











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink
INTRODUCCIÓN
La dispersión de semillas es considerada un mecanismo reproductivo fundamental en los ecosistemas y son los vertebrados la clave para que esto se pueda dar en algunas especies vegetales (Howe y Smallwood 1982, Wang y Smith 2002). La frugivoría es un hábito alimentario que ha aparecido a lo largo de la evolución en muchos vertebrados (Howe y Smallwood 1982, Fleming y Kress 2011), quienes en el trópico dispersan más del 80 % de las especies leñosas (Howe y Smallwood 1982), dentro de este grupo las aves y los murciélagos frugívoros son los agentes dispersores de mayor importancia, en especial en las zonas perturbadas o fragmentadas donde se da la dispersión principalmente de especies vegetales pioneras (Medellín y Gaona 1999, Galindo-González et al. 2000).
En Colombia hay aproximadamente 209 especies de murciélagos, 123 pertenecen a la familia Phyllostomidae (Sociedad Colombiana de Mastozoología c2017), siendo los géneros más representativos: Artibeus, Carollia y Sturnira (Medellín y Gaona 1999, Saldaña-Vázquez 2014, Arias y Pacheco 2019). La especialización en la dieta de los murciélagos depende de factores extrínsecos como características físicas de la fruta, contenido nutricional y su disposición en el follaje, así como de factores intrínsecos como la fisiología del murciélago (Muscarella y Fleming 2007). Estos factores son relevantes en la conformación de ensamblajes de murciélagos (Saldaña-Vázquez 2014).
Se ha evaluado en diferentes estudios que la dispersión de semillas es el principal beneficio que los murciélagos proporcionan a las plantas (Traveset 1998, Traveset y Verdú 2002) considerándolos así efectivos dispersores de un gran número de especies vegetales, consumiendo frutos de alrededor de 549 especies (Lobova et al. 2009), siendo Vismia, Cecropia, Piper, Solanum y Ficus los géneros más consumidos (Charles-Dominique 1986, Galindo-González 1998, Muscarella y Fleming 2007). Debido a su capacidad para defecar mientras vuelan, generan una lluvia de semillas (Charles-Dominique 1986) a lo largo de grandes distancias recorridas que les permiten incluso llegar a zonas perturbadas, siendo así importantes para la restauración de los bosques (Fleming y Heithaus 1981, Wunderle Jr 1997, Galindo-González et al. 2000).
Otro de los servicios evaluados es la mayor probabilidad de germinación de la semilla al pasar por el tracto digestivo del murciélago, y aunque en algunos estudios respaldan esta idea (Traveset y Verdú 2002), trabajos recientes indican que las semillas consumidas por los murciélagos no incrementan su germinación y que la probabilidad de esta varía entre las especies de plantas dependiendo, además, de la especie de murciélago que la consume (Saldaña-Vázquez et al. 2019).
Los estudios que se han realizado sobre la dispersión o germinación de semillas por murciélagos se han enfocado principalmente en los bosques húmedos tropicales y en ecosistemas no boscosos (Galindo-González et al. 2000, Estrada-Villegas y Pérez-Torres 2010, Heer et al. 2010, Arias y Pacheco 2019, Daza-Gallo et al. 2019) y son pocas las investigaciones al respecto en el bosque seco tropical (Bs-T) (Novoa et al. 2011, Ríos-Blanco y Pérez-Torres 2015, García-Herrera et al. 2019), a pesar de ser un ecosistema altamente amenazado en Colombia y en el mundo (Pizano y García 2014).
El propósito de este trabajo fue registrar cuáles son los frutos que están siendo consumidos por el ensamble de murciélagos frugívoros y cuál es su efecto sobre la germinación de dos especies de semillas en el Área Natural Única Los Estoraques, Norte de Santander, Colombia.
