










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
Print version ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.35 no.135 Bogotá Apr./June 2011
FITOPATOLOGÍA
1 Magíster en Fitopatología. Correo electrónico:cmqo@hotmail.com
2 Magíster en Fitopatología. Correo electrónico:mjbotero24@hotmail.com
3 Ph.D. Profesor Titular. Universidad de Caldas- Correo electrónico:castano_z@udecaldas.edu.co
Resumen
La higuerilla (Ricinus communis L.) tiene gran importancia a nivel industrial, porque es una alternativa para la obtención de biocombustible, además de los múltiples usos del aceite de ricino. Pero, al incrementarse las áreas de cultivo, se han detectado enfermedades no registradas previamente. Esta investigación se realizó con el fin de identificar el agente causante de la Necrosis de los brotes terminales de la higuerilla o Muerte apical, en las variedades Nordestina y Energía. Al microorganismo aislado, se le realizó pruebas de patogenicidad para cumplir con los postulados de Koch y una vez demostrada su patogenicidad, se determinó sus características culturales, morfológicas y bioquímicas. Los resultados demostraron que la enfermedad es causada por Xanthomonas campestris, la cual se manifiesta por la presencia de manchas de apariencia aceitosa y de color negro o café, en los brotes terminales y hojas jóvenes; además de exudado bacterial. La bacteria fue encontrada en semilla de las dos variedades en estudio, contaminando la testa y el embrión, constituyéndose en la fuente primaria de inóculo de la enfermedad.
Palabras clave: higuerilla, necrosis, brote, patogenicidad, bacteria, Xanthomonas.
Abstract
Castor (Ricinus communis L.) has become a very important crop for the industry, because it is an alternative for the production of biofuel, besides of its multiple uses like the castor-oil. But, as the planting area increases, several new diseases have been detected. This research was conducted in order to identify the causal agent of the necrosis of the terminal buds or apical death, on the varieties Nordestina and Energía. To the microorganism isolated, was conducted pathogenicity tests to demonstrate the Koch's postulates and once demonstrated its pathogenicity, was determined its cultural, morphological and biochemical characteristics. The results showed that the Necrosis of the terminal shoots of castor is caused by Xanthomonas campestris, which is manifested by the presence of oily-looking patches of brown or black color in the terminal buds and young leaves, in addition to bacterial exudates. The bacterium was found in the seeds of both varieties tested, contaminating the seed coat and embryo, concluding that the seed is the primary source of inoculum of this disease.
Key words: castor, necrosis, bud, pathogenicity, bacterium, Xanthomonas.
Introducción
La elevada demanda energética mundial ha hecho necesario buscar nuevas fuentes de energía, especialmente renovables y menos perjudiciales para el medio ambiente. El cultivo de higuerilla (Ricinus communis L.) se constituye en una alternativa válida para la obtención de biocombustible, además de tener un alto potencial para contribuir al desarrollo de la economía campesina, a corto, mediano y largo plazo. Es por esto, que su cultivo de manera comercial, en países como Colombia, puede constituirse en un medio rentable para los pequeños y medianos productores, además es un sistema productivo que puede convertirse en un gran generador de empleo y dinamizador de la economía nacional (CORPOICA, 2008; Da Silva Cunha, 2008).
Como esta especie ha sido considerada por mucho tiempo maleza y en Colombia hasta recientemente no se había cultivado de forma comercial, los resultados preliminares muestran la necesidad de profundizar en estudios sobre la viabilidad económica del cultivo y ampliar el conocimiento en cuanto al manejo fitosanitario de la higuerilla (CORPOICA, 2008).
Con respecto a la sanidad del cultivo, al incrementarse las siembras de higuerilla, se han detectado enfermedades ocasionadas por microorganismos que afectan los diferentes órganos de la planta, haciendo que esta no tenga un desarrollo normal y que los rendimientos sean alterados. Estas enfermedades y los microorganismos asociados, aún son objeto de estudio. Actualmente se adelantan estudios etiológicos en algunas enfermedades tales como: lesiones y pudriciones en cuello y raíces relacionadas con los hongos Fusarium sp., Rhizoctonia sp. y Pythium sp.; lesiones en tallo y defoliación de las ramas principales relacionadas con Colletotrichum sp. y Botryodiplodia sp.; lesiones foliares relacionadas con Cercospora sp., Alternaria sp. y Cladosporium sp. y lesiones y pudriciones en fruto ocasionadas por Botrytis sp. (CORPOICA, 2008).
