











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los Humedales del Chaco argentino, declarados Sitio Ramsar (denominado de ese modo porque en Ramsar, Irán, fue firmada por primera vez la Convención Relativa a los humedales de Importancia Internacional), presentan un ambiente muy heterogéneo. El área corresponde a la planicie de inundación de los ríos Paraná, Paraguay, Bermejo, Negro, Salado y numerosos tributarios que conforman una extensa red hidrográfica que le confiere al paisaje rasgos singulares, tanto en su alta biodiversidad como en su dinámica dentro del ecosistema (Humedales Chaco c2019; (Chaco, www.argentina.gob.ar).
En la actualidad el avance de las actividades antrópicas en dicho sitio incrementa la reducción de bosques nativos, ello conduce a la homogeneización del ambiente e influye tanto en la disminución de la cantidad de especies que pueden coexistir como en la diversidad de gremios tróficos (Uetz 1991, Tews et al. 2004, Avalos et al. 2009). A nivel de familia taxonómica, como sucede con Thomisidae, en general se ven afectadas aquellas que son menos abundantes o que se encuentran representadas por unas pocas especies y que mayormente son reportadas con baja riqueza en los trabajos de diversidad de arañas en Argentina (Avalos et al. 2007, Rubio et al. 2008, Avalos et al. 2009, Avalos et al. 2013, Achitte-Schmutzler et al. 2016, Nadal et al. 2018).
Las Thomisidae, llamadas arañas cangrejo, no fabrican telas para cazar, viven casi exclusivamente en ramas, sobre hojas o flores y se encuentran dentro del gremio cazadoras por emboscada (Cardoso et al. 2011). Existen diversos trabajos orientados a redescribir y escribir diferentes especies de tomísidos y a resolver las relaciones filogenéticas intra-grupo. Al respecto se puede citar a Benjamin et al. (2008); Benjamin (2011, 2013, 2015, 2017); Sirvid et al. (2013); Silva-Moreira y Machado (2016); Machado et al. (2019a, 2019b, 2019c); Grismado y Achitte-Schmutzler (2020) y Machado y Texeira (2021) entre otros. Los conocimientos acerca de su ecología, taxonomía y diversidad aún son incipientes en Argentina, actualmente se conocen 60 especies (CAA c2018) mientras que a nivel mundial se registran 2160 especies de Thomisidae (WSC c2018).
La transformación y fragmentación de bosques representan las principales causas de la pérdida de diversidad biológica a nivel mundial (Myers et al. 2000, Silva-Matos et al. 2005). Consecuentemente, los estudios de la biodiversidad pasaron a ser fundamentales en muchas investigaciones enfocadas a la ecología o en la búsqueda de estrategias para la conservación de especies.
Clarke y Warwick (1998, 2001) proponen varios índices basados en las relaciones taxonómicas entre las especies, dichos índices cuantifican y promedian el grado de relación de un ensamble respecto a otro. Para estos autores es importante la estructura filogenética y sostienen que un ensamble compuesto por un grupo de especies cercanamente emparentadas debe ser considerado como menos diverso que un ensamble de igual riqueza específica, pero con relaciones filogenéticas más distantes entre sus especies.
El índice de diversidad taxonómica es utilizado para analizar y comparar las comunidades de diversos organismos entre distintos hábitats, localidades y ecorregiones (Moreno et al. 2009, Cruz-Elizalde et al. 2014, Capetillo-Piñar et al. 2015, Hidalgo et al. 2015, García-de Jesús et al. 2016, Kong et al. 2018). Con respecto a la fauna de arañas se citan los trabajos de Rubio y Moreno (2010) quienes utilizan este índice para evaluar los ensambles de Araneidae en diferentes localidades y hábitats de la reserva Iberá (humedales Ramsar); y el de Rubio (2016) quien evalúa los ensambles de Salticidae en las ecorregiones de la provincia de Misiones, Argentina.
La estructura de la vegetación es un factor importante que influye en la diversidad de arañas (Jiménez-Valverde y Lobo 2006), esto es, a mayor complejidad estructural vegetal, mayor riqueza específica, debido a la biomasa del follaje y la disponibilidad de presas (Hore y Uniyal 2008). En este sentido, ello podría influir en la fauna de Thomisidae que comúnmente abundan en áreas naturales (Podgaiski et al. 2007, Ricetti y Bonaldo 2008, Rubio et al. 2008) con varias especies típicas del follaje.
