










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
Print version ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.35 no.137 Bogotá Oct./Dec. 2011
BOTÁNICA
1 Investigador independiente, Claustro de San Agustín, Villa de Leyva, Boyacá, Colombia. Correo electrónico: hummendoza@gmail.com
2 Real Jardín Botánico RJB-CSIC, Plaza de Murillo 2, 28014 Madrid, España. Correo electrónico: jlfernandeza@rjb.csic.es
Resumen
Centronia es un género neotropical de 15 especies que presenta problemas de circunscripción con otros géneros de la Tribu Merianieae como son Meriania y Graffenrieda. Para evaluar su monofilia, se realizó un análisis cladístico con base en caracteres morfológicos. En el muestreo se incluyeron 45 especies de 10 géneros; como grupo más externo se utilizó una especie del género Miconia de la tribu Miconieae; para Centronia se consideraron 13 especies. Un total de 103 caracteres morfológicos fueron utilizados en el análisis. Centronia es un grupo polifilético con 10 especies que se asocian al género Meriania, una al género Wurdastom, dos de posición genérica incierta y un grupo monofilético de dos especies que conforman a Centronia "sensu stricto". Centronia s. str. no posee sinapomorfías y los caracteres que lo sustentan también se encuentra en Graffenrieda, pero la resolución del análisis no permitió concluir aun sobre su transferencia a este género. Se propone que el grupo de especies de Centronia del norte de los Andes sea transferido a Meriania.
Palabras clave: Centronia, filogenia, Graffenrieda, Melastomataceae, Meriania, Merianieae, Wurdastom.
Abstract
Centronia is a neotropical genus with 15 species that present circumscription problems with other genus of the tribe Merianieae like Meriania and Graffenrieda. To test its monophylie, a cladistic analysis of the genus based on morphological characters was carried out. Fourth five species of 10 genera there were included in the sample; one species of Miconia from tribe Miconieae was the most external group; of Centronia we considered 13 species. A whole of 103 morphological characters was employed. Centronia is a polyphyletic group with 10 species associated to the genus Meriania, one to Wurdastom, 2 of generical uncertain position and 2 species that conform the monophyletic group Centronia "sensu stricto". Centronia s.str. don't has sinapomorphies its characters support also been in Graffenrieda, however the resolution of this analysis is not enough to merge it in Graffenrieda. Here we propose that the Andean group of this genus should be transfer to Meriania.
Key words: Centronia, Graffenrieda, Melastomataceae, Meriania, Merianieae, phylogeny, Wurdastom.
Introducción
Centronia D.Don es un género neotropical conformado por 15 especies distribuidas entre Colombia, Venezuela y Perú. Las especies se encuentran desde el nivel del mar hasta zonas de páramos, pero la mayoría crecen en el norte de los Andes entre los 1200 y 3000 m de altitud. Dos de sus especies se encuentran en el Escudo Guayanés de Colombia y Venezuela y otras dos en las zonas de la Cordillera del Cóndor en Ecuador y Perú (Wurdack, 1980; Lozano & Becerra, 1999a; Mendoza & Ramírez, 2006). El género fue descrito por Don (1823) y su especie tipo es C. laurifolia D.Don.
Centronia se ubica dentro de la tribu Merianieae, la cual está constituida por 15 géneros y alrededor de 280 especies neotropicales (Mendoza-Cifuentes & Fernández-Alonso, 2010). Triana (1871) y Cogniaux (1891) produjeron monografías del género como parte de sus tratamientos de toda la familia, pero desde entonces no se ha realizado una evaluación taxonómica integral y la literatura existente sólo comprende publicaciones de nuevas especies y tratamientos en floras regionales (Almeda, 1993; Lozano & Becerra, 1999a; Wurdack, 1973, 1980).
La circunscripción de Centronia comprende la presencia de cáliz caliptrado con dehiscencia circuncísil y estambres con poro de las anteras orientado ventralmente (Wurdack, 1973). No obstante, estos caracteres son variables dentro del género y además se encuentran en otros de la misma tribu como son Graffenrieda DC. y Meriania Sw. (Mendoza-Cifuentes & Fernández-Alonso, 2010). Todas las especies de Centronia poseen caliptras, sin embargo, son de diferentes formas y tienen dehiscencias variables, no sólo circuncísil (Mendoza-Cifuentes & Fernández- Alonso, 2010). La orientación del poro de la antera, que se ha utilizado para separar este género de Meriania (Wurdack, 1973), es polimórfico en Centronia, pues el 67% de sus especies presentan poros dorsales, el 26% ventrales y el 7% apicales (Mendoza-Cifuentes & Fernández-Alonso, 2010). Centronia es el único género en Merianieae con un taxón con flores haplostemonas y con caracteres que se asocian al género Wurdastom B.Wall., de posición tribal incierta (Mendoza-Cifuentes & Fernández-Alonso, 2010). Estas variaciones e inconsistencias en los caracteres diagnósticos no permiten actualmente diferenciar a Centronia de géneros cercanos como Graffenrieda y Meriania y es un indicio de que este taxón no es un grupo natural.
