Palm species that grows in anthropically transformed environments show a different demographic response to the surroundings. The palm species Astrocaryum malybo H.Karst., A. chambira Burret and A. standleyanum, which grow in disturbed surroundings exhibit finite population growth rates (X) over one (1), (García et al. 2016, 2017) while the palms Sabal yapa C.Wright ex Becc. and Desmoncus orthacanthos Mart., located in the forest fragments within an area dedicated to paddock and agriculture, show X values of 0.959 (Pulido et al. 2007) and 0.964 (Escalante et al. 2004), respectively. A study using three palm species found that, under similar conditions, Astrocaryum aculeatissimum (Schott) Burret and Geonoma schottiana Mart. had a higher tolerance to anthropic disturbances, while Euterpe edulis Mart. had a negative growth rate (Portela et al. 2010). In Euterpe precatoria Mart. a positive growth rate of 3 % has been observed in secondary forest, while in a primary forest the population is reduced to a growth rate of 1.5 % (Fernández Otárola and Avalos 2014).
Although the short period of observations of the CV population (one year) might affect our estimations of vital rates, we think that it does not affect our conclusions. Other palm species under similar environmental conditions have shown not significantly different in vital rates between consecutive years (García et al. 2016). Additionally, population studies of Chamaedorea radicalis Mart. have found that vital rates may have remained constant in long-term studies (Endress et al. 2004, 2006).
Population decrease observed in VC locality (Guaduas) suggests that Attalea nucifera could be less tolerant of environmental disturbances caused by the expansion of agricultural frontier. However, it is premature to generalize this trend, since results of population structure show the persistence of this species in other very disturbing locations in the Middle Magdalena Basin. In this sense, it is necessary to improve our understanding of its population dynamics, as well as seed germination and seedling establishment in different disturbance conditions, to evaluate its natural regeneration and propose conservation strategies to restore its populations.

