











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Las aves limícolas (Aves: Charadriiformes) son un grupo acuático numeroso a nivel mundial que se caracterizan por sus largas migraciones que llegan a conectar continentes o hemisferios (van de Kam et al. 2004, Gill Jr et al. 2005, Stuuds et al. 2017). Las extensas distancias recorridas anualmente y la consiguiente dependencia de una amplia variedad de humedales han condicionado que en la actualidad las aves limícolas estén sujetas a varias amenazas. Por ejemplo, son susceptibles a los efectos de la expansión humana y la consecuente modificación de los hábitats, la sobreexplotación y cambio climático (Morrison et al. 2001, Sutherland et al. 2012, Melville et al. 2016). Diversos estudios han evidenciado una alarmante disminución a nivel mundial de varias especies de aves limícolas tanto en áreas de cría, como de paso e invernada (Morrison et al. 2006, Bart et al. 2007, Clemens et al. 2016, Stuuds et al. 2017). Este hecho ha constituido una alarma regional sobre la cual se han volcado numerosos esfuerzos de conservación.
Los humedales cubanos son los más extensos dentro del área del Caribe (Aguilar et al. 2019) lo que unido a la posición geográfica de Cuba los convierten en áreas potencialmente utilizables por las aves acuáticas durante sus migraciones anuales. Varios estudios dentro de la isla evidencian como este tipo de ecosistema constituye un hábitat clave para las aves limícolas durante la migración (otoñal y primaveral) y periodo de residencia invernal para el descanso y la alimentación (Jiménez 2006, Aguilar et al. 2019). Dentro de los aspectos ecológicos más estudiados sobre las aves limícolas en la isla se encuentran: dinámica poblacional (Acosta et al. 1992, Blanco 2006, González y Jiménez 2011) y morfometría (García-Lau et al. 2012, Jiménez et al. 2015a). Sin embargo, no existe información sobre la disponibilidad y uso de recursos tróficos de este grupo de aves dentro de los humedales naturales que utilizan. Las escasas investigaciones sobre la ecología trófica de aves limícolas en Cuba están restringidas a humedales agrícolas como los arrozales (Acosta et al. 1994, Mugica et al. 2001), donde las variables abióticas, así como el tipo y abundancia de presas difieren de aquellas en hábitats naturales. Las arroceras son ecosistemas de agua dulce lo que representa una diferencia marcada con algunos humedales naturales como las llanuras intermareales donde los valores de salinidad llegan a sobrepasar la del mar.
Se ha descrito que la disponibilidad de alimento en los sitios de invernada puede regular la distribución y dinámica de las poblaciones de aves limícolas migratorias (Sprague et al. 2008, Jiménez et al. 2015b). Por tanto, comprender qué tipo de recursos tróficos están disponibles en los humedales naturales cubanos y cuáles de ellos son consumidos por las aves limícolas constituye un aspecto indispensable para posibles acciones de manejo y conservación en Cuba. En este estudio se analiza la ecología trófica de Calidris minutilla (Vieillot, 1819), Calidris pusilla (Linnaeus, 1766) y Calidris mauri (Cabanis, 1857). Estas tres especies de aves limícolas se encuentran entre las especies más pequeñas del género Calidris, se asocian en grandes bandos mixtos para forrajear, y dentro de los Calidris que utilizan la isla son los más comunes y abundantes (Jiménez et al. 2006, Acosta et al. 2011). Con este fin, se proponen como objetivos: 1) caracterizar la dieta de las tres especies en dos humedales naturales y 2) evaluar la relación entre la dieta de las tres especies y los recursos tróficos disponibles en el hábitat de alimentación.
MATERIALES Y MÉTODOS
Áreas de estudio
El estudio se realizó en dos importantes humedales naturales de la región central de Cuba: Río Máximo y Tunas de Zaza. Río Máximo se encuentra en la costa noreste de la provincia de Camagüey (21°44' Norte 67°31'Oeste) y Tunas de Zaza se ubica en la costa sur de la provincia Sancti Spíritus (21°38' Norte 79°32'Oeste) (Fig. 1). Ambas localidades son áreas protegidas incluidas en el Sistema Nacional de Áreas Protegidas bajo la categoría de Refugio de Fauna (CNAP 2004). Específicamente, la investigación se desarrolló en el hábitat de lodazal intermareal presente en ambas localidades, donde C. minutilla, C. pusilla y C. mauri forman bandos mixtos durante las actividades de forrajeo y descanso.
