











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El halomorfismo genera alteraciones en las propiedades físicas y químicas del suelo y es una de las causas más importantes de degradación y contaminación ambiental en regiones áridas y semiáridas (Losinno et al. 2005, Lamz-P y González-C 2013, Prause et al. 2014). Este problema da origen, principalmente, a dos tipos de suelos: los salinos y los sódicos. En los suelos salinos predomina una elevada concentración de sales solubles, siendo los cloruros, sulfatos y carbonatos de metales alcalinos y alcalinotérreos los principales compuestos químicos presentes, y en los sódicos, prevalece el catión sodio en la solución micelar (Breckle 1990, Cantero et al. 1996, Obregón-S 1996, Cobo-M 2001a, Lamz-P y González-C 2013, Prause et al. 2014). Debido a un manejo inadecuado del suelo y del agua de riego, la alta concentración de sales en regiones con tierras de uso agrícola genera estrés en los cultivos, limita la absorción del agua, disminuye el crecimiento y desarrollo de las plantas y merman los rendimientos y la calidad de los productos obtenidos (Lamz-P y González-C 2013, Sánchez et al. 2016, Aruani et al. 2017), lo que conduce en algunos casos al abandono de los cultivos (Alonso-S et al. 2016).
En este sentido, Maas y Hoffman (1977) estudiaron la relación entre la salinidad del suelo y los rendimientos relativos de los cultivos. Por ejemplo, en Vitis spp. hallaron que el rendimiento de uvas comienza a decrecer cuando el suelo excede un valor de conductividad de 1,5 mmho/ cm y que, a partir de este valor y por cada unidad de conductividad eléctrica que se eleva, el rendimiento relativo disminuye 9,6 % aproximadamente.
Según Puchulu (2008), Argentina es el tercer país a nivel mundial con mayor extensión edáfica perjudicada por halomorfismo. De un total de 584 049 ha afectadas por salinidad, el 7,9 % corresponde a la provincia de Río Negro (Sánchez et al. 2016).
En el Alto Valle de Río Negro, ubicado en el norte de la Patagonia argentina, la infraestructura hidráulica generó un área productiva bajo riego que permitió el desarrollo de cultivos frutihortícolas, forrajeros (Apcarian et al. 2014) y forestales (Thomas 2015). La excesiva aplicación de láminas de riego y el drenaje deficiente ocasionan el ascenso de la capa freática con sales disueltas que provocan su acumulación progresiva en el perfil del suelo y un aumento en el valor de pH y R.A.S. (relación de adsorción de sodio), además de modificar el entorno radical de las plantas y propiedades del mismo (Sánchez y Curetti 2006). Consecuentemente, los suelos se degradan y se limita su uso para el cultivo de distintas especies (Sánchez y Villareal c2012).
La concentración y tipo de sales, junto a la textura, son parámetros edáficos que promueven el halomorfismo de los suelos (Obregón-S et al. 1996, Sánchez-B y Guevara-F 2013). Como consecuencia, prosperan halófitos, plantas capaces de sobrevivir en suelos fisiológicamente secos, para lo cual han desarrollado distintos mecanismos de adaptación como suculencia de los órganos, acumulación de sales en las vacuolas y desarrollo de glándulas almacenadoras de sales, entre otros (Breckle 1976, Braun-B 1979, Flowers et al. 1986, Font-Q 2001).
Si bien existen estudios florísticos vinculados con las actividades productivas de la región del Alto Valle de Río Negro (Conticello y Gandullo 1991, 1992, Cerazo et al. 2000, Conticello y Bustamante 2001), resulta de interés investigar la flora halófila presente en suelos halomórficos de distintos grupos texturales y bajo diversos cultivos. El conocimiento y el estudio de la flora halófila en sistemas cultivados ayuda a interpretar la calidad edáfica para una posterior intervención mediante labores de mejora. El objetivo del presente trabajo es analizar la diversidad de taxones halófilos en diferentes cultivos y grupos texturales de los suelos halomórficos en el área del Alto Valle de Río Negro.