En este trabajo se analiza por primera vez la estructura taxonómica de los ensambles de tomísidos en ambientes con distintos grados de complejidad vegetal como pastizales, palmares y bosques, del sitio Ramsar Humedales Chaco, se espera que los ambientes con mayor complejidad vegetal presenten una estructura taxonómica balanceada, es decir, con mayor grado de representantes en los niveles taxonómicos superiores.
MATERIALES Y MÉTODOS
Área de estudio
El área de estudio integra la lista Ramsar, desde febrero de 2004, con el nombre Humedales Chaco, y se encuentra situada en la provincia del Chaco, Argentina. El sitio tiene una superficie de 508 000 ha (Humedales Chaco c2019; (Chaco, www.argentina.gob.ar), abarca la franja oriental de la Provincia del Chaco e incluye los Departamentos San Fernando, Primero de Mayo y Bermejo. Limita al Norte por el cauce del Río Bermejo, al Sur el paralelo 28°, al Este por los cauces de los ríos Paraná y Paraguay y al Oeste por el trazado de la Ruta Nacional N°11. Biogeográficamente el área está incluida en la Región Neotropical, Subregión Chaqueña, que corresponde el Centro y Norte de Argentina, Sur de Bolivia, Oeste de Paraguay y Sureste de Brasil (Morrone 2001).
Trabajo de campo
Los muestreos se efectuaron en siete localidades del sitio Ramsar Humedales Chaco (Fig. 1), en las unidades ambientales Selva en galería (SG), Bosque bajo abierto (BA), Pastizal (Pz) y Palmar (Pr) en los meses de noviembre a marzo durante los años 2013 a 2016. En cada localidad se efectuaron dos muestreos: en bosques, pastizales y palmares se realizaron en noviembre de 2013, y en marzo de 2015; en las selvas en galerías, pastizales y palmares en noviembre de 2014 y en marzo de 2016.
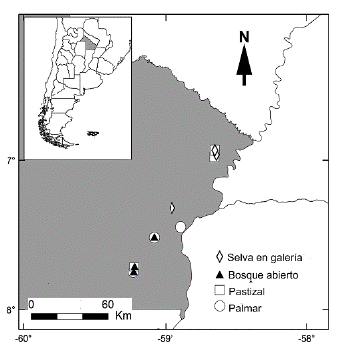
Figura 1 Localización de los sitios de muestreos en el sitio Ramsar Humedales Chaco, provincia del Chaco, Argentina.
En todos los sitios se trazaron tres transectos de 200 m de longitud por 2 m de ancho, separados entre sí por 500 m; dentro de los mismos se seleccionaron cinco puntos distanciados entre sí por 50 m. En cada uno de esos puntos se recolectaron arañas aplicando las siguientes técnicas de captura: en bosques y selvas: golpeteo de follaje y captura directa diurna; en pastizales y palmares, aspirado y captura directa diurna. De este modo se buscó obtener una mejor representación de la comunidad de arañas (Hodge et al. 2007).
Golpeteo de follaje
Consistió en quince golpes sobre la vegetación arbustiva y en la porción baja del estrato arbóreo, el material se recolectó sobre un lienzo blanco cuadrado de 2,50 m de lado.
Captura directa diurna
Ocurrió durante diez minutos de observación por cada punto de muestreo por transecto. Tanto en bosques como en pastizales. A fin de agilizar los muestreos, cinco personas participaron, ubicadas en los transectos y una persona por punto de muestreo; al cabo de diez minutos nos trasladábamos al segundo transecto y así al tercero.
Captura por aspiración
Se efectuó mediante una aspiradora de jardín G-vac (Mod. 220 V-AR) sobre la vegetación. El aspirador posee un tubo de 1,10 m de longitud y 12 cm de diámetro (caudal 710 m3/h), cada muestra de succión de la vegetación se tomó en 4 m2 durante un minuto.
Se obtuvieron quince muestras por cada técnica de captura, es decir 30 muestras en cada unidad ambiental y 720 muestras en el total del muestreo (cuatro ambientes por tres localidades, dos técnicas (por ambiente), quince muestras, dos repeticiones).
Cada muestra fue considerada independiente y se rotularon indicando la técnica utilizada, fecha y el ambiente de muestreo. Todos los ejemplares fueron colocados en frascos con etanol 70 % y depositados en la colección CAR-TROUNNE del Laboratorio de Biología de los Artrópodos, Universidad Nacional del Nordeste (UNNE), Corrientes Argentina.