Esta investigación se enfocó en una patología que afecta a los brotes terminales de la higuerilla y de la cual se desconocía el agente causante, impidiendo la implementación de un plan de manejo de la enfermedad. Por lo tanto, el objetivo de este trabajo fue identificar y caracterizar el agente causante del Necrosamiento de los brotes terminales o Muerte apical de la higuerilla.
Materiales y métodos
La investigación se realizó en la granja Montelindo, de la Universidad de Caldas, situada en Palestina (Caldas), vereda Santágueda, con altitud de 1.010 m, temperatura media de 23°C, humedad relativa de 80% y precipitación promedio anual de 1.800 mm; y en el laboratorio de Fitopatología, ubicado en el Departamento de Fitotecnia de la Universidad de Caldas, con altitud de 2.088 m, temperatura media de 17,5°C y humedad relativa de 73%.
La recolección de muestras se hizo en dos cultivos de higuerilla, establecidos con las variedades Nordestina y Energía, ubicados en la granja Montelindo. Para la identificación y caracterización de los microorganismos fitopatógenos asociados a la patología estudiada, se tomaron muestras de brotes, hojas tiernas y semilla de las dos variedades. Se realizó un solo muestreo, en el cual se avaluaron 80 plantas para la variedad Nordestina y 43 para la variedad Energía, correspondiente al 10% de la población. El muestreo de las plantas se hizo al azar.
El reconocimiento y descripción de los síntomas asociados con la enfermedad, se hizo con base a la observación en campo de brotes y hojas tiernas de higuerilla naturalmente infectadas, revisándose los brotes y hojas de cada planta, desde la base hasta la parte apical.
Aislamiento de bacterias fitopatógenas de brotes y hojas tiernas
El aislamiento se hizo por medio de siembra directa del tejido afectado, en agar nutritivo (AN, peptona 5 g/L, extracto de carne 4 g/L, agar 15 g/L). Se tomaron trozos de tejido sano y enfermo de 4 mm2, los cuales se colocaron en hipoclorito de sodio al 2,5% por un min. y se enjuagaron con agua destilada estéril, enseguida los trozos de tejido se dejaron secar por un min. sobre toallas de papel y posteriormente se transfirieron al medio de cultivo sembrándose cuatro trozos por caja Petri, éstas se sellaron y se incubaron 24°C en una incubadora marca WTB Binder, por 72 h. Al cabo de este tiempo, se observaron las siembras y de las colonias bacterianas que se desarrollaron, se hicieron repiques para purificarlas y someterlas a la tinción de Gram. La patogenicidad se comprobó mediante los postulados de Koch (Botero & Ramírez, 1997), en brotes y hojas de higuerilla silvestre.
Aislamiento de bacterias de semilla
Siembra directa: Se evaluaron 160 semillas por variedad. La semilla fue desinfectada con alcohol etílico al 70%, por un min. y con hipoclorito de sodio al 2,5% por cuatro min., seguidamente, se enjuagó con agua destilada estéril (ADE) por dos min. y se secó con toallas de papel, posteriormente, la semilla se sembró en AN y agar peptona sucrosa (APS, nitrato de calcio 0,35 g/700 mL, sulfato ferroso 0,35 g/ 700 mL, fosfato dibásico de sodio 1,4 g/700 mL, peptona 3,5 g/700 mL, sucrosa 14 g/700 mL, agar 10,5 g/700 mL), en cada medio se sembraron 80 semillas del total utilizado para cada variedad y se colocaron ocho semillas por caja de Petri, finalmente éstas se sellaron y se incubaron a 24°C, en una incubadora marca WTB Binder, por 72 h. Al cabo de este tiempo, se evaluó el porcentaje de contaminación de la semilla con bacterias fitopatógenas, lo cual se corroboró con la tinción de Gram (Mehta et al., 2005).