MATERIALES Y MÉTODOS
Área de estudio
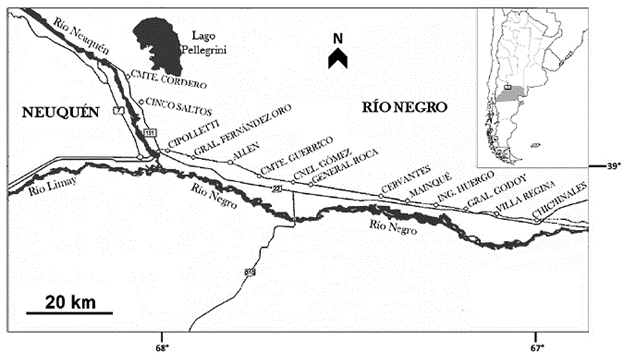
El presente estudio se llevó a cabo en distintos establecimientos productivos del Alto Valle de Río Negro (Argentina), ubicado entre los 66°45' Este y 68°30' Este y los 38°45' Sur y 39°10' Sur, con una extensión de 652 km2 (Fig. 1). El relieve es plano a muy suavemente ondulado, presenta una pendiente general Oeste-Este de aproximadamente 0,8 % (Apcarian et al. 2014). Los suelos son de naturaleza aluvial, con pertenencia a distintos subgrupos y familias de los órdenes Aridisoles y Entisoles, las texturas varían desde arcillosas a arenosas, con fragmentos gruesos; los valores de pH oscilan entre 7,5 y 8,5 (CIL 1991, Apcarian et al. 2014). El clima se caracteriza por ser mesotermal (Thornthwaite 1948) y xerofítico seco (Papadakis 1980). Las precipitaciones, de régimen mediterráneo, alcanzan en promedio los 240 mm anuales; la evapotranspiración anual es de 790 mm, la temperatura media anual del aire es igual a 15,5 °C, el promedio de las temperaturas medias del mes más cálido (enero) corresponde a 24,2 °C y, el del mes más frío (julio), a 6,7 °C; los vientos son fríos y secos, de suaves a fuertes, frecuentes entre los meses de septiembre a diciembre, con velocidades superiores a 40 km/h y con dirección predominante del cuadrante Oeste-Suroeste (Rodríguez y Muñoz 2006, Rodríguez et al. 2014). La vegetación prístina del área de estudio corresponde a la del tipo matorral o estepa arbustiva xerófila, sammófila o halófila, perteneciente a la provincia fitogeográfica del Monte (Cabrera 1971).
Muestreo y recolección de datos
El inventario florístico se obtuvo a partir de 104 censos de vegetación, realizados de acuerdo con la metodología propuesta por Braun-B (1979), en parches salinizados, entre los años 2015 y 2019, durante las estaciones de otoño, primavera y verano, en establecimientos productivos (frutícolas, forrajeros y forestales) y distintos grupos textura-les propuestos por Bestvater y Casamiquela (1983): barda, media barda, media costa y costa. El área mínima de 10 m2 se determinó de acuerdo con Matteucci y Colma (1982) y el porcentaje de cobertura de cada taxón se cuantificó por apreciación visual. Para los cultivos cuyas plantas se hallaban dispuestas en hileras, los datos y el material vegetal se obtuvieron de la región interfilar salinizada (espacio ubicado entre dos hileras de cultivo que permite el tránsito de maquinaria agrícola).
Descripción de cultivos y grupos texturales de los suelos
Cultivos
Cultivo forrajero (Fo) (Material suplementario-Fig. 1a): comprende cultivos perennes como alfalfa (Medicago sativa L.), trébol rojo (Trifoliumpratense L.), agropiro (Thinopyrum ponticum (Podp.) Barkworth & D.R.Dewey), pasto ovillo (Dactylis glomerata L.), festuca alta (Festuca arundinacea Schreb.), entre otros, destinados a la alimentación de animales.
Pepita y forestal (Pe+F) (Material suplementario-Figs. 1b, c): los cultivos de pepita incluyen perales (Pyrus communis L.) y manzanos (Malus domestica (Suckow) Borkh.), los forestales corresponden, principalmente a sauce (Salix spp.) y álamo (Populus spp.). Todos estos cultivos son implantados en hileras.
Vid (V) (Material suplementario-Fig. 1d): algunas de las especies cultivadas corresponden a Vitis vinifera L. y V. labrusca L., implantadas en hileras.
Cultivo abandonado (CA) (Material suplementario-Fig. 1e): corresponde a los cultivos mencionados, pero fuera de producción.
