




![ETIOLOGÍA DE LA MUERTE DESCENDENTE DEL DURAZNERO [Prunus persica (L.) BATSCH] EN LOS MUNICIPIOS DE MANIZALES Y VILLAMARÍA, DEPARTAMENTO DE CALDAS](/img/en/next.gif)





Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
Print version ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.35 no.134 Bogotá Jan./Mar. 2011
ENTOMOLOGÍA
1 Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. Correo electrónico linacamp@gmail.com
2 Museo de Historia Natural de Paris. Correo electronico: jimenoptera@gmail.com
3 Profesor Asociado, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Carrera 30 No. 45 â 03 Edificio 425 oficina 207, Bogotá D.C., Colombia. Correo electrónico: mgandradec@unal.edu.co
Resumen
Se registraron 51 especies de mariposas, pertenecientes a seis familias y 17 subfamilias en las ciénagas de Córdoba y zonas adyacentes. Se registraron por primera vez para el Caribe colombiano las especies Pseudolycaena marsyas, Zaretis itis y Phocides polybius polybius. El complejo de humedales del río San Jorge presentó un mayor número de especies frente al complejo del Sinú (Lorica y Martinica). Se elaboró una lista de 211 especies que incluye las especies encontradas en Córdoba y en las ciénagas del Cesar: Zapatosa (98), Mata de Palma—la Pachita (66) y la Jagua de Ibirico (85). Se destaca que las ciénagas de Córdoba mostraron altos patrones de afinidad (50% de especies compartidas), mientras que en la llanura del Caribe (Jagua de Ibirico), encontramos valores cercanos al máximo de valor de disimilitud, uno.
Palabras clave: mariposas, humedales, biodiversidad, conservación, Córdoba, Colombia.
Abstract
51 species of butterflies, belonging to six families and 17 subfamilies, were found in Córdoba's wetlands and adjacent areas. The species Pseudolycaena marsyas, Zaretis itis y Phocides polybius polybius are registered for the first time in the Caribbean area. The wetland complex of the San Jorge river has a higher number of species in comparison with the Sinu complex (Lorica and Martinica).A list of 211 species was elaborated including the species found in Córdoba and the species of Cesar's wetlands: Zapatosa (98) and Mata de Palma, the Pachita (66) and La Jagua de Ibirico (85). We emphasize that the Cordoba's wetlands show high afinity patterns (50% of shared species), while in the Caribbean swamps (Jagua de Iribico), we found values approaching the maximum value of dissimilarity.
Key words: butterflies, wetlands, biodiversity, conservation, Córdoba, Colombia.
Introducción
El complejo de ciénagas de Córdoba, en las cuencas del río San Jorge y Sinú, abarca una gran extensión de espejos de agua (110.000 hectáreas). Las ciénagas de Ayapel (40.000 hectáreas), ciénaga Grande de Lorica (38.000 hectáreas), Betancí (3.250 hectáreas), Martinica (2.000 hectáreas), El Arcial (2.000 hectáreas), El Porro (1.500 hectáreas) y Cintura (1.000 hectáreas), contribuyen al mantenimiento de diferentes ecosistemas y tipos de vegetación que dependen directa o indirectamente de estos ambientes acuáticos. (Cárdenas-Arévalo et al. 2010). Adicionalmente constituyen la única fuente de agua potable para los habitantes de la región.
Las diferentes actividades antrópicas ligadas a los complejos de humedales: pesca artesanal, Cultivos agrícolas semestrales (arroz Oryza sativa) en porciones de terrenos seminundables,desecación de terrenos para uso en la ganadería extensiva, tala de las áreas circundantes, entre otros; han causado cambios drásticos en la dinámica del agua y en la función biológica que cumplen estos ambientes acuáticos en las áreas del valle del río Sinú, San Jorge y sus llanuras inundables. Estos procesos han degradado y alterado la composición vegetal y en consecuencia, alterado la estructura de la comunidad faunística, entre ellas las mariposas pertenecientes a estas dos grandes superfamilias (Hesperioidea y Papilionoidea), son uno de los grupos que mejor pueden reflejar estos cambios, así mismo responder a las características ambientales y su adaptación a los hábitat disponibles (Fagua, 1996; Fagua et al. 1999). Es por esto que articular trabajos de vegetación y de grupos faunísticos como las mariposas y otras especies capaces de reflejar el estado de conservación de los ecosistemas, se convierte en una herramienta importante en el establecimiento de planes de manejo y conservación de hábitats (Coddington et al. 1991).