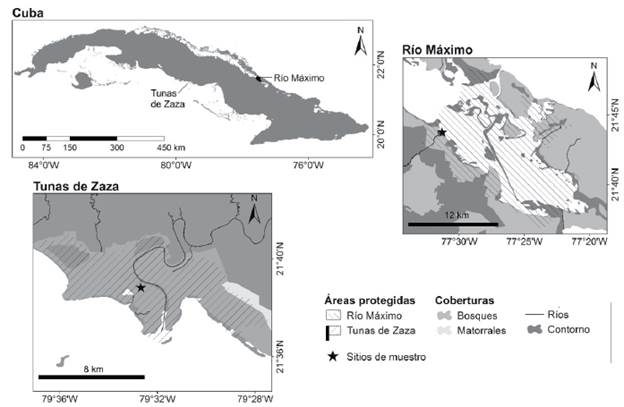
Figura 1 Ubicación geográfica de los Refugios de Fauna Río Máximo y Tunas de Zaza donde fue realizado el presente estudio.
En general ambas localidades presentaron similitudes en la temperatura del sedimento (Río Máximo: 26,0 ± 2,3 °C; Tunas de Zaza: 25,8 ± 2,2 °C) y el porcentaje de sedimento fino (Río Máximo: 72,1 ± 22,3 %; Tunas de Zaza: 84,0 ± 8,3 %) (U de Mann-Whitney, U = 190,0; P = 0,787; N = 20 y U = 30,0; P = 0,354; N = 20, respectivamente). La salinidad en cambio fue significativamente mayor en Tunas de Zaza (45,9 ± 3,5 que en Río Máximo (17,5 ± 11,6 %o) (U = 0,5; P < 0,001; N = 20). La temperatura fue tomada con un termómetro VWR L37823 (± 0,5 °C) y la salinidad utilizando un refractómetro portátil Vista (± 1 ppt). Por su parte, las muestras de sedimento para análisis granulométricos fueron recolectadas con una jeringuilla modificada con una abertura de 26 mm de diámetro interno (González 2011). La entrada a las áreas de trabajo y las recolecciones de individuos fueron aprobadas por el Centro de Inspección y Control Ambiental (CICA) mediante los permisos ordinario y de recolección No.2008/05 y No.47/2008, respectivamente.
El muestreo se realizó en el mes de octubre de 2008 (11 -18 octubre) durante la migración otoñal de las aves limícolas. Varios trabajos han evidenciado la importancia de los humedales cubanos, tanto naturales como antrópicos, durante esta etapa para este grupo de aves (Acosta et al. 1992, Acosta et al. 2002, González y Jiménez 2011). De igual modo, previas observaciones anecdóticas en los humedales estudiados constataron la presencia de altos valores poblacionales de aves limícolas durante la migración otoñal. El horario de estudio en los dos sitios fue de 0800 a 1030.
Caracterización de la dieta
La descripción de la dieta se realizó a partir de contenidos estomacales de individuos de C. minutilla, C. pusilla y C. mauri recolectados utilizando redes de niebla de 12 m y 18 m de largo por 2,5 m de alto. Para garantizar registros más completos de la alimentación de estas especies, las aves fueron capturadas al menos una hora después de iniciada la actividad de forrajeo dentro del área. Los individuos fueron sacrificados siguiendo el método de compresión torácica (Winker 2000), más recientemente conocido como compresión cardíaca rápida (Engilis Jr et al. 2018). Este método fue seleccionado considerando: 1-el pequeño tamaño de las aves estudiadas, 2-que el sacrificio fue realizado durante el trabajo de campo y 3-que se iban a utilizar las pieles para el museo y órganos como el hígado para otros estudios. La abundancia total de las tres especies de Calidris fue mayor en Río Máximo que en Tunas de Zaza (7990,0 ± 4256,2 ind./km2 y 1943,3 ± 1630,8 ind./km2, respectivamente), y en ambos sitios la proporción en los bandos de C. minutilla estuvo por encima de 70 % (González 2011). Teniendo en cuenta esto, fueron sacrificados 43 individuos en Río Máximo (C. minutilla N = 15; C. pusilla N = 12 y C. mauri N = 16) y 33 individuos en Tunas de Zaza (C. minutilla N = 22; C. pusilla N = 7 y C. mauri N = 4). Los especímenes fueron etiquetados y conservados en una nevera para su posterior análisis en el laboratorio.