Análisis de datos
La diversidad taxonómica se analizó mediante los índices propuestos por Warwick y Clark (1995), los cuales calculan la distancia taxonómica promedio (Δ+), es decir mide la separación taxonómica promedio entre dos especies seleccionadas al azar en una comunidad; y la variación taxonómica promedio (Λ +). Ambos índices incluyen en su cálculo las distancias entre los linajes filogenéticos, son independientes del tamaño de la muestra y se pueden relacionar con la diversidad funcional y posibles impactos antropogénicos en el ecosistema (Warwick y Light 2002, Sathianandan et al. 2011).
Las categorías taxonómicas utilizadas para el análisis de diversidad taxonómica fueron las siguientes: Subfamilia, Tribu, Género y Especie/morfoespecie. Las morfoespecies fueron individuos adultos que se diferenciaron por las estructuras de sus genitales. Debido a que hasta el momento las relaciones filogenéticas del grupo no están debidamente establecidas y no se elaboraron nuevas propuestas taxonómicas formales (Benjamin et al. 2008, Benjamin 2011, Ramírez 2014, Wheeler et al. 2016, Machado et al. 2019b), se optó por la clasificación de subfamilias y tribus de Thomisidae con base en los trabajos de Ono (1988) y Lehtinen (2003). Este criterio se ve respaldado, además, en el análisis del árbol filogenético propuesto por Wheeler et al. (2016), en el mismo se puede observar que los agru-pamientos obtenidos se corresponden con las tribus, así los autores citados conservan la subfamilia Aphantochilinae; en una rama como hermanos están los géneros Tmarus y Titidius (Tamarini), Thomisus (Thomisini), por fuera del grupo que incluye a Misumenoides, Misumenops y Run-cinioide (Misumenini). En otra rama separada se ubica Diaea (Diaenini), y por último, los Stephanopinae quedan claramente separados del resto, las especies de Epicadus forman el grupo bien definido "Epicadus" (Silva-Moreira y Machado 2016, Machado y Texeira 2021).
La diversidad taxonómica utiliza la clasificación jerárquica de Linneo como un sustituto de las filogenias (Pérez-Hernández 2019) sin desconocer que estas se modifican constantemente en función de la filogenia (Ellingsen et al. 2005). Sin embargo, son una aproximación simple y discreta a una filogenia completa, inclusive aun cuando la taxonomía es la única información disponible, esta puede interpretarse como representativa de las relaciones filogenéticas (Moreno et al. 2009).
Para el cálculo de estos índices se utilizó el software PRI-MER-E versión 5, a partir de 1000 aleatorizaciones (Clarke y Gorley 2001).
Se evaluó la disimilitud de especies (1-PJ) con base en el índice de similitud de Jaccard (J) (Moreno 2001), donde PJ es la proporción que representan las especies compartidas de la riqueza total de las dos comunidades. A partir del valor de similitud se calcula el de disimilitud aplicando 1-PJ; este índice varía de cero, cuando la composición de especies es idéntica, hasta uno, cuando la composición de especies es totalmente diferente (García-de Jesús et al. 2016).
También se evaluó la disimilitud taxonómica (1-ΔT), donde ΔT es el índice de similitud taxonómica propuesta por Izsák y Price (2001) y modificada por Bacaro et al. (2007). Este índice mide la proporción que representan los taxo-nes compartidos del número total de taxones en dos sitios. Los valores de disimilitud taxonómica (1-ΔT) van de cero, cuando la estructura taxonómica del conjunto de especies de ambas comunidades es idéntica, a uno cuando la estructura taxonómica de las especies de ambas comunidades es totalmente diferente (Bacaro et al. 2007, García-de Jesús et al. 2016).
Además, como complemento, se realizó un análisis de ordenación no paramétrica NMDS (Nonmetric Multidimensional Scaling) con los valores obtenidos de 1-βJ y 1-ΔT a fin de ubicar y comparar la posición relativa de los ensambles de Thomisidae (García-de Jesús et al. 2016). De acuerdo con estos autores, el NMDS jerarquiza los valores de disimilitud entre pares de comunidades en un procedimiento iterativo para ubicar a cada comunidad en un espacio de dos dimensiones que mantiene las distancias entre comunidades, como un reflejo de su disimilitud. Para el análisis del NMDS se utilizó el índice de Bray-Curtis.
Por último, para calcular el efecto de las distancias geográficas respecto a la variación de la composición de los ensambles, se aplicó el test de Mantel el cual evalúa la correlación entre matrices de distancia (Sokal y Rohlf 1995).