En los trópicos de América la diversidad de lepidópteros alcanza 180.000 especies, de las cuales 17.950 son mariposas diurnas (Andrade-C., 2007). Para Colombia, se registran 3.274 de las cuales 350 son endémicas.En el presente estudio se obtuvieron datos importantes sobre las especies de mariposas de las ciénagas de Córdoba y sus áreas de influencia, ya que estas aportan un número significativo de especies a la fauna de mariposas del Caribe Colombiano y que, hasta la fecha no se tenían registros que ampliaran los rangos de distribución para Colombia de las especies presentes. Se incluyen además, datos sobre la diversidad al interior de las ciénagas que conforman este complejo, y una comparación de la composición y riqueza del complejo de ciénagas de Córdoba con las ciénagas del Cesar y áreas de las llanuras en el Caribe como La Jagua de Ibirico en el Cesar, obtenidos en estudios precedentes.
Métodos
Área de estudio
El trabajo se desarrolló en el nor-oriente del departamento de Córdoba, en los alrededores del complejo de humedales del valle inundable del río San Jorge, en altitudes comprendidas entre los 40 y 100 metros. Se efectuaron salidas de campo a varias localidades de la ciénaga de Ayapel en el municipio del mismo nombre, y las ciénagas de Arcial, El Porro y Cintura en el municipio de Pueblo Nuevo, estas tres últimas ciénagas se tomaron como una unidad, por su cercanía e interconexión. Otra parte del estudio incluyó el complejo de ciénagas y llanuras de inundación que hacen parte del valle del río Sinú, este sistema lagunar presenta dos áreas importantes: Betanci y Montería, ubicadas en el municipio de Montería y la ciénaga Grande de Lorica en el municipio del mismo nombre.En la ciénaga de Ayapel, las colectas se realizaron en cinco localidades, La Balsa, La Ceja, La Cucharita, La Escobilla y Playa Blanca, entre los meses de abril y marzo del 2004. En las ciénagas de Arcial, El Porro y Cintura las localidades estudiadas fueron ocho, hacienda Praga, hacienda Toronto, vereda La Coroza en la hacienda La Baqueta, inmediaciones del corregimiento Arcial, sector Playón de Cintura, corregimiento Café Pisaó en la hacienda La María, corregimiento el Porro, en la hacienda La Embajada y en la parcela de La Aranza, en sector anexo a la ciénaga el Porro, en el mes de julio del mismo año.En las ciénagas del valle del río Sinú los muestreos se realizaron en el sector La Isla, Abisnia (ITA), Cubinca (ciénaga de Lorica), sitio el Tiestico (ciénaga de Lorica), sitio La Peinada (ciénaga de Lorica), Corregimiento el Sillón (ciénaga de Lorica), San Sebastián (ciénaga de Lorica),vereda de Martinica (Ciénaga de Martinica), entrada al caserío de Martinica, hacienda Diluvio (Ciénaga de Martinica), corregimiento Betanci, en la margen de la ciénaga, margen izquierda del poblado y en las inmediaciones Escuela Mixta Maracayo, en los meses de julio-noviembre y diciembre de 2004.
Descripción de los ambientes adyacentes de las ciénagas
La vegetación que circunda los complejos de las ciénagas pertenecen a Bosque Seco Tropical, donde se puede encontrar vegetación de tipo arbórea y arbustiva, dispersa o formando pequeños parches con diferentes grados de conservación por lo cual es posible encontrar desde áreas bien conservadas hasta sectores con relictos de formaciones originales (Rivera-Díaz, 2010). En las localidades pertenecientes al complejo de humedales del río San Jorge, son predominantes las intervenciones de tipo antrópico, que corresponden a las actividades de entresaca, tala y pastoreo; esta disposición del uso del suelo evidencia la ausencia de coberturas vegetales que incentiven la regeneración natural del bosque (Rangel et al. 2010). En general el complejo de ciénagas de San Jorge (Arcial-El porro y Cintura) presenta aún fragmentos grandes de bosques (Cárdenas-Arévalo et al. 2010). En contraste el complejo cenagoso del bajo Sinú, presenta una vegetación típica de bosque seco tropical, caracterizado por áreas destinadas a las prácticas agroforestales y pecuarias que han dado origen a áreas más homogéneas dominadas por potreros y pequeños relictos de bosque muy intervenidos que parecen comportarse como ecotonos por su tamaño (Cárdenas-Arévalo et al. 2010).