Evaluación de la disponibilidad de presas
La disponibilidad de presas se evaluó mediante la toma de muestras de sedimento. Dado que el estudio estaba dirigido a identificar los recursos alimentarios de C. minutilla, C. pusilla y C. mauri, se tomaron muestras de sedimento dentro de las áreas utilizadas por bandos mixtos de forrajeo de más de 1000 individuos. Una vez identificada el área de forrajeo, se designó un área lo suficientemente amplia (5 ha) como para poder albergar tres transectos de 200 m de longitud, perpendiculares a la línea de costa y separados 100 m entre sí. Dicha metodología fue diseñada para este estudio basado en las características de ambas localidades.
Las muestras de sedimento se extrajeron en puntos aleatorios ubicados a cada lado de estas líneas utilizando una jeringuilla (50 CC) con una abertura de 26 mm de diámetro interno similar a la empleada por Mathot et al. (2010). Previamente, el extremo apical fue eliminado para poder introducir la muestra. La jeringuilla fue enterrada a una profundidad de 10 mm la cual constituye el estrato más accesible para las tres especies de Calidris según sus longitudes de pico (16,0 - 29,7 mm; García-Lau et al. 2012). En total se tomaron 20 muestras de sedimento en cada humedal. Todas las muestras fueron guardadas en bolsas de nylon previamente rotuladas y conservadas en frío en una nevera para su transportación al laboratorio. Una vez allí, las muestras fueron congeladas (-20 °C) hasta su procesamiento.
Caracterización de la dieta y evaluación de la abundancia de las presas
A las aves sacrificadas se les extrajo el sistema digestivo en el laboratorio con ayuda de pinzas y tijeras. Para facilitar la clasificación e identificación de las presas en las muestras de sedimento, las mismas fueron filtradas por un tamiz de 500 μm.
Los invertebrados presentes, tanto en el contenido estomacal como en las muestras de sedimento, fueron identificados y cuantificados con la ayuda de un estéreo Leica wild M8 50X. En la mayoría de los casos, la identificación de los invertebrados se realizó a partir de los restos encontrados en las muestras (e.j., mandíbulas, patas, cabeza, conchas) y para ello se utilizaron guías de identificación de invertebrados acuáticos (Higgins y Thiel 1988, Bouchard 2004). En otros casos se encontraron invertebrados enteros lo que hizo más fácil la identificación. El nivel taxonómico al cual se llegó durante la identificación varió según el grupo. A nivel de Phylum quedaron los nemátodos (Nematoda), a nivel de Clase los poliquetos (Polychaeta) y los moluscos (Gasteropoda), a nivel de Orden los foraminíferos (Foraminiferida), los insectos acuáticos (Hemiptera, Diptera y Plecoptera) y los crustáceos (Tanaidacea y Ostracoda), y a nivel de Familia los ácaros (Halacaoroidea) y los sipun-cúlidos (Sipunculidae). Los restos de invertebrados que no pudieron ser identificados bajo una categoría taxonómica fueron agrupados en la categoría Otros.
Análisis de los datos
Caracterización de la dieta y evaluación de la abundancia de las presas
La caracterización de la dieta se basó en la determinación de la abundancia proporcional y frecuencia de aparición de cada presa identificada en los estómagos de las aves. La abundancia proporcional representa la proporción de cada ítem dentro de la muestra total de ítems identificados, mientras que la frecuencia de aparición se refiere a la proporción de estómagos donde se encontró cada uno.