MATERIALES Y MÉTODOS
Área de estudio
El trabajo se realizó en el Área Natural Única Los Estoraques (ANULE) perteneciente al Sistema de Parques Nacionales Naturales de Colombia; se encuentra ubicada en la Cordillera Oriental de los Andes en el departamento de Norte de Santander. El ANULE tiene una extensión de 640,2 ha, se caracteriza por sus severos procesos de erosión debido a la acción de la escorrentía, precipitación y acción del viento que ha generado numerosas geoformaciones que pueden llegar hasta los 30 m de altura (Aparicio y Rojas 2003), pertenece a una zona de vida de bosque seco premontano (bs-PM) (Holdridge 1947, Espinal y Montenegro 1977). La vegetación del ANULE se considera subxerofítica, clasificada como vegetación semiárida subandina, siendo un tipo de bioma azonal donde hay zonas con déficit de humedad, con una vegetación discontinua y en su mayoría de tipo arbustivo y subarbustiva de 5-7 m, encontrando especies de las familias Myrtaceae, Clusia-ceae, Hypericaceae y Cactaceae (Suárez et al. 2016).
Captura de murciélagos
La captura de murciélagos se realizó durante cinco sesiones de campo en 2019: 15-27 abr, 17-29 jun, 8-18 ago, 11-21 sep y 10-13 dic, abarcando tanto la época seca como la época lluviosa. Se instalaron cuatro redes de niebla de 2,5x12 m por 24 noches en las diferentes coberturas del ANULE: vegetación asociada a las geoformaciones, arbustales densos en poca pendiente, bosque de galería y piedemonte de bosque subandino. Las redes se mantuvieron abiertas durante cuatro horas y media (19:00-23:30 h) y fueron revisadas cada 25 min para extraer los murciélagos (Novoa et al. 2001, Ríos-Blanco y Pérez-Torres 2015); el esfuerzo de captura fue de 432 redes/hora/noche. Las especies de murciélagos se identificaron siguiendo a Díaz et al. (2016) y se verificó su distribución geográfica siguiendo a Solari et al. (2013). A cada uno de los individuos se les tomaron medidas morfométricas generales y datos de sexo, peso, estado de desarrollo; también se realizó registro fotográfico (Kunz y Parsons 2009).
Recolección e identificación de semillas
Luego de ser capturados los murciélagos se mantuvieron en bolsas de papel, al interior de bolsas de tela durante 25 min para obtener las muestras fecales, éstas fueron almacenadas en tubos de microcentrífuga y fueron etiquetadas con el número de captura del murciélago, posteriormente las semillas se limpiaron con agua y se fijaron en FAA (for-maldehído, alcohol, acético), el cual se usa comúnmente para fijar tejidos vegetales. Para la identificación de las semillas se utilizaron artículos y guías especializadas en identificación de semillas las cuales tienen en cuenta las características morfológicas como el color, tamaño, forma, textura, presencia de cicatrices y quillas, entre otros (Ríos R et al. 2004, de Oliveira y Pereira 2016, Daza-Gallo et al. 2019), también se usaron las muestras de referencia tomadas del área, permitiéndonos llegar a la identificación a nivel de especie. Las semillas y material de referencia fueron depositados en la Colección Biológica de la Universidad CES (CBUCES), con los números CBUCES-J 100 al 309, CBUCES-J 415 al 424 y CBUCES-H 484 al 503 (Anexo).
Germinación de semillas
Para evaluar el efecto de la germinación sobre las semillas obtenidas de las excretas se escogieron dos especies vegetales, con base en su abundancia en las excretas y en los frutos tomados de las muestras de referencia garantizando un mayor número de réplicas; estas fueron Vismia baccifera (L.) Planch. & Triana y Piper sp4; esta última se escogió además por su disponibilidad para el manejo. Se realizaron dos tratamientos para el ensayo de germinación: las semillas obtenidas de las excretas de los murciélagos, de las cuales no se diferenció la especie de murciélago fuente para obtener la mayor cantidad de muestras, y las semillas obtenidas de frutos directamente. Para acercarse lo más posible a las condiciones naturales como temperatura y humedad, los ensayos de germinación se realizaron en el invernadero del ANULE.
Para Vismia baccifera se sembraron 20 semillas por cada caja petri, con un total de diez cajas por cada tratamiento y para Piper sp4 se sembraron 20 semillas por cada caja de petri, con un total de seis cajas por cada tratamiento.