Diseño de colecta
Para cada una de estas localidades se dedicó un día de colecta, en las cuales se recorrieron transectos lineales de 100 x 2 m en zonas de bosque y borde de bosque; se usó la técnica de captura con jama entomológica y trampasVan Someren Rydoncon cebo (pescado descompuesto) que se instalaban en la mañana y se recogían en horas de la tarde. La totalidad del esfuerzo de muestreo se realizó durante las horas del día (09:00-16:00)realizando la mayor cantidad de recorridos posible. Para cada ciénaga el esfuerzo de muestreo fue de siete horas/hombre/día, abarcando homogéneamente cada unidad de vegetación. Los muestreos incluyeron diferentes unidades vegetales como: bosques con diferentes estados de intervención y conservación (bosque seco tropical), áreas abiertas, pastizales, cercas vivas, potreros, rastrojos y bosques inundables. Los ejemplares fueron sacrificados por presión digital en el tórax y depositados en sobres de papel milano blanco con los datos de colecta (localidad, sitio, fecha, colector, altitud), se anotaron datos de observaciones de las plantas hospederas, las horas de actividad y zonas donde fueron encontradas; una vez los ejemplares llegaron al laboratorio, se procedió a realizar su extendido, montaje, etiquetado, catalogación y determinación por medio de claves e ilustraciones, siguiendo a Adams(1985), Le Crom et al. (2004) y DeVries(1987,1988). Se utilizó la revisión taxonómica editada por Lamas et al. (2004) y la referencia de la colección del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, Bogotá (ICN-MHN-L) para la identificación taxonómica.
Análisis de datos
Los datos de riqueza se generaron a partir del número total de especies presentes para cada localidad de estudio. Los datos de abundancia fueron tomados como el número de individuos para cada especie, (Anexo1). Se determino la riqueza específica con el índice de Margalef que presupone una relación funcional entre el número de especies y el número total de individuos, y se obtuvo la equitatividad por medio del índice de Shannon-Wiener que expresa la uniformidad de los valores de importancia a través de todas las especies de la muestra (Magurran, 1988).En el análisis comparativo de cada una de las ciénagas construida a partir de los datos de diversidad (Shannon-Wiener), se utilizó la grafica de cajas y bigotes que representa la distribución estadística unidimensional que refleja cinco parámetros: límite inferior, primer cuartil, mediana, tercer cuartil y límite superior; siendo una medida de la simetría o asimetría de la distribución, del sesgo y de la dispersión de los datos (Sokal & Rolf, 1985).Mediante un análisis de varianza se contrastaron los resultados de estos índices de diversidad de especies dentro de las ciénagas, con el fin de comparar las medias de las diversidades y verificar si las diferencias numéricas son significativas (Magurran, 1988). Para medir la diversidad Beta, se realizó un análisis de similitud y complementariedad de la composición de especies entre el complejo de ciénagas de Córdoba, a partir de la riqueza total y las especies comunes entre las ciénagas comparadas, empleando los datos de presencia/ausencia para hacer las combinaciones en pares de biotas (Cowell & Coddington, 1994), en este caso mientras más disímiles son las localidades comparadas el valor se acerca más a uno (Moreno, 2001). Adicionalmente, se realizó una comparación nominal y porcentual de la lista de las especies de mariposas registradas en este trabajo frente a las listas de especies reportadas en otros estudios de las ciénagas del Caribe colombiano, tal como en la ciénaga de Zapatosa, municipio de Chimichagua, departamento de Cesar (Pulido-B., 2008), ciénagas de Mata de Palma y La Pachita, municipio El Paso, departamento del Cesar (Pulido-B., 2008) al igual que las especies del municipio de la Jagua de Ibirico, en el departamento del Cesar (Pulido-B., 2008), presentes en áreas cenagosas o en áreas planas del Caribe. Para esto se tomaron como una localidad los sectores del valle inundable del río San Jorge (ciénaga de Ayapel y las ciénagas de Arcial, El Porro y Cintura), y como otra localidad las que hacen parte del valle del río Sinú (ciénagas de Lorica, Martinica y Betanci).
Resultados
Para el complejo de ciénagas de Córdoba se encontraron 215 individuos pertenecientes a 51 especies, 17 subfamilias y seis familias; Hesperidae (dos subfamilias, seis géneros), Papilionidae (una subfamilia, dos géneros), Pieridae (dos subfamilas,seis géneros), Riodinidae (una subfmalia,una género), Lycaenidae (dos subfamilias, tres géneros) y Nymphalidae (nueve subfamilias, 21 géneros) (Anexo 1). Figura 1. Riqueza de especies para cada unas de las familias y subfamilias presentes en el complejo de ciénagas de Córdoba.