Además, se calculó la media y error estándar de la abundancia de presas registradas (ind./muestra). Para verificar la contribución de cada grupo de presas a la comunidad de invertebrados disponibles en ambos humedales se determinó la abundancia proporcional de cada uno de ellos. La abundancia de invertebrados no cumplió con las premisas de las pruebas estadísticas paramétricas (Normalidad; prueba Kolmogorov-Smirnov y homogeneidad de varianza; prueba Levene), por ello la comparación entre localidades se realizó mediante la prueba no paramétrica U de Mann-Whitney.
Relación entre la dieta y las presas disponibles
Se calculó el Índice de Similitud Proporcional (ISP) (Hurlbert 1978 eq. 1) para establecer similitudes entre la dieta de las tres especies de Calidris y las presas disponibles en el hábitat de forrajeo. El ISP varía entre cero y uno. Los valores cercanos o iguales a uno del ISP indican muestras que comparten un gran número de artículos. Autores como Feinsinger et al. (1981) avalan la utilización de este índice como una medida de amplitud de nicho.
Para verificar si las especies de Calidris realizaron selección positiva, negativa o ausencia de selección de las presas disponibles en el sedimento, se calculó el índice de Ivlev (1961). Cuando los valores se encontraron entre -0,6 y 0,6 se asumió que las presas fueron consumidas en correspondencia con su disponibilidad en el hábitat. En tanto, los valores mayores de 0,6 se consideraron selección positiva y los menores de -0,6 selección negativa.
Todos los resultados de las pruebas estadísticas se consideraron significativos con un a < 0,05. Los análisis se realizaron con el programa Statistica 8.0 (StatSoft Inc 2008)
RESULTADOS
Caracterización de la dieta de las tres especies de Calidris
En los estómagos de las aves recolectadas se identificaron un total de once tipos de presas (huevos, larvas y pupas de dípteros, hemípteros acuáticos, ostrácodos, nemátodos, arácnidos acuáticos, foraminíferos, gasterópodos, semillas y otros). El tipo de presa, la frecuencia con que fueron consumidas, así como su abundancia proporcional, variaron por especie y localidad (Fig. 2). En forma general, los dípteros (huevos, larvas y pupas) fueron las presas más frecuentes y abundantes en la dieta de C. minutilla, C. pusilla y C. mauri. La mayoría de los recursos tróficos estuvieron presentes en la dieta de al menos dos de las especies de aves limícolas. La excepción estuvo con los nemátodos y arácnidos acuáticos solo presentes en los contenidos estomacales de C. minutilla y los foramíniferos encontrados en la dieta de C. mauri. Calidris minutilla consumió nueve tipos de presas, cuatro en Río Máximo y siete en Tunas de Zaza, pero su dieta en ambas localidades estuvo dominada por los huevos, larvas y pupas de dípteros (Fig. 2). El resto de los ítems encontrados estuvo pobremente representado.
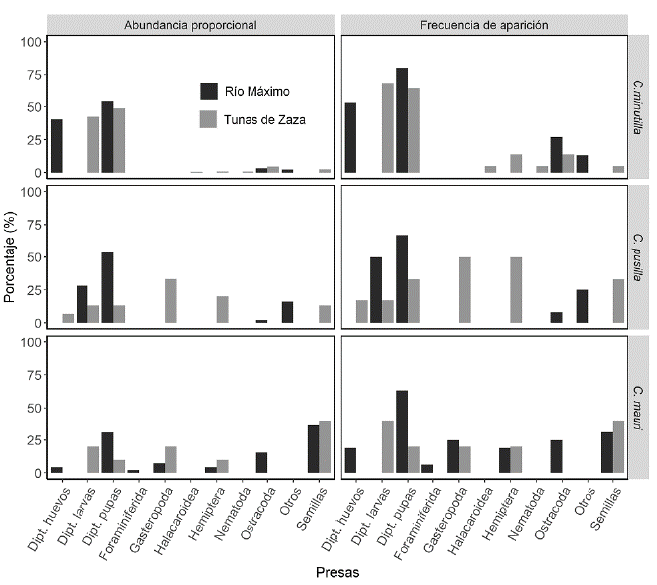
Figura 2 Abundancia proporcional y frecuencia de aparición de los recursos tróficos identificados en los estómagos de C. minutilla, C. pusilla y C. mauri en los humedales de Río Máximo y Tunas de Zaza.