Considerando lo expuesto, este trabajo evaluó la monofilia de Centronia por medio de un análisis filogenético basado en caracteres morfológicos. También buscó establecer pautas para redefinir su circunscripción o la posible inmersión de sus especies en otros géneros de la tribu.
Métodos
Taxones analizados
Se consideraron 45 especies que representan 10 géneros, 9 de ellos pertenecientes a Merianieae. De las especies se evaluaron un total de 192 ejemplares de 11 herbarios en Colombia, Ecuador y EE.UU. (Apéndice 1). Como grupo más externo se utilizó a Miconia dodecandra (Desr.) Cogn. de Miconieae, tribu hermana de Merianieae (Clausing & Renner, 2001; Michelangeli et al., 2004). De Merianieae se consideraron los siguientes géneros reconocidos para la tribu (Renner, 1993): Phainantha Gleason (1 sp), Pachyloma DC. (1 sp), Huberia DC. (1 sp),Meriania (13 spp), Graffenrieda (7 spp), Adelobotrys DC. (3 sp) y Axinaea Ruiz & Pav. (3 spp).
Como grupo interno de Centronia se consideraron 13 de las 15 especies del género. Se excluyeron C. pulchra Cogn. por contar con poco material y ser muy similar a C. brachycera (Naudin) Triana (Wurdack, 1973), y C. eximia (Naudin) Triana por no encontrarse ejemplares en los herbario visitados o tener dudas de las determinaciones. También se excluyeron especies inéditas (nomen nudum) como C. nobilis, C. lehmannii y C. subrotundifolia. De C. reticulata Triana se utilizaron ejemplares determinados como afín, pero cuya morfología y descripción coinciden con la del tipo. Dada la afinidad de C. ruizii Lozano con el género Wurdastom, en el análisis filogenético se incluyeron dos especie de este último.
Caracteres y estados
Se examinaron ejemplares vivos (8 especies) cuando fue posible. Se colectaron flores y frutos en alcohol para realizar disecciones. En caso de no contar con material fresco, se rehidrataron con agua caliente estructuras florales y del fruto obtenidas de ejemplares de herbario plenamente identificados y en algunos pocos casos se observaron directamente los tipos. Se obtuvieron semillas de ejemplares de herbario, las cuales fueron fotografiadas con un microscopio electrónico de barrido (SEM). Las semillas fueron recubiertas con Gold-Paladium por 2 minutos y luego fueron fotografiadas en un microscopio marca Hitachi 4500S-SEM. Para algunas especies se establecieron los estados de caracteres con observación por medio de un microscopio óptico.