La subfamilia Heliconiinae es la que presentó el mayor número de especies (ocho) y géneros (seis), con igual número de especies Biblidinae con ocho y cinco géneros, Nymphalinae y Danainae con igual número de especies (tres) y con dos y un géneros respectivamente; las subfamlias Charaxinae, Ithomiinae y Limenitidinae solo estuvieron representadas por una sola especies en cada caso (Figura 1, Anexo 1). Las ciénagas de Ayapel y Betanci presentaron la mayor riqueza de especies con igual número 23, seguida por la ciénaga grande de Lorica con 22, las ciénagas de Arcial, El Porro y Cintura con 21, mientras que la ciénaga de Martinica presenta la menor riqueza, 17 especies (Anexo 1, Figura 2).
El sistema de humedales del río San Jorge: Arcial. El Porro y Cintura presentó 21 especies pertenecientes a 11 subfamilias, donde Heliconiinae fue la subfamilia más rica (Figura 2). En la subfamilia Charaxinae las especies Zarethis itis, y en Biblidinaela especie Callicore pitheas fueron los dos únicos registros para las ciénagas de Córdoba.
En cuanto a la ciénaga de Ayapel, que también hace parte de este valle, encontramos que Coliadinae registró el mayor número de especies, seguida por Heliconiinae, Pyrginae y Biblidinae (Figura 2). Hesperiinae con Polites vibex y Limenitidinae con Adelpha iphiclus, fueron los únicos registros obtenidos en las ciénagas de Córdoba.
Se estableció en forma general que las ciénagas influenciadas por el río San Jorge presentaron un mayor número de especies frente a las del complejo del Sinú (Lorica y Martinica), sin embargo la ciénaga grande de Lorica, aumenta los valores de riqueza a partir de los registros de Nymphalinae con especies como Anartia jatrophae. Aunque Betanci iguala en número de especies a las presentadas por Ayapel (San Jorge) debido en gran medida a especies de Heliconiinae como Heliconius melpomene, Dryas iulia y Agraulis vanillae.
Los análisis de diversidad, realizados a partir de los datos de riqueza y número de individuos para cada una de las ciénagas (Tabla 1), muestran que no existen diferencias estadísticamente significativas entre la diversidad de las ciénagas (índice de Shannon-Wiener: F (4,101; 0,05) = 1.31; p = 0,27).
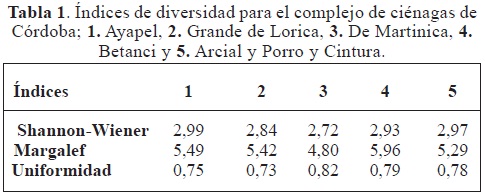
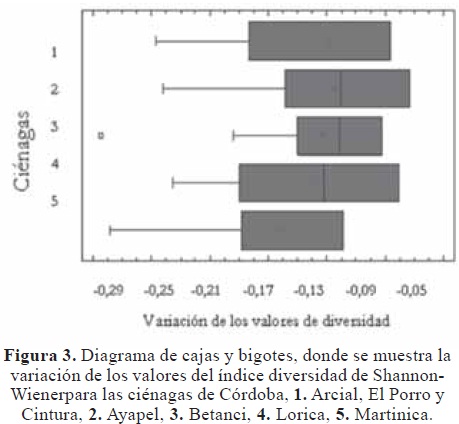
Los valores de diversidad por ciénagas no presentaron diferencia estadística, debido a que la dispersión de los datos claramente se sobrepone entre sí (Figura 3). La media de la diversidad para cada sitio de muestreo en cuanto a la composición de especies, expresan un patrón de uniformidad similar (Tabla 1).
De acuerdo con los valores de complementariedad y las especies encontradas en los diferentes sitios, se estableció una baja afinidad en la similitud y composición de los ensambles de especies; además se encontró que entre Lorica- Arcial, El Porro y Cintura se presentó la mayor disimilitud, seguida por Martinica-Arcial, El Porro y Cintura y Ayapel- Arcial, El Porro y Cintura, con igual proporción de especies compartidas; en cuanto a Lorica y Betanci son los que comparten la mayor proporción de especies (Tabla 2).
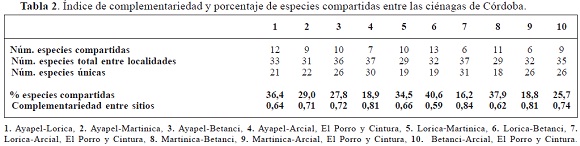
Entre Lorica-Arcial, El Porro y Cintura, y entre Ayapel - Arcial, El Porro y Cintura, se presentaron el más alto número de especies únicas 31 y 30 respectivamente, en cuanto a Lorica-Martinica y Lorica-Betanci, presentaron los más bajos valores de especies únicas, 19 cada una de ellas, seguidas por Martinica-Betanci con 18 especies compartidas (Tabla 2).