Centrifugación: Se muestrearon 160 semillas por variedad. Previo a la centrifugación, la semilla se esterilizó con alcohol al 70% por un min. y con hipoclorito al 2,5% por cuatro min., después, se lavaron con ADE por dos min. y se secaron con toallas. Luego, las semillas se transfirieron a un recipiente de vidrio con 600 mL de solución salina al 0,5%, el cual se agitó en un agitador mecánico marca AREC, durante seis h. continuas. Pasado el periodo de agitación, la solución obtenida se distribuyó por medio de filtración, en cinco tubos de 10 mL y estos se colocaron en una centrífuga marca Clay Adams, donde fueron centrifugados por 10 min., a 3.800 rpm, el precipitado obtenido en cada tubo se suspendió en 5 mL de ADE, se realizó una homogenización y se hizo la siembra de las suspensiones en AN y APS. En cada caja de Petri, se sembraron cuatro muestras, de 20 µL de la solución, las cuales se esparcieron en ziz zag, con un asa bacterial redonda estéril. Finalmente, las cajas se sellaron y se llevaron a incubar a 24°C, en una incubadora marca WTB Binder, por espacio de 72 h., pasado este tiempo, se observaron las siembras y se evaluó el porcentaje de contaminación, contabilizando el número de semillas con presencia de bacterias fitopatógenas, confirmada con la tinción de Gram (Mehta et al., 2005). Para cada variedad se sembraron en total 160 semillas, 80 en AN y 80 en APS.
Macerado de testa y embrión: Se muestrearon 360 semillas por variedad y 160 por tejido evaluado, en tubos de ensayo con ADE, para un total de 80 semillas por cada tejido y utilizando dos semillas por repetición.
Previo a los macerados, las semillas se esterilizaron con alcohol al 70% por un min. y con hipoclorito de sodio al 2,5% por cuatro min., se enjuagaron con ADE y se secaron con toallas de papel estériles. Para el macerado de testa, se tomaron dos semillas, se les retiró la testa, la cual se colocó en un recipiente estéril, se le adicionó 0,5 mL de ADE y se molió en un mortero estéril por 2 o 3 min., a continuación el macerado se paso a un tubo de ensayo con 5 mL de ADE y se dejo ahí por espacio de 6 h., al cabo de ese tiempo, de cada repetición se tomó 30 µL de la solución, se depositó sobre AN y se realizó una siembra por agotamiento, finalmente, las cajas de Petri se sellaron y se llevaron a incubar a 24°C, por 72 h. Para el macerado del embrión el procedimiento fue el mismo, solo que en éste caso se utilizaron dos embriones (Mehta et al., 2005). Pasado las 72 h., se observaron las siembras y se calculó el porcentaje de contaminación, tanto en testa como en embrión, contando el número de semillas con presencia de bacterias fitopatógenas, en cada tejido, lo cual se verificó con la tinción de Gram.
Pruebas de patogenicidad
Para las pruebas de patogenicidad se utilizó solución bacteriana a una concentración de 3 x 108 bacterias/mL, la cual corresponde al tubo No. 1 de la escala de opacidad de McFarland (Botero & Ramírez, 1997) y se emplearon plántulas de higuerilla sanas, de 30 días de edad, de las variedades Nordestina y Energía, de las cuales se inocularon 20 plántulas por variedad, además se tuvieron 10 testigos, tanto para Nordestina como para Energía, los cuales fueron inoculados con agua destilada estéril.
La inoculación se hizo por medio de heridas realizadas en hojas tiernas y brotes apicales, las cuales se hicieron con un par de cuchillas, insertadas en un bloque de madera de 7 x 5 cm y con bisturí, respectivamente, sumergidos con anterioridad en la solución bacteriana. Posteriormente, a las plántulas se les adecuó una cámara húmeda, colocándoles una bolsa plástica transparente a cada una, con una mota de algodón humedecido en su interior (Castaño- Zapata & del Río, 1997; Nico et al., 2006). Las plántulas inoculadas se dejaron en cámara húmeda por 3 d. y al cabo de ese tiempo se revisaron para valorar el resultado de la prueba de patogenicidad. Se evaluó la incidencia de la enfermedad, tomándose como positivo aquellas plántulas cuyos brotes y hojas tiernas expresaron los síntomas de la enfermedad. Finalmente, de las plántulas afectadas se hizo el reaislamiento de la bacteria en AN y se realizó su identificación, con el fin de complementar los postulados de Koch (Nico et al., 2006).
Pruebas culturales, morfológicas y bioquímicas
La identificación y caracterización de las cepas bacterianas, se hizo determinando las características que se indican a continuación:
Características culturales. Se observó la forma, color, opacidad, superficie, bordes, pigmento y consistencia de las colonias, en agar, extracto de levadura, dextrosa, carbonato de calcio (YDC, extracto de levadura 10 g/L, dextrosa 20 g/L, carbonato de calcio 20 g/L, agar 15 g/L) (Botero & Franco, 2007).