Los análisis de correlación, de similitud de Jaccard, el NMDS y el test de Mantel se realizaron mediante el software PAST versión 2.16 (Hammer et al. 2001).
RESULTADOS
En total se registraron 464 ejemplares, de los cuales fueron utilizados para el análisis 288 individuos, por ser adultos y juveniles identificables taxonómicamente. Se identificaron 34 especies/morfoespecies pertenecientes a cinco subfamilias, siete tribus y diez géneros (Tabla 1). Cada una de dichas categorías taxonómicas estuvo mayormente representada en la selva en galería.
Tabla 1 Subfamilias, tribus, géneros y riqueza de Thomisidae recolectados en cada unidad ambiental del sitio Ramsar Humedales Chaco, Argentina.
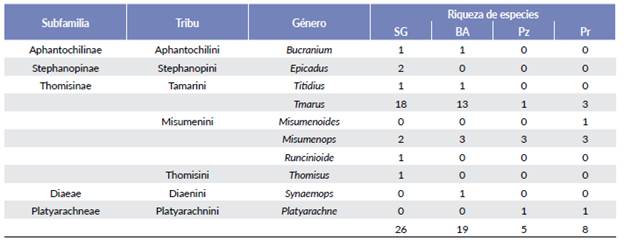
SG= selva en galería, BA= bosque bajo abierto Pz= pastizal y Pr= Palmar.
En cuanto a la distancia taxonómica promedio (Δ +) los pastizales (Pz) y palmares (Pr) estuvieron por encima del promedio esperado (Δ+=49), mientras que la selva en galería (SG) y el bosque bajo abierto (BA) mostraron una diversidad taxonómica menor a la esperada (Fig. 2). El mayor valor de este índice se obtuvo en el pastizal (Pz Δ +=56) pese a la menor riqueza específica registrada en este ambiente; sin embargo, estas diferencias no fueron significativas ya que todos estos valores cayeron dentro del contorno pro-babilístico (Fig. 2).
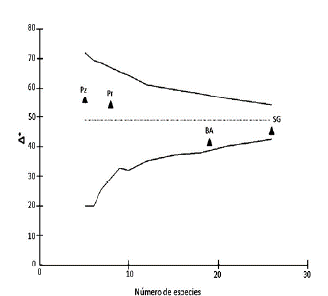
Figura 2 Distancia taxonómica promedio (Δ+) de los ensambles de Thomisidae en los ambientes muestreados del Sitio Ramsar Humedales Chaco, con límites de confianza del 95 %. SG= selva en galería, BA= bosque bajo abierto, Pz= pastizal y Pr= Palmar.
Respecto a la variación taxonómica promedio (Λ +), la Selva en galería presentó el mayor valor lambda (SG, A+=689,44) seguido por el pastizal (PZ, Λ +=624), ambos estuvieron por encima del promedio esperado por azar (Λ + = 449). Nuevamente, estos valores estuvieron dentro de los contornos probabilísticos de la distribución esperada (Fig. 3).
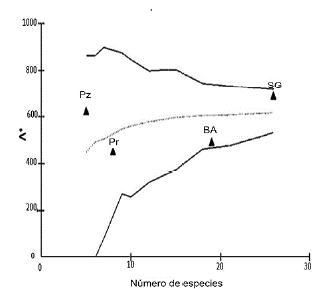
Figura 3 Variación Taxonómica Promedio (Λ +) de los ensambles de Thomisidae en los ambientes muestreados del Sitio Ramsar Humedales Chaco, con límites de confianza del 95%. SG= selva en galería, BA= bosque bajo abierto, Pz= pastizal y Pr= Palmar
Pese a que el pastizal y el palmar (Pr) presentaron la menor riqueza específica respecto a los otros ambientes, sus valores de Δ+ fueron elevados.
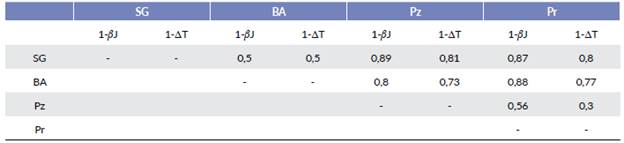
Los resultados de la disimilitud de especies (1-βJ) y de la disimilitud taxonómica (1-ΔT) entre las localidades se muestran en la Tabla 2. En la misma se observa que la mayor disimilitud tanto en la composición de especies como en la estructura taxonómica se obtuvo entre la selva en galería y el pastizal, mientras que los ambientes más similares fueron ambos bosques entre sí y el pastizal con el palmar.