En Ayapel, se registró como especies únicas individuos de Polites vibex, Phocides polibius polybius, Heliconius erato, Caligo illioneus oberon y Adelpha iphiclus iphiclus, siendo exclusivas para ésta ciénaga, mientras que Chioides catillus, Leptotes cassius, y Nica flavilla se presentaron como especies únicas para Betanci (Anexo 1). Arawacus oetolus y Anartia amathea solo se presentaron en Lorica; mientras que Arcial, El Porro y Cintura presentaron el mayor número de especies únicas, entre las que se encuentran Morpho helenor peleides, Marpesia alcibiades, Zarethis itis, Battus polydamas y Theope virgilius (único ejemplar de la familia Riodinidae colectado en el estudio); entre tanto Martinica registró el menor número de especies (17), de las cuales Danaus plexippus y Heraclides thoas fueron especies únicas para la localidad.
Al contrastar las especies halladas en las ciénagas de Córdoba con otras regiones del Caribe, se encuentra un total de 211 especies, asociadas a áreas de ciénagas y llanuras del Caribe, el mayor número de especies la registra Zapatosa (98), seguido por La Jagua de Ibirico (85), y Mata e Palma (Cesar) (66), mientras que las ciénagas de Córdoba presentan bajos valores de riqueza (Anexo 2).
En general, entre las ciénagas muestran patrones de afinidad, mientras que se marcó una gran diferencia con la llanura del Caribe (Jagua de Ibirico); sin embargo, se resalta las diferencias de composición de especies entre las ciénagas de Córdoba y Cesar, el caso de Lorica-Martinica y Betanci con La Jagua de Ibirico con tan solo el 4,3% de especies compartidas, incluso presentando esta última el mayor número de especies únicas (110) (Tabla 3).
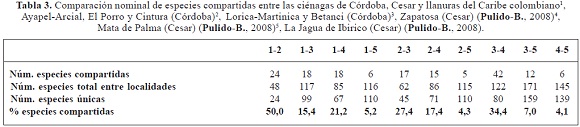
La composición de especies entre las ciénagas de Córdoba son muy afines con el 50% de especies compartidas, estas dos áreas tienen la mayor similitud de especies, mientras que al compararlas con las especies presentes en la llanura de Caribe (La Jagua de Ibirico), se presentan valores que casi alcanzan el máximo valor de disimilitud (uno) (Tabla 3).
Especies como Dryadula phaetusa, Euptoieta hegesia, Heliconius erato hydara, Adelpha iphiclus iphiclus, Ascia monuste monuste y Theope virgilius se presentan únicamente en áreas de ciénagas, sin que se encuentren para otros sitios de la llanura del Caribe, también sobresalen Pseudolycaena marsyas, Phocides polybius polybius, Marpesia alcibiades y Caligo illioneus oberon registrados para las ciénagas del valle del río San Jorge.
Se resaltan la captura de Mechanitis polymnia único Ithomiinae que se registró en áreas aledañas a las ciénagas de Córdoba, sin que se haya sido registrado aun en las ciénagas del Cesar y llanuras del Caribe (Anexo 2). Para las ciénagas del valle del río Sinú, se capturaron Nica flavilla, Heraclides anchisiades, como especie única, así como Pyrisitia proterpia y Hesperocharis nera, únicas para éstas ciénagas (Figura 4 y Anexo 2).
Heliconiinae aparece como la subfamilia con mayor número de especies para las ciénagas de Córdoba por encima de las ciénagas de Zapatosa y Mata de Palma en el Cesar, mientras que Biblidinae aparece como la segunda subfamilia con mayor número de especies con respecto a Mata de Palma ya que Zapatosa registró un número más alto de especies en esta subfamilia (Figura 4). De igual forma Mata e Palma superan la riqueza de especies de Ayapel en Córdoba.
Discusión
En las ciénagas de Cordoba, se encontraron representadas las diferentes familias de mariposas tropicales. La familia Nymphalidae aportó el mayor número de géneros y especies, esto concuerda con la mayoría de los estudios realizados en Colombia ya que ésta familia presenta el mayor número de subfamilias y el mayor número de especies, por esta razón, las comparaciones a nivel de familias no aporta información diferencial. Sin embargo al interior del complejo de ciénagas influenciadas por el río San Jorge y el río Sinú se evidencian diferencias en la composición y la riqueza de especies.