Características morfológicas. Se estableció con la tinción de Gram y su confirmación mediante solubilidad en KOH al 3%, además se observó la forma de la bacteria, al microscopio a 100X (Botero & Franco, 2007).
Características bioquímicas. Se determinó utilizando las siguientes pruebas: producción de la enzima catalasa, oxidasa, crecimiento en agar nutritivo glucosa, crecimiento en agar Mac Conkey, oxidación-fermentación para glucosa (Hugh- Leifson-OF), indol-movilidad-ácido sulfhídrico (SIM), licuefacción de la gelatina, utilización del citrato, agar-triple-azúcar hierro (TSI), producción de ácido a partir de arabinosa, glucosa y mannosa, crecimiento en agar nutritivo a 36 °C, hidrólisis del almidón, crecimiento en agar bilis esculina, crecimiento en agar, extracto de levadura, YDC, leche tornasolada, urea, crecimiento en agar sangre y crecimiento en agar eosina y azul de metileno (EMB) (Schaad, 1988).
Resultados y discusión
Reconocimiento de síntomas: Al comienzo de la enfermedad, se observaron lesiones o puntos de color negro o café en la base de los brotes terminales y con el tiempo, el necrosamiento del tejido avanzó hasta comprometer todo el brote, imposibilitando la formación de hojas nuevas. Finalmente el brote necrosado se seco y se cayó. Las lesiones inicialmente eran acuosas o húmedas y de apariencia aceitosa, además, se podía observar en los puntos de unión entre el tallo y el brote, y sobre el tejido necrosado un exudado bacterial café. Cuando el brote no había sido atacado completamente, se desarrollaba la hoja, pero ésta se deformaba en las partes donde era afectada por el patógeno, porque las porciones de tejido necrosado se desprendían y caían (Figura 1).
Referente a la edad de susceptibilidad de la planta, se ha observado que la enfermedad se puede presentar en brotes de plantas desde las seis semanas de edad, afectando a los retoños desde el estrato inferior de la planta, hasta el superior; sin embargo, esta patología no afecta el desarrollo general de la planta, porque esta continua creciendo normalmente.
Según Swings & Civerolo (1993), la necrosis de los tejidos, es un síntoma característico de muchas especies del género Xanthomonas, especialmente de aquellas que causan manchas en las hojas, ya que al avanzar la enfermedad el fenol de la vacuola reacciona con el citoplasma y la membrana, envolviéndolos de oxidasas y glicosidasas; la oxidación del fenol es lo que resulta en el desarrollo de una pigmentación café o negra de los tejidos de la planta. Estos autores, también afirman, que las lesiones acuosas se vuelven necróticas al final de la infección, probablemente debido a la desintegración continua de los tejidos y en infecciones severas las lesiones pueden coalecer, destruyendo áreas grandes de la lámina foliar, lo que se conoce como Tizón bacterial. Finalmente el área necrótica se separa de la parte sana del tejido y se desprende.
Lozano & Sequeira (1974), reportan que Xanthomonas campestris pv. manihotis, causante del Tizón bacterial de la yuca, no invade los tejidos maduros de la mayoría de los cultivares de esta planta, por eso las ramas adultas y los tallos de las plantas infectadas permanecen sanos; estos autores indican que la diseminación de la bacteria a partir de plantas infectadas, se da hacia las plantas jóvenes, puesto que la penetración a través de tejidos tiernos probablemente es mayor en comparación con los tejidos maduros.
Aislamiento de bacterias fitopatógenas de brotes y hojas tiernas de higuerilla
De las siembras directas de tejido, se obtuvo colonias bacterianas amarillas, confirmándose la presencia de bacterias fitopatógenas con la tinción de Gram (Figura 2). De estos cultivos, se obtuvieron 67 aislamientos bacterianos, que fueron obtenidos de cada tipo de tejido evaluado (brote y hoja de Nordestina, brote y hoja de Energía), los cuales se caracterizaron por presentar bacilos Gram negativos y formar colonias de color amarillo y de apariencia butirosa.
Posteriormente, a las cepas bacterianas se les comprobó su actividad patogénica por medio de pruebas de patogenicidad realizadas en brotes y hojas tiernas de higuerilla silvestre; de esta manera se identificaron 18 cepas altamente patogénicas, puesto que a los tres días después de la inoculación, mostraron los síntomas de la enfermedad. Todos los aislamientos fueron rayados en YDC, obteniéndose colonias amarillas, de superficie lisa, brillantes y de apariencia butirosa (Figura 3).