Tabla 2 Diversidad beta basada en los valores de disimilitud de especies (1-(βJ) y en los de disimilitud taxonómica (1-ΔT) entre los ensambles de Tho-misidae de cada unidad ambiental del sitio Ramsar Humedales Chaco, Argentina.
En general los análisis de NMDS agruparon a los ambientes boscosos por un lado y por el otro los pastizales con los palmares, con valores óptimos de estrés para la disimilitud de especies (estrés = 0,05) y para la disimilitud taxonómica (estrés=0,07) (Figs. 4a y b).

Figura 4 Ordenación no paramétrica multidimensional (NMDS) de los ensambles de Thomisidae en distintas unidades ambientales del sitio Ramsar Humedales Chaco, a con base en la disimilitud de especies (1-(βJ) y b con base en la disimilitud taxonómica (1-ΔT). SC= Estancia San Carlos, GV= General Vedia, CB= Colonia Benitez, AN= Antequeras, SF= Estancia San Francisco, LQ= Estancia La Querencia, MZ= Estancia María Zaida. Ambientes: SG= selva en galería, BA= bosque abierto, Pz= pastizal, Pr= palmar.
En cuanto a la composición específica en los ambientes boscosos, el NMDS mostró dos agrupamientos, la selva en galería de General Vedia y Colonia Benítez (GV-SG, CB-SG) se agruparon con el bosque abierto de María Zaida (MZ-BA) y el bosque abierto de San Francisco con la selva en galería de San Carlos (SF-BA, SC-SG) (Fig. 4a), no obstante, en cuanto a su estructura taxonómica no se observaron agrupamientos, y hubieron cambios en la similitudes entre los sitios por ejemplo CB-SG mostró mayor similitud a las selvas en galería de Antequeras (AN-SG) (Fig. 4b).
Por su parte, el bosque abierto de La Querencia (LQ-BA) fue similar a los pastizales y a los palmares tanto a nivel de especies como a nivel de estructura taxonómica.
Mediante el test de Mantel se obtuvo que la distancia geográfica entre los sitios no difiere en forma significativa (R= 0,27, P = 0,06).
DISCUSIÓN
La riqueza de Thomisidae registrada en los ambientes boscosos del Sitio Ramsar Humedales Chaco fue alta en comparación con otros estudios (Avalos et al. 2007, Rubio et al. 2008); sin embargo, se obtuvo una baja diversidad taxonómica (Δ+) en dichos ambientes a diferencia de lo reportado por Rubio y Moreno (2010). Al respecto, Moreno et al. (2009) afirman que valores bajos de A+ implican que las relaciones filogenéticas de las especies son más cercanas a bajo nivel taxonómico. De este modo, nuestros resultados se correspondieron con el hallazgo de varias especies de Tmarus, llegando a representar más de la mitad del total de las especies en la selva en galería.
Asimismo, estos autores sostienen que los valores altos de variación taxonómica (Λ +) se relacionan con una sobre o subrepresentación de algunos grupos taxonómicos, de allí que el alto valor de Λ + implica un árbol filogenético poco uniforme, esto también se correspondería con los valores observados en las selvas en galerías en este estudio.
En concordancia con Clarke y Warwick (2001) quienes sostienen que cuantas más especies pertenecientes a diferentes géneros y familias haya en un lugar, mayor será el valor de Δ+ y por lo tanto mayor será la diversidad taxonómica, en los ambientes de pastizales y palmares se observaron mayor Δ+ y, a pesar de que se obtuvo baja riqueza en estos ambientes, las especies estuvieron uniformemente representadas en el árbol filogenético.
Asimismo, Moreno et al. (2009) aseveran que una alta diversidad taxonómica se esperaría para una comunidad con una distribución uniforme de las especies entre los niveles taxonómicos y unidades, situación que daría lugar a un bajo valor de Λ +; en este sentido y de acuerdo con los resultados obtenidos, el palmar sería el ambiente de mayor diversidad taxonómica.
A menudo surgen discrepancias entre los valores de riqueza y los índices de diversidad taxonómica, por ejemplo, Argañaraz et al. (2017) obtuvieron mayor riqueza de especies de saltícidos en uno de los hábitats muestreados, pero tanto la Δ+ como la Λ + mostraron valores bajos. Al respecto, en este trabajo no se obtuvo una correlación entre la riqueza y la diversidad taxonómica en coincidencia con los resultados de García-de Jesús et al. (2016), quienes evaluaron las comunidades de escarabajos en México. Esto reivindica la importancia del análisis de diversidad taxonómica como un enfoque diferente y que además integra la historia evolutiva y biogeográfica del taxón (Webb et al. 2002).