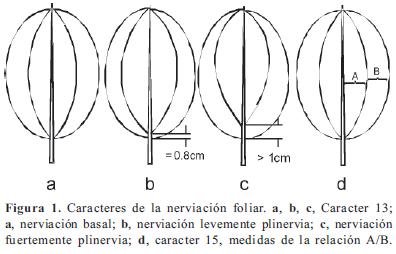
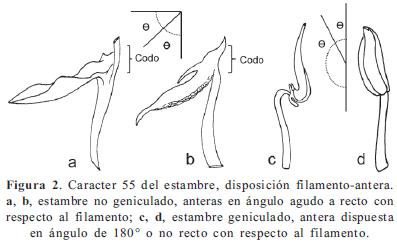
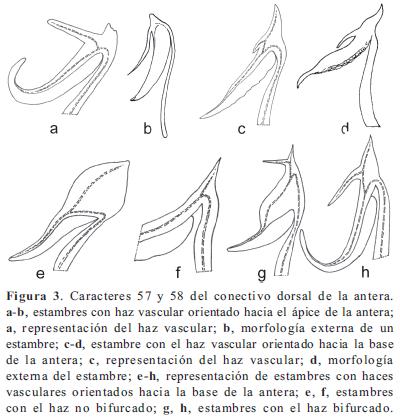
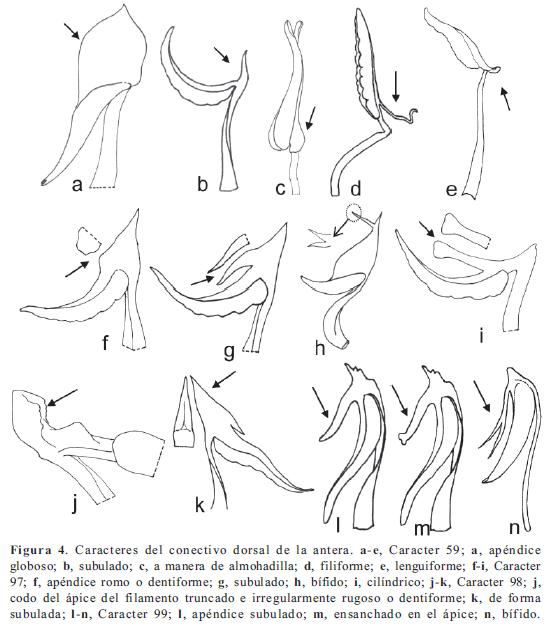
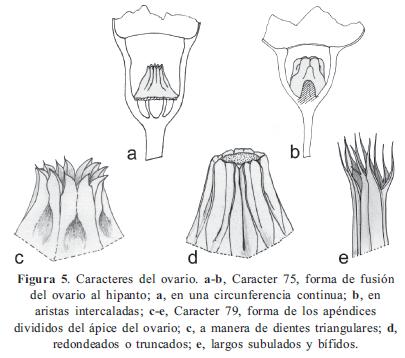
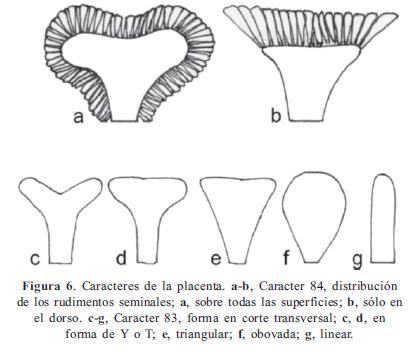
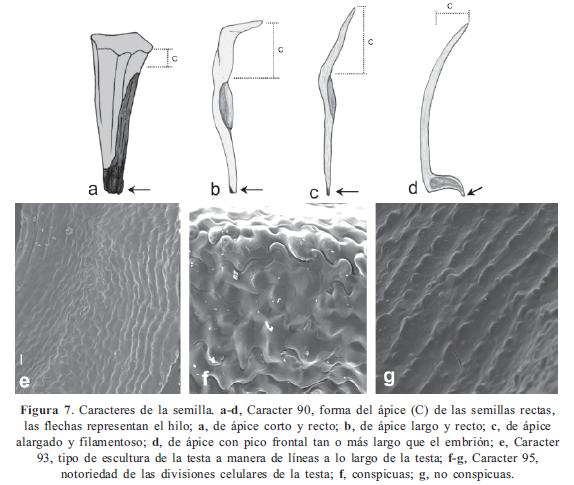
Los caracteres y sus estados, en parte, se basaron en los trabajos de Becquer (2008), Mendoza-Cifuentes & Fernández-Alonso (2010), Michelangeli (2000), Penneys (2007) y Schulman & Hyvönen (2003). Los caracteres de las semillas fueron basados parcialmente en Michelangeli (2000). En total se consideraron 103 caracteres, de los cuales 48 fueron doble-estado y los demás multiestado; ningún caracter fue considerado como aditivo. La terminología empleada se basó en Mendoza & Ramírez (2006) y Wurdack (1973, 1980), y para caracteres referentes al concepto de sinflorescencia se empleó la terminología propuesta por Mora-Osejo (1992). Los caracteres utilizados se presentan en el Apéndice 2 y en las Figuras 1, 2, 3, 4, 5, 6, 7 y 8; la lista de los caracteres excluidos se presenta en el Apéndice 3.

Análisis cladístico
La matriz de datos fue construida y editada con WinClada (Nixon, 1999). La filogenia fue realizada por medio del programa Nona y utilizando su interfase WinClada (Golobof, 1993). Se consideraron los siguientes parámetros de análisis: estrategia de búsqueda heurística de árboles más parsimoniosos con un número máximo de árboles retenidos de 10000, 1000 réplicas y 10 árboles por réplica de inicio; todos los caracteres fueron pesados igual; finalmente se obtuvo el árbol de Consenso Estricto. Se obtuvieron soportes de los clados usando el Índice de Soporte de Bremer (SB) por medio del programa Paup* versión 4.0b10 (Swofford, 2002), y de acuerdo al procedimiento propuesto por Hovekamp (2005).