Schaad (1988), indica que las colonias de Xanthomonas, son mucoides, convexas y brillantes sobre agar nutritivo glucosado (NGA) y agar, extracto de levadura, dextrosa, carbonato de calcio (YDC), también afirma que una vía de apoyo para confirmar la presencia del género Xanthomonas es la identificación del pigmento amarillo, el cual le da el color a las colonias.
Aislamiento de bacterias fitopatógenas de semilla de higuerilla
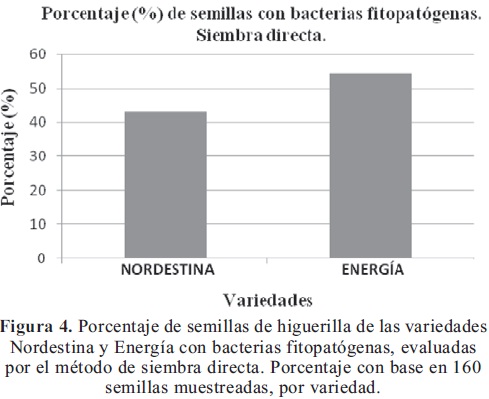
Aislamiento por siembra directa: Se aislaron bacterias de colonias amarillas, las cuales resultaron ser fitopatógenas, corroborándose con la tinción de Gram. Respecto al porcentaje de semillas con bacterias fitopatógenas, se encontró que el 43% de las semillas de Nordestina estaba contaminada y el 54% de Energía (Figura 4).
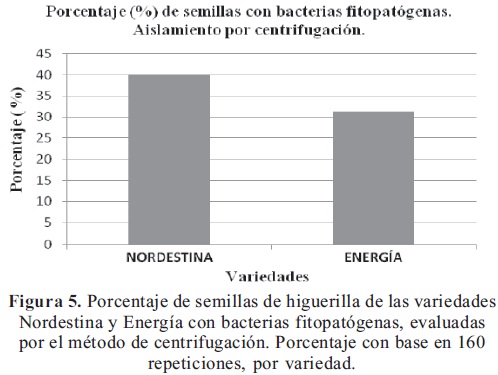
Aislamiento por centrifugación. Se observó el crecimiento de colonias bacterianas de color amarillo, confirmándose la presencia de bacterias fitopatógenas con la tinción de Gram. Se encontró bacterias fitopatógenas en el 40% de los cultivos de la variedad Nordestina y 31% en los cultivos de Energía (Figura 5).
Aislamiento por macerado de testa y embrión. Se aislaron colonias bacterianas fitopatógenas, tanto de las siembras de macerado de testa, como de las siembras de macerados de embrión. Se observó bacterias fitopatógenas en 79% de las testas de la variedad Nordestina y 28% en el embrión. Así mismo, en las testas de Energía se detectó un 31% de bacterias fitopatógenas y 10% en el embrión (Figura 6). Como se desprende de estos resultados, la bacteria se encuentra en mayor porcentaje sobre la testa de la semilla, siendo mayor en Nordestina que en Energía; y, muy significativamente en los tejidos internos de las semillas de ambas variedades, lo cual dificulta su detección y manejo.
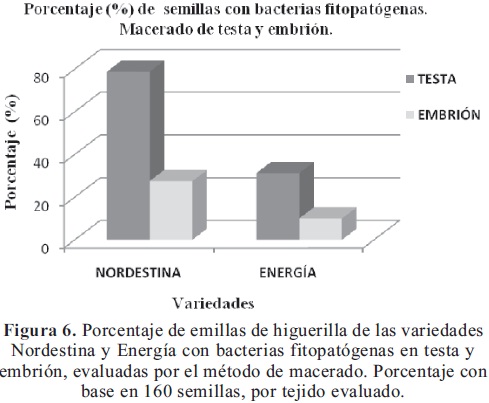
Teniendo en cuenta que en el muestreo de semilla de las variedades Nordestina y Energía, evaluada por los tres métodos, se encontraron bacterias fitopatógenas, corroborado con pruebas de patogenicidad, se dedujo que la semilla es la fuente primaria de inóculo y que posiblemente podría considerarse como un medio de diseminación del agente causante de la Necrosis de los brotes terminales de la higuerilla.
Resultados similares fueron reportados por Mehta et al. (2005), al recuperar a Xanthomonas axonopodis pv. malvacearum, de semilla de algodón, naturalmente infectada, las colonias típicas de Xanthomonas crecieron alrededor de la semilla, muestreada por medio de siembra directa en un medio semiselectivo, las colonias fueron amarillas, lisas, convexas y brillantes.