Grupos texturales
Suelos de barda (B): predominan fracciones arenosas, en ocasiones muy gruesas, de origen aluvial. Presentan velocidades de infiltración muy rápidas y baja capacidad de retención de humedad. Bordean el sector Norte del Alto Valle.
Suelos de media barda (MB): dominan fracciones más finas, desde franco-limosas hasta francas, de origen coluvial en interacción con sedimentos aluviales provenientes de los ríos Neuquén y Negro. La velocidad de infiltración es menor y la capacidad de retención de humedad mayor con respecto a los suelos anteriores, por presentar partículas más finas, lo que favorece los problemas de salinización o sodificación.
Suelos de media costa (MC): incluyen texturas desde franco-arenosas a arenoso-francas. Su textura indica buena infiltración.
Suelos de costa (C): son de textura arenosa, con partículas de menor diámetro que las de los suelos de barda. Consiste en una franja interrumpida por causes del curso principal del río.
Identificación del material recolectado
El material recolectado se herborizó y determinó taxonómicamente mediante floras regionales (Correa 1971, 1978, 1984a, 1984b, 1988, 1999, Zuloaga et al. 2012, 2013, 2014, 2015, Zuloaga y Belgrano 2018, 2019) y publicaciones de Múlgura de Romero (1981, 1982), Kiesling (2003), Brignone et al. (2016) y Minué y Gandullo (2019). El material recolectado se cotejó además con el depositado en el Herbario Agronomía Región Comahue (ARC) y se ingresaron los taxones no registrados.
Composición y diversidad florística
Para cada taxón se indicó la familia botánica, el origen geográfico (exótico/nativo) y hábito (hierba/subarbusto/ arbusto) según Zuloaga et al. (2019), forma de vida (fanerófito/criptófito/terófito/hemicriptófito/caméfito) según Raunkiaer (1934) y su mecanismo de resistencia a la salinidad (pseudohalófito/crinohalófito/euhalófito) según Breckle (1976).
Para cada grupo textural y cultivo se calculó la riqueza, índice de Shannon-Wiener (H), índice de equitatividad de Pielou (J) y valor de importancia (VI) (Pielou 1975, Magurran 2004). Se indicaron aquellos taxones con un VI superior al 10 % en cada grupo textural y cultivo.
La comparación florística de los distintos grupos texturales y cultivos se efectuó mediante un análisis de agrupamiento jerárquico con el método UPGMA (unweighted pair group method with arithmetic mean) y matriz de similitud (índice de Morisita) desarrollados mediante el software Past 4.01 (Hammer et al. 2001).
RESULTADOS
Composición y diversidad florística
Se registró un total de 111 taxones infragenéricos, distribuidos en 26 familias y 84 géneros (Material suplementario-Tabla 1). El 71,2 % (79) son Dicotyledoneae mientras que el 28,8 % (32) restante, Monocotyledoneae. De los 111 taxones, 88 fueron determinados a nivel de especie, seis de subespecie y 17 de variedad. Las familias mejor representadas correspondieron a Poaceae (31, 27,93 %), Asteraceae (22, 19,82 %) y Chenopodiaceae (doce, 10,81 %). Los géneros con mayor riqueza de taxones fueron Atriplex, Hordeum y Polypogon, con cuatro cada uno.
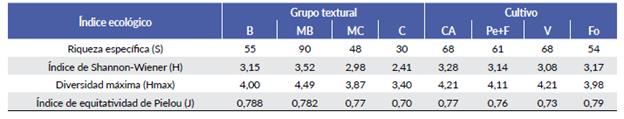
Tabla 1 Indices ecológicos obtenidos para cada grupo textural y cultivo. B= barda, MB= media barda, MC= media costa, C= costa, CA= cultivo abandonado, Pe+F= pepita y forestal, V= vid, Fo= cultivo forrajero.
El 58,56 % de los taxones fueron exóticos, mientras que el 41,44 %, nativos. El 70,27 % de los taxones correspondieron a pseudohalófitos, el 17,11 % a crinohalófitos y, el 12,62 % a euhalófitos. El 55,87 % (62) de los taxones correspondieron a terófitos, seguido por los hemicriptófitos (23,42 %, 26) y criptófitos (12,61 %, 14). En menor proporción se ubicaron los fanerófitos (4,5 %, 5) y caméfitos (3,6 %, 4). El 90,1 % (100) de los taxones correspondieron al tipo de hábito herbáceo, mientras que el 6,3 % (7) y el 3,6 % (4) a los hábitos arbustivo y subarbustivo, respectivamente.