Las semillas se sembraron con base en el protocolo presentado por Ríos-Blanco (2010). Se colocaron en cajas de Petri con papel absorbente, se rotularon y fueron cubiertas por una malla que permitiera el paso del agua y evitará la pérdida de las semillas; para el suministro de agua se emplearon los dispersores automáticos del invernadero del área programados para regarlas dos veces al día. Se revisaron semanalmente, se contó el tiempo y número de semillas germinadas y se definió que una semilla había germinado una vez se podía identificar la radícula. El conteo se realizó semanalmente hasta que no se presentaron cambios en el número de semillas germinadas en el lapso de una semana, según lo propuesto por Ríos-Blanco (2010).
Análisis estadísticos
Mediante el programa Estimates 9.1.0 (Colwell c2013) se construyó una curva de acumulación para evaluar la riqueza de especies de semillas que fueron consumidas por los murciélagos, ésta se comparó con los estimadores Chao1: especies esperadas basado en el número de especies raras, Jack 1: basado en la presencia-ausencia, y Bootstrap: riqueza estimada basada en la proporción de cada especie (Villarreal et al. 2006). Se calculó el índice de mejor dispersor DII para evaluar la importancia de cada especie de murciélago como potencial dispersor, el cual se basa en la abundancia relativa de los murciélagos capturados y el porcentaje de semillas obtenidas por cada uno, indicando valores del 0-10 siendo 0 la especie que no dispersa ninguna semilla y 10 que dispersa todas las semillas registradas (Galindo-González et al. 2000). Se calculó el índice de Mo-risita para caracterizar el solapamiento de dieta por pares de especies de murciélagos, como una medida de cómo se reparten las semillas disponibles en el bosque, evaluando la proporción de las semillas obtenidas entre las dos especies, este tiene valores de 0-1, siendo 0 un solapamiento nulo y 1 solapamiento completo (Krebs 1999), para este índice solo se analizaron las especies de murciélagos que tenían más de cinco registros de semillas.
El éxito de germinación de las semillas se evaluó mediante la comparación del índice de capacidad de germinación (ICG), el cual indica el porcentaje de semillas que germinaron al finalizar el experimento (González-Zertuche y Orozco-Segovia 1996, Segura-Linares 2017). Para evaluar si había diferencias en el índice de germinación entre los dos tratamientos se utilizó la prueba Mann-Whitney-Wilcoxon (Bauer 1972) usando el software R versión 3.0.2 (R Core Team c2018), con una confiabilidad del 0,90.
RESULTADOS
Se capturaron 290 murciélagos frugívoros pertenecientes a nueve especies de la familia Phyllostomidae. Las especies más abundantes fueron Carollia brevicauda (Schinz, 1821) (64,5 %), Dermanura glauca (Thomas, 1893) (16,6 %) y Enchisthenes hartii (Thomas, 1892) (4,8 %) (Tabla 1).
Se obtuvieron 138 excretas con semillas provenientes de las nueve especies de murciélagos, 26 de estas contenían semillas de dos, tres o cuatro especies vegetales, por lo cual el número de registros de semillas total fue de 164. Se identificaron 18 morfoespecies de semillas (Fig. 1) siendo Vismia baccifera, Piper sp3, Piper sp1 y Calycolpus moritzianus (O. Berg) Burret las más frecuentes con una representatividad del 25 %, 15,2 %, 14,6 % y 14 % respectivamente (Tabla 1). De acuerdo con la curva de acumulación se encontraron 18 especies de semillas que cuando se compararon con los indicadores de riqueza se obtuvo una representatividad del muestreo para Chao1, Jack 1 y Bootstrap del 90,0 %, 74,20 % y 86,30 % respectivamente (Fig. 2).

Figura 1 Morfoespecies de semillas obtenidas de las excretas de murciélagos frugívoros en el Área Natural Única Los Estoraques (ANULE), a. Cecropia sp, b. Calycolpus moritz ianus, c. Ficus spi, d. Ficus sp2, e. Solanaceae spi, f. Solanaceae sp2, g. Pi per spi, h. P iper sp2, i. Piper sp3, j. P iper crass inervium, k. Piper sp4, l. Piper sp5, m. Piper sp6, n. Vismia baccifera, o. Xanthosoma spi, p. Psammisia penduliflora, q. Cavendishia pubescens, y r. Syzygium jambos. Escala = imm
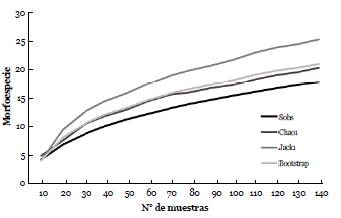
Figura 2 Curvas de acumulación y de estimación de las morfoespecies de semillas encontradas en las excretas de murciélagos para el Área Natural Única Los Estoraques (ANULE). Sobs: especies observadas, Chaol, Jack 1 y Bootstrap.