Swings & Civerolo (1993), afirman que las bacterias de muchos patovares de Xanthomonas campestris, están asociadas con la semilla de sus plantas hospedantes, la semilla puede encontrarse contaminada externamente e infectada internamente con éste género bacterial. Lo anterior confirma los resultados obtenidos en esta investigación, puesto que la bacteria fitopatógena aislada de la semilla de higuerilla se encontró tanto en la testa, como en el embrión.
Se han reportado diferentes formas de contaminación de semilla para las enfermedades causadas por bacterias; la contaminación puede ser externa, como consecuencia del contacto de la semilla con poblaciones bacterianas sobre tejidos enfermos, o a una alta cantidad de inóculo sobre los frutos, o por el roce con residuos que acarrean una alta población bacterial. La semilla puede ser infectada por la invasión de las estructuras florales y también puede ser contaminada internamente, por el movimiento sistémico del patógeno de plantas madres infectadas a plantas hijas (Walcott et al., 2003).
Desarrollo de las pruebas de patogenicidad
Los resultados de las pruebas de patogenicidad, expresados en porcentaje de incidencia de la enfermedad, demostraron la patogenicidad de la bacteria, la cual es capaz de reproducir los síntomas de la Necrosis de los brotes terminales o Muerte apical de la higuerilla.
La incidencia de la enfermedad en las plántulas de la variedad Nordestina fue del 100%, en cambio en los testigos de esta variedad no se presentó la enfermedad, y para la variedad Energía, el comportamiento fue el mismo, con un porcentaje de infección del 100%, en las plántulas inoculadas con la bacteria y de 0% en las plántulas testigo (Figura 7).
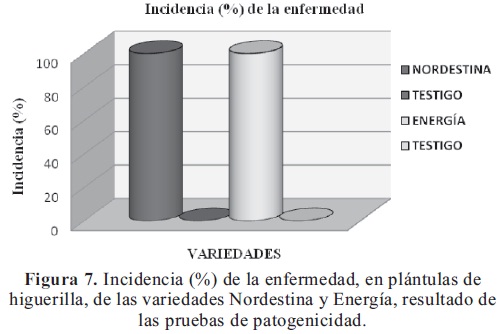
Estas pruebas también permitieron determinar que el periodo de incubación de esta bacteria, bajo condiciones controladas de humedad, es de tres días en las dos variedades de higuerilla, porque las plántulas presentaron los síntomas de la enfermedad a los tres días después de la inoculación, observándose en los puntos de corte en los brotes y hojas tiernas, lesiones de color café y de apariencia acuosa, evidencia del inicio de la necrosis de los tejidos. También se pudo observar la presencia de un exudado bacterial de color café en las lesiones. Este periodo de incubación es similar al obtenido por Botero et al., (2006), en el patosistema mayacuyá-Xanthomonas campestris pv. passiflorae.
Finalmente, a los cinco días los brotes y hojas tiernas se encontraban totalmente invadidas por el patógeno, manifestándose en una desintegración total de los tejidos (Figura 8 A).
Por último, de las plántulas inoculadas y enfermas se tomaron muestras de brotes y hojas tiernas, para hacer el reaislamiento de la bacteria fitopatógena, obteniéndose cepas bacterianas con características típicas del género Xanthomonas y por lo tanto similares a la utilizada para la inoculación, completándose de esta manera los postulados de Koch.
Por medio de la comprobación de la asociación intima de Xanthomonas sp., con las plantas de higuerilla, afectadas por la Necrosis de los brotes terminales, su aislamiento en medio de cultivo puro, seguido por la inoculación de ésta en plántulas sanas, la posterior reproducción de la sintomatología de la enfermedad en las mismas y finalmente el reaislamiento del mismo patógeno en cultivo puro, se afirma que Xanthomonas sp. es el agente causal de la Necrosis de los brotes terminales de higuerilla.
Tushemereirwe et al. (2004), hicieron un primer reporte de Xanthomonas campestris pv. musacearum, atacando a banano, en Uganda. Ellos comprobaron por medio de los postulados de Koch, que la bacteria mencionada es la causante de un desorden patológico caracterizado por un rápido amarillamiento y marchitamiento de hojas jóvenes, ocasionalmente una muerte descendente, que se inicia en la parte floral y pudrición de la parte interna del fruto.