La ciénaga de Ayapel registró el mayor número de especies, esto puede deberse en gran parte a una matriz mixta de vegetación. Se encontraron áreas bien demarcadas, fragmentos de bosque poco intervenidas como el bosque La Cucharita y áreas abiertas muy degradadas como el sector de Playa Blanca. En este sector se encontraron especies como Polites vibex, Phocides polibius polybius que prefieren las áreas abiertas. En las áreas de bosque La Cucharita se hallaron dos ejemplares de Pseudolycaena marsyas (macho y hembra), asociadas a áreas con abundante vegetación y presencia de claros, donde puede encontrar alimento fácilmente, este Lycaenidae es una especie de difícil captura, debido a aspectos ecológicos como la estacionalidad y preferencias por espacios abiertos sobre la copas de los árboles (De la Maza, 1987). Otras especies que aparecen como exclusivas para la ciénaga de Ayapel son Heliconius erato, especie típica de áreas intervenidas y abiertas (Andrade-C. et al. 2007A), Caligo illioneus oberon y Adelpha iphiclus iphiclus asociados a bosques de ribera que aumentan la riqueza para esta ciénaga. El incremento en la heterogeneidad del hábitat y el acceso a fuentes de néctar en los márgenes del bosque o parches de vegetación en zonas despejadas facilitaron registro de especies de Theclinae y Limenitidinae, que normalmente no abandonarían el dosel (Raguso & Llorente, 1997), así mismo de mariposas "saltarinas" pertenecientes a Pyrginae y Hesperiinae que poseen cuerpo robusto y alas pequeñas que les otorgan un vuelo rápido, por lo que requieren de áreas abiertas e iluminadas al igual que una alta oferta de flores para desarrollar sus actividades (Vélez, 1989), estas se encuentran asociadas a sitios donde les es posible conseguir lugares de percha para recibir el sol y áreas despejadas para su potente vuelo (Andrade-C. et al. 2007).
En el sistema de humedales del río San Jorge: Arcial. El porro y Cintura, se encontraron especies como:Zarethis itis, Callicore pitheas, Theope virgilius, Marpesia alcibiades y Taygetis laches que además de aparecer como únicas para este complejo, toman importancia debido a que son poco colectadas y sus requerimientos de hábitat se restringen a áreas poco alteradas o al menos a fragmentos de bosque y áreas de borde donde les es fácil acceder a la oferta de alimento. Heliconiinae aparece como la subfamilia más diversa y abundante en las ciénagas de Córdoba, esto en gran medida se puede atribuir a su estrecha relación con plantas de la familia Passifloracea con hábito de crecimiento tipo trepador (hospedera) y cuyo néctar es la fuente de alimentación en estado adulto (Gilbert, 1984; Prieto & Constantino, 1996), varias especies de estas plantas se encontraron en el estrato herbáceo de los relictos de bosque de la Ciénaga de Ayapel, al igual que en la vegetación de bosques de galería presentes en la ciénaga de Lorica, y en parches de vegetación herbácea de áreas anegadas como la presentada por las ciénagas de Arcial, el Porro y Cintura.
Se observó una relación entre las zonas de bosque con un menor grado de intervención y el aumento de la riqueza de especies. Tal es el caso las ciénagas de Arcial, El Porro y Cintura y de algunos sectores en la Cienaga de Ayapel, donde encontramos un gran número de especies exclusivas (30) en comparación con las ciénagas del complejo Sinú.La ciénaga de Betanci en valle del río Sinú, presentó igual riqueza que la ciénaga de Ayapel, sin embargo es una de las ciénagas con el menor número de especies exclusivas (3), lo que sugiere que la disminución del tamaño del bosque disminuye a su vez la diversidad de especies. Un gran número de especies encontradas en esta zona se encuentran igualmente asociadas a hábitats tanto intervenidos como poco alterados, tales como Heliconius melpomene, Hesperocharis nera y Urbanus simplicius, en general las especies allí registradas se encuentran asociadas a áreas abiertas de potreros y clara tolerancia a este tipo de hábitats.
La ciénaga de Martinica es la que presenta el menor número de especies y menor número de especies exclusivas, esto concuerda con el estado de alteración de la zona: diferentes procesos como desecación y tala de la vegetación del área de influencia para convertirlos en potreros, han causado la transformación del hábitat original. Las especies con amplia distribución, tolerancia por habitas alterados y generalistas en el uso del hábitat como Danaus eresimus y Danaus gilippus pudieron ser registradas en esta zona.
Es importante tener en cuenta que a pesar de que en general las áreas con alto grado de intervención presentaron valores significativos de diversidad, las especies presentes allí son en su gran mayoría especies consideradas como generalistas, ya que son capaces de colonizar diferentes habitas y no requieren de condiciones ambientales especificas para su establecimiento, por lo tanto no pueden ser consideradas como indicadores de un buen estado de conservación de un ecosistema. Entre estas encontramos: Anartia amathea, Anartia jatrophae y Phoebis sennae (Campos-Salazar, 2008). En este mismo sentido, especies como Phoebis argante y Ascia monuste son propias de dosel, sin embargo, cuando se reduce la cobertura vegetal también pueden hallarse en áreas abiertas aumentando la riqueza en la ciénaga de Lorica. Tal como lo indica Kremen (1992), el incremento aparente en la riqueza puede ser ocasionado por las especies típicas de hábitats muy intervenidos que se encuentran junto a las especies del bosque.