Análisis florístico por grupo textural
Suelos de barda (B): presentaron una riqueza de 55 taxones, de los cuales el 52,7 % (29) fueron terófitos, el 63,6 % (35) pseudohalófitos, el 92,7 % (51) herbáceos y el 56,3 % (31) exóticos. Ocho taxones fueron exclusivos de este grupo textural (Material suplementario-Tabla 1). La especie con valor de importancia mayor al 10 % fue Atriplex prostrata Boucher ex DC. (11,66 %).
Suelos de media barda (MB): presentaron una riqueza de 90 taxones, de los cuales el 55,5 % (50) fueron terófitos, el 72,2 % (65) pseudohalófitos, el 93,3 % (84) herbáceos y el 64,4 % (31) exóticos. Los taxones exclusivos para este grupo textural fueron 31 (Material suplementario-Tabla 1). Ningún taxón superó el valor de importancia de 10 %.
Suelos de media costa (MC): presentaron una riqueza de 48 taxones, de los cuales el 47,9 % (23) fueron terófitos, el 62,5 % (30) pseudohalófitos, el 87,5 % (42) herbáceos y el 54,1 % (26) exóticos. Seis taxones fueron exclusivos de este grupo textural (Material suplementario-Tabla 1). El taxón con valor de importancia mayor al 10 % fue Leptochloa fusca (L.) Kunth subsp. fusca (14,95 %).
Suelos de costa (C): presentaron una riqueza de 30 taxones, de los cuales el 43,3 % (13) fueron terófitos, el 56,6 % (17) pseudohalófitos, el 86,6 % (26) herbáceos y el 56,6 % (17) exóticos. Dos taxones fueron exclusivos de este grupo textural (Material suplementario-Tabla 1). Los taxones con valores de importancia mayor al 10 % fueron Muhlenbergia asperifolia (Nees & Meyen ex Trin.) Parodi (14,51 %), Melilotus albus Desr. (11,85 %) y Malvella leprosa (Ortega) Krapov. (11,67 %).
El análisis florístico demostró que los suelos de barda y media barda fueron los que presentaron mayor riqueza específica y diversidad. Los valores más bajos de riqueza específica y diversidad se obtuvieron en los suelos de costa y media costa. Los valores de equitatividad para todos los grupos texturales se ubicaron entre 0,7 y 0,8 (Tabla 1).
Si bien en todos los grupos texturales predominaron los pseudohalófitos, el mayor número de crinohalófitos y euhalófitos se registró en el grupo textural de media barda. Se observó una tendencia al aumento del número de taxones de este tipo de halófitos en texturas finas.
Los taxones en común a los cuatro grupos texturales fueron: Atriplex prostrata, Bassia scoparia (L.) A.J. Scott, Cichorium intybus L., Conyza bonariensis (L.) Cronquist var. bonariensis, Cynodon dactylon (L.) Pers. var. dac-tylon, Distichlis spicata (L.) Greene var. spicata, Echinochloa crusgalli (L.) P. Beauv. var. crusgalli, Malvella leprosa, Melilotus albus, Plantago lanceolata L., Prosopis strombulifera (Lam.) Benth. var. strombulifera, Suaeda altissima (L.) Pall., Taraxacum officinale F.H. Wigg. y Tessaria absinthioides (Hook. & Arn.) DC.
Análisis florístico en cultivos
Cultivo forrajero (Fo): se registró una riqueza de 54 taxones para este cultivo. El 50 % (27) fueron terófitos, el 61,1 % (33) pseudohalófitos, el 90,7 % (49) herbáceos y el 57,4 % (31) exóticos. Dos taxones crecieron exclusivamente en este cultivo (Material suplementario-Tabla 1). Ningún taxón superó el valor de importancia de 10 %.
Pepita y forestal (Pe+F): se registraron 61 taxones para este cultivo. El 49,1 % (30) fueron terófitos, el 77 % (47) pseudohalófitos, el 91,8 % (56) herbáceos y el 63,9 % (39) exóticos. Nueve taxones crecieron exclusivamente en estos cultivos (Material suplementario-Tabla 1). El taxón con valor de importancia mayor al 10 % fue Atriplex prostrata (14,21 %).