Pruebas culturales, morfológicas y bioquímicas
Para la identificación y caracterización del agente causante de la Necrosis de los brotes terminales de la higuerilla, se evaluaron 42 cepas bacterianas distribuidas así: 18 cepas obtenidas a partir de la siembra directa de brotes y hojas tiernas, de las dos variedades, 12 cepas obtenidas a partir del muestreo de semilla y 12 cepas provenientes de las pruebas de patogenicidad.
En cuanto a las características culturales, todas las cepas se evaluaron en YDC, obteniéndose el desarrollo de colonias de color amarillo, de forma redonda, de bordes enteros, brillantes, mucoides, de superficie lisa, elevación convexa y de apariencia butirosa, características típicas del género Xanthomonas. Con respecto a las características morfológicas, todos los aislamientos exhibieron bacilos Gram negativos, de tamaño pequeño y uniforme, con bordes redondeados y de presencia individual.
Referente a la caracterización bioquímica, todos los aislamientos fueron sometidos a 22 pruebas. Los resultados de éstas pruebas para determinar la identidad de la bacteria como perteneciente al género Xanthomona y a la especie campestris, se presentan en la Tabla 1.
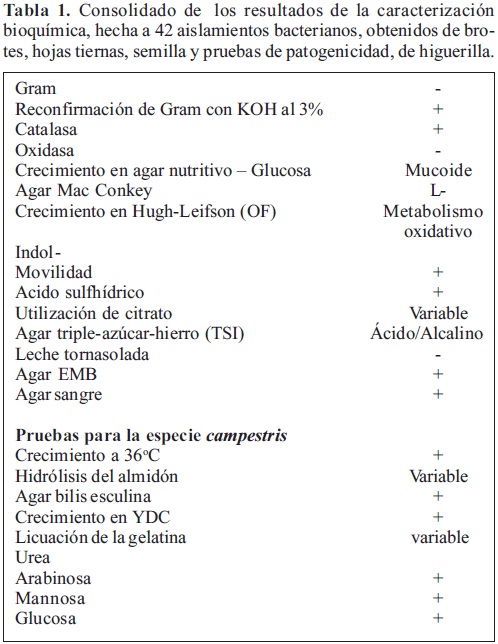
En las pruebas bioquímicas básicas para la especie Xanthomonas campestris, los 40 aislamientos reaccionaron positivamente a catalasa, movilidad, producción de acido sulfhídrico e hidrólisis de la esculina; los mismos reaccionaron negativamente a oxidasa, producción de indol y urea; presentaron metabolismo oxidativo en Hugh-Leifson (OF) y la reacción de las cepas fue variable a hidrólisis del almidón, licuación de la gelatina y utilización de citrato, todos crecieron a 36°C y produjeron ácido a partir de arabinosa, mannosa y glucosa. Finalmente se hizo una comparación de los resultados de la caracterización bioquímica de los aislamientos bacterianos obtenidos en ésta investigación, con patrones de referencia de X. campestris y X. axonopodis (Tabla 2).

Schaad (1988), indica que las bacterias del género Xanthomonas, son bacilos Gram negativos, aeróbicas, catalasa positiva, no utilizan la asparagina como una sola fuente de carbono y nitrógeno y la producción de ácido a partir de carbohidratos es débil.
La caracterización de la especie, comparándola con X. campestris y X. axonopodis, mediante pruebas culturales, morfológicas, y bioquímicas, permitió concluir que la bacteria aislada de los brotes, hojas tiernas y semilla, de higuerilla, se comporta como Xanthomonas de la especie campestris; por lo tanto, el agente causal de la Necrosis de los brotes terminales o Muerte apical, de la higuerilla es Xanthomonas campestris.
De acuerdo a Schaad (1988) y Holt (1994), X. campestris presenta crecimiento mucoide sobre agar extracto de levadura, dextrosa, carbonato de calcio (YDC) o agar, glucosa, extracto de levadura, carbonato de calcio (GYCA), crece a 35°C, hidrolisa la esculina, la licuefacción de la gelatina es variable (+/-), la digestión de proteína es positiva, la producción de urea es negativa y produce acido a partir de arabinosa, glucosa y manosa. Teniendo en cuenta esto, se puede afirmar que la bacteria evaluada, pertenece a esta especie, ya que en las pruebas mencionadas, los resultados fueron los mismos.