La dieta de C. pusilla estuvo compuesta por ocho tipos de presas, cuatro en Río Máximo y seis en Tunas de Zaza (Fig. 2). En Río Máximo, las larvas y pupas de dípteros constituyeron el recurso más frecuente y abundante en los contenidos estomacales de la especie. Sin embargo, en los individuos recolectados en Tunas de Zaza se vio mayor presencia y abundancia de gasterópodos y hemípteros acuáticos.
El contenido estomacal de C. mauri reveló la presencia de ocho tipos de presas (Fig. 2). Los individuos recolectados en Río Máximo consumieron un mayor número de ítems que aquellos recolectados en Tunas de Zaza (siete y cinco, respectivamente). Las semillas fueron el recurso más abundante en la dieta de la especie en ambas localidades. Sin embargo, las pupas y larvas de dípteros fueron las presas más frecuentemente consumidas en Río Máximo y Tunas de Zaza, respectivamente.
Presas potencialmente disponibles para las tres especies de Calidris
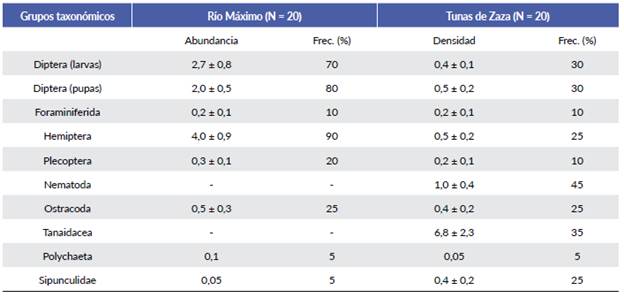
La abundancia de presas potencialmente disponibles no difirió entre localidades (Río Máximo: 9,9 ± 1,6 ind./muestra, Tunas de Zaza: 10,2 ± 2,4 ind./muestra; prueba U de Mann-Whitney U = 170,0; P = 0,770). Las presas disponibles estuvieron integradas por ocho grupos taxonómicos en Río Máximo y diez en Tunas de Zaza (Tabla 1). De ellos, ocho (72,7 %) fueron compartidos entre sitios y dos (Nematoda y Tanaidacea) resultaron exclusivos de Tunas de Zaza. En ambas localidades el arreglo de presas disponibles estuvo caracterizado por la marcada dominancia de solo dos taxones. Los hemípteros y dípteros fueron las presas predominantes en Río Máximo, mientras que en Tunas de Zaza las más frecuentes y abundantes fueron los tanaidáceos y los nemátodos (Tabla 1).
Relación entre la dieta de las tres especies de Calidris y las presas disponibles
El Índice de Similitud Proporcional entre la dieta y las presas disponibles no varió entre especies, pero sí entre localidades (Fig. 3). En Río Máximo la dieta de las tres especies guardó relación con la disponibilidad de presas en el sedimento, pero en Tunas de Zaza la similitud entre dieta y disponibilidad de presas fue baja. Consecuentemente, el análisis de selección reveló que en Río Máximo las aves limícolas estudiadas consumieron presas de acuerdo con su disponibilidad en el sedimento (Fig. 4). La excepción estuvo dada por C. mauri quien seleccionó positivamente a los ostrácodos y negativamente a los hemípteros (índice de Ivlev de 0,68 y -0,69, respectivamente). A diferencia de Río Máximo, en Tunas de Zaza las tres especies de aves mostraron un mayor grado de selectividad trófica (Fig. 4). Las larvas y pupas de dípteros fueron seleccionadas de forma positiva por las tres especies (índice de Ivlev de 0,70 - 0,85). En tanto, los hemípteros fueron seleccionados positivamente por C. pusilla y C. mauri, y negativamente por C. minutilla. Esta última especie también realizó selección negativa de Nematoda.
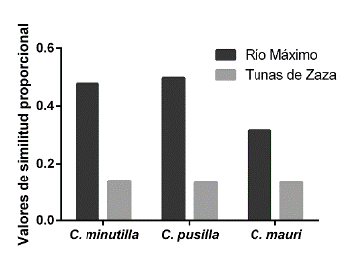
Figura 3 Valores del Indice de Similitud Proporcional para las tres especies de Calidris en Río Máximo y Tunas de Zaza.