Resultados y discusión
La matriz morfológica construida tuvo un total de 3,1% de caracteres perdidos por desconocimiento de estados de caracter, y 11,8% por estados inaplicables; el 3,6% de los datos representan polimorfismos (Apéndice 4). De la totalidad de caracteres, el 14,5% (15) fueron autopomórficos, el 44,7% (44) presentaron IR menor de 0,50 y el 44,7% (44) IR mayor o igual de 0,50. Con NONA se obtuvieron 356 árboles más parsimoniosos de 515 pasos incluyendo todos los caracteres y valores del IC de 0,33 y del IR de 0,58 (Farris, 1989).
Clados que consideran a Centronia
La Figura 9 muestra uno de los árboles más parsimoniosos y en donde se mapean los caracteres diagnóstico de Centronia. Se observa que el género se anida en siete clados distintos, lo que indica su artificialidad. La especie tipo de Centronia (C. laurifolia) se anida con C. reticulata en un clado junto con Graffenrieda rupestris, mientras que el resto de especies se encuentran asociadas en diferentes clados, algunas asociadas a los géneros Meriania y Wurdastom. El clado con mayor número de especies de Centronia se asocia a Meriania huilensis en una de las partes más derivadas del árbol.
Los caracteres diagnósticos de Centronia, como son la presencia de caliptra (36/3), la dehiscencia circuncísil de la caliptra (37/2) y el poro de la antera orientado ventralmente (53/0) (Triana, 1871; Wurdack, 1973), se encuentran en diferentes partes del cladograma indicando que son homoplásicos. Estos caracteres diagnósticos del género, ya sean considerados de forma individual o como un conjunto de caracteres, no constituyen una sinapomorfía de Centronia o de un subgrupo de sus especies (Fig. 9).
En el árbol de consenso estricto colapsaron 23 nodos y los valores del Índice de Soporte de Bremer (SB) fluctuaron entre 1 y 11 (Fig. 10). En este árbol Centronia se anida en siete diferentes clados confirmando su naturaleza polifilética mostrada en uno de los árboles más parsimoniosos. De acuerdo con esto, Centronia está conformado por los siguientes grupos: 1) C. laurifolia y C. reticulata en el clado I; 2) C. neblinae, sin una relación de parentesco definida con estos resultados; 3) C. vaupesana, sin una relación de parentesco definida con estos resultados; 4) C. ruizii, que se relaciona con el género Wurdastom en el clado k; 5) C. sessilifolia, que se asocia al género Meriania en el clado N; 6) C. peruviana, asociada con especies de Meriania en el clado O; 7) C. haemantha, C. insignis, C. mutabilis, C. mutisii, C. brachycera y C. dichromantha, asociadas al género Meriania en el clado Q (Fig. 10).
Clado I. Se puede considerarse como Centronia "sensu stricto" puesto que contiene a la especie tipo del género. No posee sinapomorfías y el conjunto de caracteres que lo soporta se asocian a la forma del pecíolo (10/0), las flores sésiles (29/1), el ovario levemente fusionado al hipanto (74/0), el ápice del ovario con apéndices fimbriados (79/2), y las semillas de ápice alargado (90/1); todos estos caracteres resultaron ser poco informativos. Este conjunto de dos especies, junto con C. neblinae y C. vaupesana, comparten con el género Graffenrieda caracteres como el poro de la antera orientado ventralmente (53/0), las anteras fuertemente arqueadas hacia el dorso (63/1), las placentas triangulares en corte transversal y con rudimentos seminales sólo en la superficie dorsal (83/1, 84/1), y las anteras con el haz vascular del conectivo dorsal no bifurcado (58/1). A pesar de estas semejanzas, el nivel de resolución del árbol de consenso estricto, no permite considerar estas especies como parte de Graffenrieda y su resolución demandaría un análisis con mayor muestreo. Este último género cuenta con 54 especies (Mendoza-Cifuentes & Fernández- Alonso, 2010) y sólo siete de ellas fueron consideradas en este trabajo.