Por otra parte, la elevada diferencia tanto taxonómica (1-ΔT), como a nivel de especies (1-βJ), entre los bosques y entre pastizales-palmares indican que los ensambles son distintos entre estos ambientes. Resultados similares fueron reportados por Rubio y Moreno (2010) quienes analizaron los ensambles de arañas orbiculares en la Reserva Iberá, en dicho trabajo se obtuvo valores elevados de complementariedad entre los bosques y pastizales. Además, estos autores, con técnicas multivariadas obtuvieron agru-pamientos más cerrados de los pastizales mientras que los bosques presentaron una ordenación más dispersa; esta tendencia no se observó en este estudio en el análisis de NMDS con base en la disimilitud de especies.
Excepcionalmente el bosque de La Querencia mostró mayor similitud a los pastizales y palmares, lo que podría ser debido al deterioro de dicho bosque por el constante ingreso del ganado, en comparación con los demás bosques, éste fue el más degradado (observación personal). La estructura de la vegetación es un componente clave para las arañas que habitan en estratos arbóreos no degradados particularmente para las cazadoras por emboscada (Rodrigues y Mendonça Jr 2012); no obstante, Schirmel et al. (2016) sostienen que los tomísidos presentan elevada tolerancia en los ambientes perturbados o con una estructura vegetal simple. Al respecto en este trabajo el bosque de la Querencia presentó pocas especies y pocos representantes de niveles taxonómicos superiores, con una estructura específica y taxonómica similar al de los ambientes con menor complejidad estructural, como los pastizales y los palmares.
Los análisis taxonómicos pueden ser representativos de las relaciones filogenéticas (Pérez-Hernández 2019) las cuales han sido de gran utilidad en el estudio de comunidades para establecer los procesos que determinan los ensambles de comunidades en escalas espaciales y temporales distintas (Webb et al. 2002, Cavender-Bares et al. 2009). Asimismo, Webb et al. (2002) propusieron que el patrón de las relaciones filogenéticas entre especies puede reflejar los mecanismos que hacen posible su coexistencia dentro de una comunidad.
En este sentido, entre las especies más emparentadas dentro de una comunidad, como las numerosas especies de Tmarus obtenidos en los ambientes boscosos, el mecanismo de ensamblaje posiblemente esté mediado por filtros ambientales en donde las condiciones bióticas favorecen la coexistencia de especies con requerimientos y características similares, las cuales les permiten sobrevivir en determinados tipos de hábitats (Velásquez-Puentes y Bacon 2016). Así las especies cercanamente emparentadas son más similares en cuanto a sus requerimientos ecológicos que las especies lejanamente emparentadas (Peterson et al. 1999). Asimismo, un patrón filogenético en donde las especies de una comunidad están lejanamente relacionadas resaltaría el papel de la competencia entre especies como mecanismo principal del ensamblaje, ya que sólo especies con requerimientos ecológicos distintos podrían coexistir (Velásquez-Puentes y Bacon 2016).
La falta de correlación entre la distancia geográfica y la similitud de los ensambles de tomísidos de cada unidad ambiental coincide con lo reportado en otras investigaciones referidas a las arañas (Bonaldo et al. 2007) y otros organismos (Bacaro et al. 2012, García-de Jesús et al. 2016). Por su parte Rodríguez-Artigas et al. (2016) sostienen que las distancias geográficas no son un factor relevante en la estructura de los ensambles de Thomisidae. Al respecto, en este trabajo la distancia geográfica no estaría afectando la similitud entre los sitios muestreados.
Así, las diferencias entre los ensambles podrían ser el resultado de otros factores microambientales, como así también la estructura de la vegetación, la competencia y la disponibilidad de alimentos y refugio; que a lo largo del tiempo evolutivo posiblemente influyeron en la estructura taxonómica de los ensambles de tomísidos.
En general, en este trabajo se mostró que los ensambles de tomísidos son diferentes en las unidades ambientales muestreadas, con una estructura taxonómica en mayor o menor grado distinta entre sí. La inclusión de las relaciones entre los taxones en los análisis de diversidad podría ser clave a la hora de implementar planes de manejo y conservación. La franja occidental de la Provincia del Chaco fue declarada de interés para la conservación, sin embargo, continúa el incremento de la degradación de los hábitats naturales, lo que ha fortalecido la necesidad de conocer y evaluar los patrones de biodiversidad.