Conclusiones
La Necrosis de los brotes terminales o Muerte apical de la higuerilla, de las variedades Nordestina y Energía, es una enfermedad de origen bacterial, cuyo agente causante es Xanthomonas campestris, el cual se identificó y caracterizó por medio de pruebas de patogenicidad y caracteres culturales, morfológicos y bioquímicos.
X. campestris, es capaz de atacar a brotes y hojas tiernas de éstas variedades, causándoles lesiones cafés y de apariencia acuosa. En los brotes la infección causa una necrosis total del tejido a medida que la enfermedad avanza, imposibilitando la formación de hojas nuevas.
En las pruebas de patogenicidad se logró obtener los síntomas típicos de la enfermedad sobre los brotes y hojas tiernas de las plántulas de higuerilla de las dos variedades, tres días después de la inoculación.
Se estableció que la fuente primaria de inóculo en éste patosistema, es la semilla asintomática.
Bibliografía
Botero, M. J. & G. Franco. 2007. Identificación y caracterización preliminar del agente causal de la Mancha necrótica de las hojas de la mora (Rubus glaucus) en el municipio de Trujillo (Valle del Cauca, Colombia). Revista Corpoica - Ciencia y Tecnología Agropecuaria 8(2):22-25. [ Links ]
_____. & M. C. Ramírez. 1997. Identificación y caracterización de bacterias asociadas con enfermedades en maracuyá (Passiflora edulis var. flavicarpa), en dos zonas productoras del departamento de Caldas. Trabajo de grado (Especialista en Microbiología). Universidad Católica de Manizales. Post-Grado de Microbiología. 191 p. [ Links ]
_____., M. C. Ramírez & J. Castaño-Zapata. 2006. Determinación del periodo de incubación de Xanthomonas campestris pv. passiflorae Pereira, agente causante de la Bacteriosis del maracuyá (Passiflora edulis var. flavicarpa Degener). Boletín Fitotecnia N0. 107. Enero 2006. [ Links ]
Castaño Zapata, J. & L. Del Rio Mendoza. 1997. Manual para el diagnóstico de hongos, bacterias, virus y nematodos fitopatógenos.1ed. Manizales: Centro Editorial Universidad de Caldas, Manizales, Colombia - Zamorano Academic Press, Tegucigalpa, Honduras. 210 p. [ Links ]
Corpoica. 2008. Higuerilla: Alternativa productiva, energética, y agroindustrial para Colombia. Primera ed. Rionegro 2008. 35 p. [ Links ]
Da Silva Cunha, M. A.2008. Análisis molecular de la variabilidad genética entre genotipos de Ricinus communis L. revelada por marcadores RAPD. Brasil. 52 p. [ Links ]
Holt, J. G., N. R. Hrieg, P. H. Sneath, J. T. Staley & S. Williams. 1994. Bergey's manual of determinative bacteriology. 9th ed. USA: Williams & Wiilkins, Ed. 787 p. [ Links ]
Lozano, J.C. & L. Sequeira. 1974. Bacterial blight of cassava in Colombia: Etiology. Phytopathology 64:74-82. [ Links ]
Mehta, Y. R., C. Bonfeti & V. A. Bolognini. 2005. A semi selective agar médium to detect the presence of Xanthomonas axonopodis pv. malvacearum in naturally infected cotton seed. Fitopatología Brasilera 30(5):489-495. [ Links ]
Nico, A.I, A. M. Alippi, E. Dal Bo & L. B. Ronco. 2006. Interacción de Pseudomonas corrugata y Pseudomonas viridiflava y diferentes genotipos de tomate. Revista de la Facultad de Agronomía 106:37-45. [ Links ]
Schaad, N.W. 1988. Laboratory guide for identification of plant pathogenic bacteria. St. Paul. University of Georgia. Departament of Plant Pathology. 164 p. [ Links ]
Swings, J.G. & E. L. Civerolo. 1993. Xanthomonas. First ed. London: Chapman & Hall. 399 p. [ Links ]
Tushemereirwe, W., A. Kangire, F. Ssekiwoko, L. C. Offord, J. Crozier, E. Boa, M. Rutherford & J. J. Smith. 2004. First report of Xanthomonas campestris pv. musacearum on banana in Uganda. Plant Pathology 53:802. [ Links ]
Walcott, R.R., R. D. Gitaitis & A. C. Castro. 2003. Role of blossoms in watermelon seed infestation by Acidovorax avenae subsp. citrulli. Phytopathology 93:528-534. [ Links ]
Recibido: febrero 16 de 2011.
Aceptado para su publicación: junio 1 de 2011.