Tabla 1 Cantidad de individuos capturados por especie de murciélagos frugívoros, índice de mejor dispersor (Dll) y frecuencia de morfoespecies de semillas consumidas por especie.
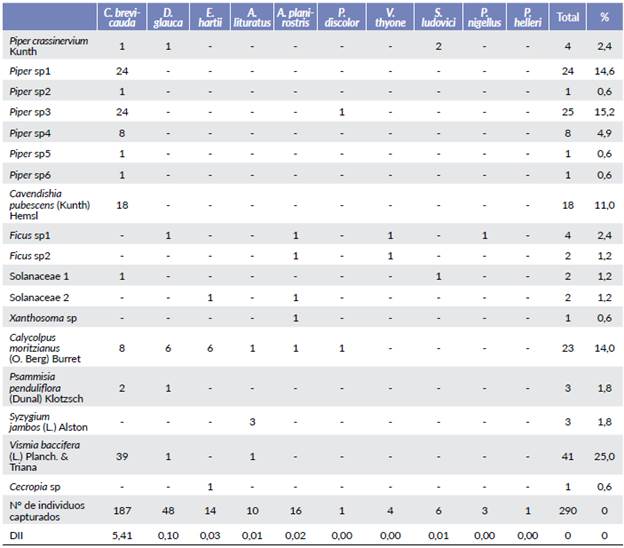
La dieta de Carollia brevicauda fue la más amplia con once morfoespecies (128 muestras) siendo Vismia baccifera la más frecuente (39), seguida de Piper sp1 (24) y Piper sp3 (24). Dermanura glauca tuvo la segunda dieta más amplia con cuatro morfoespecies (diez muestras) predominando la especie Calycolpus moritzianus (diez) (Tabla 1).
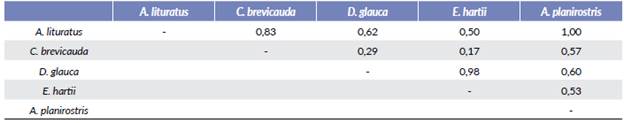
Carollia brevicauda fue el dispersor de mayor importancia (DII=5,41) seguido por Dermanura glauca (DII=0,10) (Tabla 1). Según el índice de Morisita, el mayor solapa-miento lo presentaron Enchisthenes hartii y Dermanura glauca (0,98) seguidas por Carollia brevicauda y Artibeus lituratus (Olfers, 1818) (0,83) (Tabla 2).
Tabla 2 Solapamiento de dieta calculado mediante el índice de Morisita para los murciélagos frugívoros del Área Natural Única Los Estoraques (ANULE). En negrita los valores más altos.
Experimento de germinación
Las semillas de V. baccifera empezaron a germinar a la tercera semana y terminaron hacia la sexta semana. El porcentaje de germinación de las semillas sembradas de las excretas fue del 73,5 % y de las sembradas del fruto fue de 86,0 %, sin embargo, esto no representó una diferencia significativa (U=0,37, P = 0,22). Las semillas de Piper sp4 empezaron a germinar a la cuarta semana y terminaron de germinar en la sexta semana. El porcentaje de germinación de las semillas sembradas de las excretas fue del 14,5 % y de las sembradas del fruto fue de 4,0 %, pero no se presentaron diferencias significativas (U=0,41, P = 0,18).
DISCUSIÓN
Para el ANULE se conocían únicamente seis especies de murciélagos frugívoros antes de la realización de este trabajo, en el cual se registraron adicionalmente las especies D. glauca, V. thyone y S. ludovici. Nuestros resultados mostraron que estas especies adicionales participan activamente en el consumo de diferentes especies vegetales en el área protegida, especialmente D. glauca que fue una especie frecuente en los muestreos y consumió frutos no reportados anteriormente en la literatura como C. moritzianus (Castaño et al. 2018).