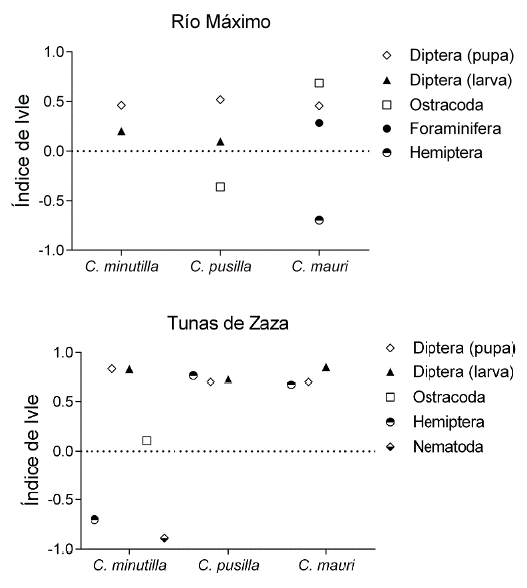
Figura 4 Valores del índice de selección de Ivlev para las presas consumidas por C. minutilla, C. pusilla y C. mauri en Río Máximo y Tunas de Zaza. Por encima de 0,6 se consideró selección positiva y por de debajo de -0,6 se consideró negativa. Los valores intermedios fueron indicativos del consumo de artículos en correspondencia con su disponibilidad en el hábitat.
DISCUSIÓN
Los resultados del estudio revelaron que en los humedales naturales estudiados los bandos mixtos de C. minutilla, C. pusilla y C. mauri pueden flexibilizar su respuesta trófica en relación con la disponibilidad de presas en el sedimento, lo cual es consistente con lo que se ha descrito en la literatura (Baker 1977, Andrei et al. 2009). Dado que estas aves viajan grandes distancias entre sus áreas de reproducción e invernada, esta conducta de forrajeo oportunista constituye una ventaja adaptativa que les permite incrementar las reservas de grasa rápidamente y completar de modo satisfactorio su migración (Davis y Smith 2001).
Las aves estudiadas hicieron uso de los diferentes estadios de los dípteros que habitan en el sedimento de ambos humedales. La importancia de los primeros estadios (huevos, larva y pupa) estuvo acentuada por el comportamiento selectivo hacia estas presas en Tunas de Zaza donde estos invertebrados no fueron abundantes. Los dípteros, en particular aquellos de la familia Chironomidae, se encuentran ampliamente representados en los lodazales intermareales (Roth y Brown 1980, Cheng y Frank 1993). Este grupo de insectos presenta un amplio espectro de tolerancia a variables abióticas como la salinidad (intervalo: 4-50 %o) y temperatura (intervalo: 10-40 °C) lo que le garantiza su supervivencia en sistemas altamente variables como los lodazales intermareales (Oliver 1971, Neumann 1976). Los valores promedio de temperatura y salinidad registrados en Río Máximo y Tunas de Zaza se encontraron dentro de estos intervalos descritos. Un importante elemento que favorece el marcado uso de los primeros estadios de dípteros es su distribución en los estratos superiores del sedimento en gran parte de su ciclo de vida (Kranzfelder et al. 2015), lo cual los hace más accesibles para las aves de orilla.