Vid (V): se registró una riqueza de 68 taxones para este cultivo. El 51,4 % (35) fueron terófitos, el 64,7 % (44) pseudohalófitos, el 88,2 % (60) herbáceos y el 54,4 % (37) exóticos. Un total de 18 taxones crecieron exclusivamente en este cultivo (Material suplementario-Tabla 1). El taxón con valor de importancia mayor al 10 % fue Distichlis spi-cata var. spicata (11,58 %).
Cultivo abandonado (CA): se registró una riqueza de 68 taxones. El 55,8 % (38) fueron terófitos, el 64,7 % (44) pseudohalófitos, el 94,1 % (64) herbáceos y el 63,2 % (43) exóticos. Trece taxones crecieron exclusivamente en este ambiente (Material suplementario-Tabla 1). Ningún taxón superó el valor de importancia de 10 %.
El análisis floristico demostró que los cultivos abandonados y los forrajeros fueron los que presentaron mayor diversidad y equitatividad. La mayor riqueza especifica se presentó en cultivos abandonados y de vid. Por el contrario, la menor riqueza especifica se observó en los cultivos de pepita y forestales y forrajeros. Los valores de equitatividad para todos los grupos texturales se ubicaron entre 0,7 y 0,8 (Tabla 1).
Los taxones en común a los cuatro cultivos fueron: Atri-plex prostrata, Bassia scoparia, Chenopodiastrum murale (L.) S. Fuentes, Uotila & Borsch, Cichorium intybus, Convolvulus arvensis L., Conyza bonariensis var. bona-riensis, Cynodon dactylon var. dactylon, Distichlis spicata var. spicata, Echinochloa crus-galli var. crusgalli, Festuca arundinacea, Hoffmannseggia glauca (Ortega) Eifert, Hordeum euclaston Steud., Hordeum stenostachys Godr., Hypochaeris chondrilloides (A. Gray) Cabrera, Lactuca serriola L., Lepidium draba L., Leptochloa fusca subsp. fusca, Malvella leprosa, Medicago sativa, Plantago lanceolata, Polygonum aviculare L., Portulaca oleracea L., Prosopis strombulifera var. strombulifera, Sonchus ole-raceus L., Suaeda altissima, Taraxacum officinale, Tessa-ria absinthioides y Thinopyrum ponticum.
Similitud florística
Los valores de similitud florística resultaron, en general, de bajos a medios (Material suplementario, Tabla 2). Los suelos de barda y media barda fueron los más similares, con un valor de similitud de 0,68. Por el contrario, la menor similitud estuvo entre los suelos de media costa y los de costa, con un valor de 0,39 (Fig. 2a).
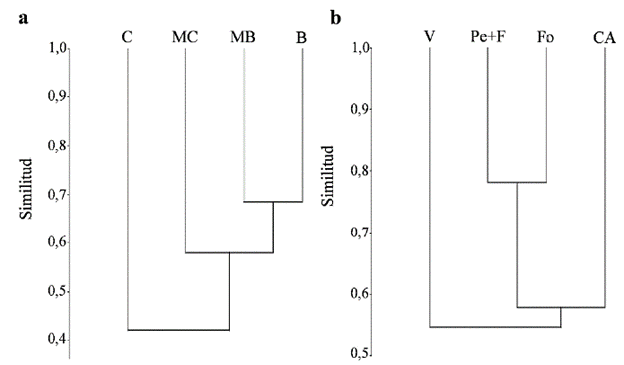
Figura 2 Dendrogramas de similitud mediante método UPGMA e índice de Morisita: a. para los distintos grupos texturales, correlación cofenética = 0,95. B= barda, MB= media barda, MC= media costa, C= costa. b. para los distintos cultivos, correlación cofenética= 0,96. CA= cultivo abandonado, Pe+F= pepita y forestal, V= vid, Fo= cultivo forrajero.
Los valores de similitud florística por cultivo fueron, en general, de medios a altos (Material suplementario, Tabla 3). Los cultivos de pepita y forestal y forrajeros presentaron los valores más altos de similitud, igual a 0,78. En contraste, los cultivos abandonados y de vid mostraron el valor de similitud más bajo, igual a 0,53 (Fig. 2b).