El ecosistema seco presente en el ANULE es un tipo de bioma azonal (Suárez et al. 2016), condición que pudo favorecer el registro de nuevas interacciones planta-animal, por ejemplo, hasta ahora no se había registrado en la literatura el consumo de la especie C. moritzianus por parte de los murciélagos, a pesar de que esta planta tiene una distribución amplia en los Andes tropicales (Landrum 2010, Yá-ñez Rueda et al. 2016). Los frutos de esta especie fueron consumidos por seis especies de murciélagos en al área de estudio y fue común en la dieta de tres de ellos (Tabla 1). Si bien se han reportado el consumo de al menos seis especies de plantas de la familia Myrtaceae, esta no suele ser de las familias más frecuentemente consumidas por los murciélagos, los cuales tienen mayor asociación con las familias Piperaceae, Urticaceae, Moraceae, Solanaceae y Clusiaceae (Castaño et al. 2018, Regolin et al. 2020).
La riqueza de las especies vegetales consumidas por murciélagos que se obtuvo en el ANULE fue mayor a la que se ha registrado en otros estudios realizados en el bosque seco, como los encontrados por Segura-Linares (2017) de ocho morfoespecies vegetales, García-Herrera et al. (2019) de once morfoespecies y Ríos-Blanco y Pérez-Torres (2015) de quince morfoespecies vegetales consumidas. Aunque los frutos más frecuentes en las dietas encontrados en todas las localidades fueron V. baccifera y Piper spp, para el caso de ANULE, C. moritzianus fue una especie altamente consumida, a la par de algunas especies del género Piper, lo cual sugiere que este es un recurso importante localmente, al menos temporalmente, para el ensamblaje de especies frugívoras.
C. brevicauda fue la especie más abundante y la que presentó la dieta más amplia con doce morfoespecies consumidas; las especies de los géneros Piper y Vismia fueron las que presentaron mayor frecuencia de consumo lo cual ha sido previamente documentado por otros autores. Gorchov et al. (1995), Zavala-Calloapaza (2018) y Arias y Pacheco (2019) concluyen que Carollia tiene una dieta compuesta principalmente por Vismia, mientras que Novoa et al. (2011), Lobova et al. (2009) y Sánchez et al. (2012) registraron que Carollia tiene un mayor consumo de especies del género Piper. No obstante, se debe tener en cuenta que esto puede variar en diferentes épocas del año, por ejemplo, en los últimos muestreos de este trabajo, entre los meses septiembre y diciembre, V. baccifera fue la especie que encontramos con mayor fructificación en el área y de la que obtuvimos semillas en las excretas con mayor frecuencia.
El solapamiento en la dieta entre las especies de murciélagos se da principalmente cuando estos tienen tamaños y hábitos similares (Sánchez et al. 2012, Montoya-Bustamante et al. 2016, Zavala-Calloapaza 2018) como se observó entre D. glauca y E. hartii, los cuales presentaron el mayor índice de Morisita (Tabla 2). Sin embargo, aunque A. lituratus es una especie grande (AB> 55 mm) y con hábitos de forrajeo de dosel (Soriano 2000), y C. brevicauda es una especie de tamaño mediano (AB= 36-43 mm) y de estratos a nivel del sotobosque (Soriano 2000), el solapamiento que presentaron fue también alto (Tabla 2). Este resultado puede deberse a la baja estratificación del bosque en el área de estudio ocasionando que A. litura-tus consuma lo que hay disponible en los estratos medios como V. baccifera y C. moritzianus, sobreponiendo su dieta con especies más pequeñas. Así mismo en áreas intervenidas y fragmentadas, el solapamiento de nicho entre las diferentes especies de murciélagos puede aumentar debido a la baja disponibilidad de frutos de algunas especies vegetales. Si bien el ANULE es un área protegida, es un área relativamente pequeña en medio de una paisaje históricamente intervenido y fragmentado, donde la red de interacción entre murciélagos y plantas puede variar en relación con los sitios altamente conservados (Regolin et al. 2020), presentando nuevas interacciones o variaciones en las dietas esperadas.