A diferencia de lo encontrado en Río Máximo, el Índice de Similitud Proporcional mostró bajos valores para las tres especies en Tunas de Zaza. Este resultado estuvo principalmente relacionado con la ausencia de tanaidáceos dentro de la dieta de los Calidris. Por la literatura se conoce que C. pusilla y C. mauri consumen estos crustáceos (Skagen y Oman 1996, Wolf 2001, Mathot et al. 2010), y debido a su similitud en morfología y comportamiento con otros invertebrados (i.e., anfípodos), podría esperarse que C. minutilla también haga uso de ellos (Gratto et al. 1984, Colwell y Landrum 1993). La ausencia de tanaidáceos en los estómagos de las tres especies de Calidris contradice la esperada conducta oportunista de las aves limícolas en Tunas de Zaza. Este resultado podría relacionarse con la distribución de los tanaidáceos dentro del lodazal inter-mareal donde se alimentan los Calidris en Tunas de Zaza y el riesgo a la depredación que tienen las aves mientras forrajean. La abundancia promedio de estos macroinvertebrados estuvo por encima de los seis ind./muestra, en tanto el valor de frecuencia de aparición estuvo por debajo de 40 %. Al ubicar dentro del área de alimentación las unidades de muestreo donde se registraron los tanaidáceos, se constató que éstas se localizaron en los últimos 150 m de las líneas trazadas y por tanto más próximas a la vegetación. La aves limícolas podrían tener un mayor riesgo a la depredación por Falco peregrinus (Tunstall, 1771) y Falco columbarius (Linnaeus, 1758) si utilizan estos mi-crohábitats más cercanos a la vegetación. Por lo general, las rapaces suelen camuflarse dentro de los parches de vegetación que existen en los lodazales intermareales para así acechar a sus presas y realizar ataques sorpresivos. Si se tiene en cuenta el balance que realizan las aves limícolas entre el peligro de ser depredado y la tasa de ingestión de las presas (Dekker y Ydenberg 2004, Pomeroy 2006), se podría especular que evitaron alimentarse en los parches de mayor peligro a la depredación y por consiguiente dejaron de consumir a los tanaidáceos.
Otro elemento de interés fue la elevada presencia y abundancia de semillas en los estómagos de C. mauri, tanto en Río Máximo como en Tunas de Zaza. Las semillas también fueron documentadas en los contenidos estomacales de C. minutilla y C. pusilla, pero solo para la localidad de Tunas de Zaza. La importancia del uso de semillas por aves limí-colas ha estado pobremente documentada en los estudios de dieta, pero se conoce que en algunas especies como Calidris fuscicollis Vieillot, 1819 puede constituir el 100 % del contenido estomacal aun utilizando un humedal natural (Montalti et al. 2003). De igual modo, ha sido documentada ocasionalmente en aves limícolas que se alimentan en los arrozales durante el periodo migratorio (Acosta et al. 1994, Cifuentes-Sarmiento y Renjifo 2016). Sin dudas, el hallazgo de semillas en la dieta de las tres especies de Calidris fue un resultado no esperado dada su ausencia en las muestras de sedimento. Este hecho permite hipotetizar el uso de hábitats alternativos para completar sus elevadas demandas metabólicas durante el periodo migratorio otoñal. Ambas localidades estudiadas tienen ecosistemas de arroceras en la cercanía (Aguilar 2010) los cuales podrían tener un importante papel al proveer hábitats de alimentación durante el periodo de marea alta cuando los hábitats costeros dejan de estar disponibles para estas aves (Blanco 2006). Futuros estudios deberían realizarse para esclarecer la conexión entre ambos ecosistemas.
Este trabajo constituye la primera investigación que ofrece elementos sobre la dieta de Calidris en humedales naturales cubanos. Los ítems identificados dentro de la dieta de las tres especies fueron variados y guardaron correspondencia con las presas disponibles en el sedimento de los lodazales intermareales de ambas localidades. La única excepción fue la presencia de semillas, fundamentalmente en Tunas de Zaza lo que evidencia la utilización de hábitats alternativos durante el forrajeo. De igual modo, presas altamente abundantes en el sedimento como los tanaidáceos en Tunas de Zaza no se registraron en la dieta lo que influyó en el bajo valor del Índice de Similitud Proporcional. La amplia variedad de ítems encontrada indica la flexibilidad en la dieta de las tres especies dentro los humedales cubanos. Esto constituye un aspecto favorable para la conservación de estas aves de orilla durante su estancia en la isla. Incluso, el grupo de insectos más importante dentro de la dieta (dípteros) es altamente tolerante a variables abióticas por lo que la variabilidad natural de los lodazales intermareales de Río Máximo y Tunas de Zaza no afectará la disponibilidad de estos recursos y por tanto la supervivencia de las aves. De igual modo, desde el punto de vista de la conservación de las aves limícolas en los humedales cubanos, es de vital importancia dilucidar la conexión con ecosistemas de arroceras y evaluar la magnitud en que son utilizados y si pueden tener alguna categoría de protección.