Novedades corológicas para la provincia de Río Negro
A partir del análisis florístico se identificaron ocho taxones no registrados para la provincia de Río Negro: Anoda cristata (L.) Schltdl. (Material suplementario-Figs. 2a, b), Atriplex suberecta I. Verd. (Material suplementario-Figs. 2c, d), Euphorbia serpens Kunth var. microphylla Müll. Arg., Oenothera curtiflora W.L. Wagner & Hoch (Material suplementario-Figs. 2e, f), Parapholis incurva (L.) C. E. Hubb., Polypogon exasperatus (Trin.) Renvoize var. kuntzei (Mez) Finot, Spergula salina (J. Presl. & C. Presl.) D. Dietr. (Material suplementario-Fig. 2g) y Thinopyrum ponticum (Material suplementario-Fig. 2h).
DISCUSIÓN
Las familias mejor representadas en este estudio (Poaceae, Asteraceae y Chenopodiaceae) son de las más frecuentes en ambientes salinos y sódicos (Walter 1961, Liphschitz y Waisel 1982, Schirmer y Breckle 1982, Breckle 1990, Le Houérou 1993, Cantero et al. 1998, Cobo-M 2001b, Vercelli et al. 2013, Hasanuzzaman et al. 2014, Mata-F et al. 2014). Estas familias también son señaladas por Cantero et al. (1996), Lendínez et al. (2011), Cantero et al. (2016) y Orrego et al. (2018) como las de mayor riqueza de taxones en suelos salinos.
El total de taxones inventariados por Cantero et al. (1996), Lendínez et al. (2011), Cantero et al. (2016) y Orrego et al. (2018) en ambientes salinos es muy variable (362, 162, 673 y 151 taxones, respectivamente, frente a 111 taxones hallados en este estudio). Las diferencias pueden estar dadas tanto por los aspectos ambientales y antrópicos propios de cada sitio, como por el tamaño de la superficie estudiada. San Martin et al. (1992) mencionan que una condición de salinidad extrema en ambientes poco antropizados se correlaciona con una baja riqueza específica, en coincidencia con los resultados de Teillier y Becerra (2003) (21 taxones), Gandullo (2004) (16 taxones), Gandullo y Gastiazoro (2009) (8 taxones), Karlin et al. (2012) (40 taxones) y Pawar et al. (2020) (12 taxones). Esto explicaría el alto valor de riqueza específica encontrado en ambientes salinos no extremos cultivados, altamente antropizados (111 taxones en este trabajo y 133 en Sánchez-B y Guevara-F 2013).
El predominio de Dicotyledoneae en ambientes salinos se condice con los resultados de Cantero et al. (1996), Lendínez et al. (2011), Cantero et al. (2016) y Orrego et al. (2018). De acuerdo con lo mencionado por Flowers y Col-mer (2008), esta tendencia podría deberse a que las halófitas dicotiledóneas, en general, toleran salinidades más amplias que las halófitas monocotiledóneas.
En regiones áridas, la proporción de las formas de vida es variable. Oztürk et al. (2008) concluyen que en ambientes cultivados la forma de vida dominante es la hemicriptófita, mientras que Conticello y Bustamante (2001) señalan la predominancia de terófitos, en coincidencia con lo hallado en el presente estudio en los suelos halomórficos cultivados del Alto Valle de Río Negro. La variabilidad en la proporción de las formas de vida puede ser explicada tanto por el grado de salinidad, con predominio de hemicriptófitos en suelos salinos extremos (San Martin et al. 1992, Cantero et al. 2016), como por el grado de antropización de los suelos salinos, donde los terófitos se comportan como pioneros y colonizadores (Espinar 2009, Lendínez et al. 2011).
De acuerdo con lo indicado por Bustamante et al. (2003), Cerazo y Conticello (2008), Kiesling (2009), Perissé et al. (2009), Zuloaga et al. (2012), Finot et al. (2013), Troiani (2016a, b), Puricelli et al. (2018) y Minué y Gandullo (2019), los taxones registrados aquí por primera vez para la provincia de Río Negro se comportan como invasores de suelos cultivados. En particular, Atriplex suberecta, Euphorbia serpens var. microphylla, Parapholis incurva, Spergula salina y Thinopyrum ponticum prosperan en suelos salinos o sódicos (Ragonese 1941, Cobo-M 2001a, b, Marchal et al. 2002, Gandullo y Gastiazoro 2009, Gandullo et al. 2010, Zuloaga et al. 2012, Vogt 2015, Brignone et al. 2016, Cantero et al. 2016 y Leaden et al. 2016).