Al ser C. brevicauda el mejor dispersor, debido a la magnitud con la cual transporta semillas de un sitio a otro facilitando su disposición en sitios donde se pueden establecer, la especie es clave en el área de estudio y podría tener un papel central en los procesos de restauración mediante la utilización de refugios o cebaderos artificiales, aumentando la lluvia de semillas en estos lugares (Galindo-González 1998, Kelm et al. 2008, Preciado-Benítez et al. 2015). Ya que ha sido bien documentado en diferentes trabajos que la dieta de los murciélagos se ve representada en mayor proporción por especies pioneras como lo son las especies de los géneros Piper y Vismia (Oria y Machado 2007, Saldaña-Vázquez 2014, Zavala-Calloapaza 2018), esta especie debería ser considerada clave para los procesos de enriquecimiento en el área, en especial en las zonas que han sido perturbadas por factores como ganadería y agricultura (Suárez et al. 2016).
Experimento de germinación
Son escasos los trabajos sobre germinación de las especies del género Vismia, a pesar de que son muy frecuentes en la dieta de los murciélagos frugívoros (Arteaga 2007). Si bien esta fue una primera aproximación para conocer el efecto del consumo por parte de los murciélagos sobre su germinación, nuestros resultados muestran que no hay un efecto del paso de las semillas por el tracto digestivo de los murciélagos. Esto mismo ha sido encontrado por diferentes trabajos (Saldaña-Vázquez et al. 2019), por lo tanto, el beneficio de la interacción entre murciélagos y las especies del género Vismia, sería en la fase de trasporte y disposición de las semillas hacia sitios donde estas se pueden establecer, incluso a varios kilómetros de distancia (Galindo-Gonzales 1998, Regolin et al. 2020). No obstante, en nuestro experimento, tuvimos un número bajo de réplicas, no diferenciamos las especies de murciélagos de donde provenían las semillas y no tuvimos en cuenta aspectos como el tamaño de la semilla, su posición en el fruto, el grado de madurez de la semilla y la época del año, variables que pueden influir sobre la germinación de estas (Gray y Thomas 1982, Arteaga 2007). Por lo cual, consideramos que nuestros resultados deben ser interpretados con cuidado.
En el caso de Piper sp 4, a pesar de que no encontramos diferencias estadísticas en la germinación de las semillas, si se observó una tendencia a favorecer la proporción de semillas germinadas obtenidas de las excretas puesto que la diferencia neta fue alta (14,5 % vs 4 %), pero con una amplia variación en los resultados de cada réplica. Por lo cual sugerimos utilizar un mayor número de réplicas en estudios posteriores. No obstante, en investigaciones realizadas recientemente, no se ha encontrado evidencia de que la germinación de las semillas del género Piper se vea favorecida al pasar por el tracto digestivo de los murciélagos frugívoros, lo cual sugeriría que el beneficio para las plantas esté en la dispersión y no en la germinación (Saldaña-Vázquez et al. 2019, Torres et al. 2019). Como en el caso del experimento con Vismia, no tuvimos en cuenta las variables adicionales que pueden afectar la germinación de las semillas y obtuvimos menor número de réplicas, por lo que consideremos que para este caso los resultados no deben ser interpretados como concluyentes.
Este trabajo resalta el desconocimiento que aún se tiene de la diversidad al interior de las áreas protegidas en el país. En este trabajo se aumentó considerablemente el conocimiento de la riqueza de especies de murciélagos del área (siete especies en total, tres de ellas frugívoras) y se encontraron 18 especies de frutos consumidas, lo cual corresponde a la riqueza más alta hasta ahora documentada para los ecosistemas secos en el país. El ensamblaje local de especies de murciélagos frugívoros comparte mucho los ítems alimenticios lo cual puede ser una respuesta a la estructura del bosque y al grado de transformación histórica en la región.
C. brevicauda si bien es una especie común a lo largo de todos los bosques tropicales, localmente es una especie clave en el proceso de dispersión de la mayoría de las especies vegetales consumidas por murciélagos, no solamente por su amplia dieta si no por su abundancia en el área que se traduce en una mayor lluvia de semillas generada. Los murciélagos parecen no tener un efecto sobre la germinación de las semillas del género Vismia ni del género Piper, pero recomendamos replicar los experimentos con un mayor número de semillas y controlando variables adicionales como la especie del murciélago, el grado de madurez del fruto y el tamaño de la semilla.