De otra parte, al comparar los resultados de las ciénagas de Córdoba con las ciénagas del Cesar y llanuras del Caribe, especies como Dryadula phaetusa y Euptoieta hegesia de la familia Heliconiinae, pasan a tomar especial importancia, ya que a pesar de ser especies de amplia distribución en Colombia, no se registran para otras áreas del Caribe como La jagua de Ibirico.
La fauna de mariposas en las ciénagas de Córdoba, se diferencia de la fauna presentada en las ciénagas del Cesar, con valores de hasta 0,85 en complementariedad, un resultado muy alto para ser áreas con similares condiciones biológicas. De igual forma, al comparar la composición de las especies de mariposas con áreas como la Jagua de Ibirico se encontró que son completamente disímiles. Estos resultados permiten sugerir que las diferencias presentadas en la composición de la vegetación es un aspecto valioso a tener en cuenta, para esto es importante recordar que una gran proporción de mariposas se alimentan en estado inmaduro de plantas herbáceas y bejucos (Kremen et al. 1993; Ehrlich & Raven, 1964) y que su reducción en las áreas con fuerte intervención podría condicionar la riqueza de mariposas.
Finalmente hay que destacar que las diferencias encontradas entre los complejos de ciénagas evalaudas, las cuales albergan numerosas especies exclusivas, refuerzan la importancia de este estudio y la necesidad de conservación de los ambientes de humedales del Caribe y de su flora y fauna asociada.
Agradecimientos
Los autores expresan su agradecimiento a la Corporación Autónoma Regional de los Valles del río Sinú y río San Jorge (CVS) y a la Universidad Nacional de Colombia, Facultad de Ciencias por la cofinanciación del presente trabajo y al Instituto de Ciencias Naturales (ICN) por su apoyo, el presente trabajo se ejecutó dentro del Grupo de Biodiversidad y Conservación del ICN. Así mismo agradecemos a Hannier Pulido por facilitarnos la información de las mariposas de su tesis de grado para compararla con el presente manuscrito.
Bibliografía
Adams, M.J. 1985. Speciation in the Pronophiline butterflies (Satyridae) of Northern Andes, Second Symposium on Neotropical Lepidopterología (Arequipa, Perú). J. Res. Lep., Supplement: 133-49. [ Links ]
Andrade-C., M.G.2007. http://www.mariposasandinas.org/colombia_sp.html En El Proyecto Diversidad de las Mariposas Andinas Tropicales. [ Links ]
Andrade-C., M. G., Campos-Salazar, L.R., González-Montaña, L.A. & H.W. Pulido-B. 2007A. Santa María, mariposa alas y color. Serie de Guías de Campo del Instituto de Ciencias Naturales N° 2. Universidad Nacional de Colombia, Bogotá. D. C., Colombia. 248 pp. [ Links ]
Campos-Salazar., L. R. 2008. Estudio de la riqueza y composición de la comunidad de mariposas (Hesperioidea: Papilionoidea) en la cordillera oriental, bosque galilea (Tolima-Colombia). Tesis de Maestría en Entomología Facultad de Ciencias. Inédito. 87 pp. [ Links ]
De Vries, P. J. 1987. The Butterflies of Costa Rica and their natural history: Papilionidae, Pieridae, Nymphalidae, volumen I. Princeton University Press. New Jersey. 288 pp. [ Links ]
Cárdenas-Arévalo, G., O.V. Castaño-Mora, Carvajal-Cogollo, J.E. 2010. Comunidad de reptiles en humedales y áreas aledañas del departamento de Córdoba. Pp. 365-358. En J.O. Rangel Ch. (ed) 2010. Colombia Diversidad Biótica IX Ciénagas de Córdoba: Biodiversidad, ecología y Manejo Ambiental. Instituto de Ciencias Naturales-Universidad Nacional de Colombia [ Links ]
Coddington, J.A., Grisworld, E. Silva, D., Peñaranda, E. & Scott, S.1991. Designing and Testing Samplings Protocols to Estimate Biodiversity in Tropical Ecosystems, pp. 44-60. [ Links ]
Colwell, R.K. & Coddington, J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society, series B, 345:101-118. [ Links ]
De la Maza, R. 1987. Mariposas Mexicanas. Guía para colecta y determinación. Fondo de Cultura Económica, S.A.302 pp. [ Links ]
De Vries, P.J. 1988. Stratification of fruit-feeding nymphalid butterflies in a Costa Rican rainforest. Journal of Research on the Lepidoptera, 26:98-101. [ Links ]
Ehrlich, P.R. & P.H. Raven. 1964. Butterflies and plants: an study in coevolution. Evolution 18:586-608. [ Links ]
Fagua, G. 1996. Comunidad de Mariposas y Artropofauna Asociada con el Suelo de Tres Tipos de Vegetación de la Serranía de Taraira (Vaupés, Colombia). Una Prueba del Uso de Mariposas como Bioindicadores. Revista Colombiana de Entomología, 22(3):143-152. [ Links ]
Fagua, G., Amarillo, A. & Andrade-C., M. G. 1999 Mariposas como Indicadores del Grado de Intervención en la Cuenca del Río Pato (Caquetá-Colombia). En Andrade, M. G., G. Amat & F. Fernández (Eds). Insectos de Colombia, Estudios Escogidos. Colección Jorge álvarez Lleras 13: 285-315. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Bogotá [ Links ].