La abundancia de taxones exóticos registrados en los distintos grupos texturales y en los cultivos se explica por los sucesivos disturbios antrópicos (labores culturales) realizados en el suelo. En este sentido, Sánchez et al. (c2008) y Karlin et al. (2017) indicaron que las labores de desmonte y nivelación del suelo, realizadas para el inicio de la actividad agropecuaria, han eliminado parcialmente la flora nativa característica de la región fitogeográfica del Monte y han promovido la instalación de especies exóticas, más tolerantes a la presión antrópica.
La mayor diversidad y riqueza observada en los cultivos abandonados y de vid se debe a la menor intervención antrópica que presentan, resultados coincidentes con los estudios de Cañadas-S (2008), en suelos agrícolas abandonados de ambientes mediterráneos semiáridos. En este sentido, si bien un suelo abandonado tiende a una estabilidad vegetal y edáfica (Cerda 2002) y se esperaría que en cultivos abandonados existiese un mayor número de taxones nativos y arbustivos o arbóreos, los altos valores de taxones herbáceos y exóticos hallados en el área de estudio podrían deberse al abandono reciente de las tierras cultivadas.
Los halófitos que toleran mejor la salinidad son aquellos que poseen mecanismos de control específicos y efectivos para mantener los niveles de iones constantes (Breckle 1986). En este sentido, el sistema radical de los pseudohalófitos no regula niveles constantes de iones por lo que tolera bajas concentraciones de sales y son sensibles al aumento de estas (Zahran 1982). En el presente estudio la mayoría de los taxones (70,27 %) son clasificados como pseudohalófitos y reportados como malezas de cultivos en suelos normales, es decir no salinos y no sódicos y de ambientes ruderales (Conticello y Gandullo 1991, Zegarra-Z 1996, Conticello y Bustamante 2001, Conticello et al. 2008, Sánchez et al. c2008, Zegarra-Z y Arévalo-S 2015). En consecuencia, los pseudohalófitos identificados en esta investigación no advertirían el proceso de salinización del suelo. En cambio, los crinohalófitos y euhalófitos (29,73 % restante) contribuirían al diagnóstico de un suelo salino o sódico, como es el caso de los suelos de media barda, donde se halla la mayor proporción de estos tipos de halófitos. Esto se debe a que la combinación de texturas arcillosas con capas freáticas salinas promueve la salinización progresiva de los suelos (Pepi et al. 1998).
La similitud florística, tanto en los diferentes grupos texturales como en cultivos, varía. Las diferencias florísticas manifestadas por los dendrogramas están relacionadas con las características de los grupos texturales estudiados y el grado de antropicidad (labores culturales) de cada sistema, de acuerdo con lo demostrado por Cardina et al. (1991), Mack et al. (2000), Mitja et al. (2008), Brun et al. (2012) y Sánchez-B y Guevara-F (2013). Sánchez-B y Gue-vara-F (2013) destacan especies preferenciales para determinadas propiedades fisicoquímicas del suelo, en cultivo de maíz, mientras que Mahmood et al. (1994) señalan una relación entre el grado de cobertura de determinadas especies espontáneas de campos salinizados y sodificados y las variables edáficas que caracterizan estos tipos de suelos (conductividad eléctrica y R.A.S.). En este sentido, el presente estudio evidencia la presencia de taxones exclusivos que prosperan en ciertos grupos texturales o sistemas cultivados o abandonados.
Consideraciones finales
Se considera, finalmente, que la composición florística, como sustrato de otras poblaciones y comunidades vegetales que enriquecen el agroecosistema, precisa futuros estudios que permitan evaluar el comportamiento ecológico de los taxones exclusivos y su posible empleo como bioindicadores de suelos halomórficos. Se hace especial mención que, si bien en el presente trabajo solamente se ha considerado el componente vegetal de parches salinizados influenciado por los grupos texturales, este nivel de precisión no se había examinado hasta el momento. Finalmente, al considerar el papel indicador de las especies vegetales (crinohalófitos y euhalófitos), es posible desentrañar algunos aspectos ligados directamente con la naturaleza del proceso de salinización que se da en los agroecosistemas y cultivos abandonados del Alto Valle de Río Negro. Por lo tanto, la comprensión e interpretación de este proceso permite la planificación y prevención para una gestión adecuada de los establecimientos productivos.