Gilbert, L. E. 1984.The Biology of Butterfly Communities. En Vane- Wright R. I. & P. R. Ackery (eds). The Biology of Butterflies (Symposia of the Royal Entomological Society of London, 11). London: Academic Press, 41—54. [ Links ]
Kremen, C. 1992. Assessing the Indicator Properties of Species Assemblages for Natural areas Monitoring. Ecological Applications, 2(2):303.217 [ Links ]
Kremen, C., R.K. Colwell, T.L. Erwin, D.D. Murphy, R.F. Noss & M.A. Saujayan. 1993. Terrestrial arthropod assemblages: their use in conservation planning. Conservation Biology, 7(4):796- 808. [ Links ]
Lamas, G., Callaghan C.J., Casagrande, M.M., Mielke, O.H., Pyrcz, T.W., Robbins, R.K. & Viloria A.L. 2004. Checklist: Part 4a Hesperioidea-Papilionoidea. Scientific Publishers. Gainesville, Florida 439 pp. [ Links ]
Le Crom, J.F., Llorente J., Constantino, L.M. & Salazar, J.A. 2004. Mariposas de Colombia, Tomo 2 Pieridae.Carlec Ltda., Colombia, Bogotá. 133 pp. [ Links ]
Magurran, A.E. 1988.Ecological Diversity and Its Measurement. Princeton University Press, New Jersey 179 pp. [ Links ]
Moreno, C.E. 2001. Manuales y Tesis SEA 1, Métodos para Medir la Biodiversidad. Zaragoza 84 pp. [ Links ]
Prieto, M. & Constantino, L.M. 1996. Abundancia, distribución y diversidad de mariposas (Lep. Rophalocera) en el río Tatabro, Buenaventura (Valle-Colombia). Bol. Mus. Ent. Univ. Valle. 4(2):11-18. [ Links ]
Pulido-B., H.W. 2008. Las mariposas (Lepidoptera: Hesperioidea, Papilionoidea) de la serranía del Perijá, Colombia. Análisis de distribución y diversidad. Facultad de Ciencias, departamento de Biología. Tesis de grado. Maestría en Ciencias. 120 pp. [ Links ]
Raguso, R.A. & Llorente, J. 1997. Papilionoidea. En González, E. S.,R. Dirzo&R. C. Vogt (eds.) Historia Natural de los Tuxtlas. Instituto de Ciencias de Biología, UNAM. México, pp. 257-291. [ Links ]
Rangel-Ch., J.O., Garay-Pulido, H. & Avella, A. 2010. Bosques Húmedos y secos circundantes a los complejos de humedales (ciénagas), en el departamento de Córdoba. Pp 214-216. En J.O. Rangel Ch. (ed) 2010. Colombia Diversidad Biótica IX Ciénagas de Córdoba: Biodiversidad, ecología y Manejo Ambiental. Instituto de Ciencias Naturales-Universidad Nacional de Colombia. [ Links ]
Rivera-Díaz, O. 2010. Flora. En: J. Orlando Rangel Ch. (ed.) Colombia Diversidad Biótica IX, Ciénagas de Córdoba: biodiversidad-ecología y manejo ambiental. Instituto de Ciencias naturales. Bogotá, pp 125-128. [ Links ]
Vélez,J. 1989. Mariposas de la zona cafetera en Colombia. Revista Lámpara, 23(108):10-19. [ Links ]
Recibido: abril 9 de 2010. Aceptado para su publicación: febrero 28 de 2011.