Clado K. El clado K con el género Wurdastom y donde se encuentra C. ruizii, indica que esta última especie está mal ubicada genéricamente. Este clado está fuertemente soportado (SB = 10) y cuenta con tres caracteres sinapomórficos como son flores haplostemonas (50/1), ovario ínfero (74/1) y anteras oblongas (64/1) (Figs. 9 y 10). El primer caracter no es conocido en la tribu Merianieae, el segundo sólo se conoce en el género Merianthera Kuhlman y el último se encuentra sólo en algunas especies de Behuria Cham. y Bisglaziovia Cogn. (Mendoza-Cifuentes & Fernández- Alonso, 2010). Wurdastom es un género reciente y de posición tribal incierta (Wallnöfer, 1996), aunque los caracteres del ovario y el número de estambres permiten asociarlo al género Allomaeita Gleason de la tribu Cyphostileae (Lozano & Becerra, 1999b). Su anidamiento cerca de algunas especies de Graffenrieda se deba a efectos de muestreo, ya que este análisis no incluyó taxones de la tribu Cyphostileae. Centronia ruizii indudablemente debe ser transferida a Wurdastom y, en especial, ser confrontada con W. sneidernii (Wurdack) B.Wal., pues comparten muchos caracteres y la localidad tipo.
La totalidad de las especies restantes de Centronia se ubicaron a partir del clado N, en donde sólo se encuentran, adicionalmente, especies del género Meriania. Dentro de este grupo se encuentra el clado O, con C. peruviana, M. tomentosa y M. phlomoides el cual presentó el mayor soporte de todo el árbol de consenso estricto con un SB de 11. Este clado se caracteriza por las corolas tubulosas (42/0), inflorescencias con grandes forófilos en inflorescencias jóvenes (26/0), anteras seudoresupinadas y con poro apical (53/2, 62/0), y conectivo dorsal de forma irregular (98/1). Centronia peruviana fue descrita por Macbride (1941) en "Flora of Peru" y fue considerada dentro de Centronia por la presencia de la caliptra a pesar de que su dehiscencia es por fisuras irregulares. La especie comparte muchos caracteres con M. tomentosa a tal punto que puede ser considerada como una forma de ésta última y debe ser sinonimizada. El clado O se puede decir que representa un grupo muy particular dentro de Meriania por los caracteres de la corola tubulosa (40/2) y las anteras seudoresupinadas (53/2). Este grupo está conformado por unas 18 especies que crecen desde Bolivia hasta Venezuela y Costa Rica, a lo lago de los Andes y el Chocó biogeográfico, y también por algunas especies que crecen en las Antillas, como M. involucrata (Desr.) Naudin. Dentro de Meriania, este grupo es plenamente reconocible con el anterior conjunto de caracteres.
Clado Q. El clado Q, es el mayor grupo constitutivo de Centronia con cinco especies. Este grupo de Centronia se caracteriza por los estambres isomórficos (51/0), las caliptras de ápice agudo (38/1-2), cubierta del hipantocaliptra de tricomas ferrugíneos pinoide-deltoides (100/1), y cambio de coloración de los pétalos de rojo púrpura a violeta (47/0). De acuerdo con estos resultados, este conjunto de especies puede ser transferido a Meriania y es posible considerarlo como un grupo natural dentro de este género ya que el anterior conjunto de caracteres no se encuentra en otras especies de Meriania (Mendoza-Cifuentes & Fernández-Alonso, 2012).
Clado N. Finalmente, la especie C. sessilifolia, presente en el clado N, tiene caracteres que permiten transferirla a Meriania como son el tamaño de los pétalos y la forma de los estambres (caracteres 30/2, 57/1, 58/0). Esta especie comparte caracteres particulares con M. maguirei como son las caliptras de ápice redondeado (38/0), presencia de dientes dorsales del cáliz (39/1) e inflorescencias reducidas (20/0).
Clados basales (Merianieae)
En el árbol de consenso estricto se diferencian dos grupos en la tribu Merianieae: el de Phainantha, Huberia y Pachyloma (clado B) con valor del SB de 6, y el grupo de Graffenrieda, Meriania, Axinaea, Adelobotrys y Centronia (clado C), con valor del SB de 1 (Fig. 10). El clado B presentó estambres geniculados, semillas aladas o cocleadas y placentas lineares excepto por Pachyloma. El clado C presentó dos sinapomorfías que son los estambres no geniculados y las anteras rectas o curvadas hacia el dorso y no hacia el vientre; este clado también se caracterizó por las semillas cuneiformes o filamentosas. Resultados similares documentan Schulman & Hyvönen (2003), donde separan a "Core Merianieae" de otros géneros entre los que se encuentran Pachyloma, Behuria y Huberia (PBH Clade). Se puede decir que en Merianieae existen dos grupos de géneros: los de estambres geniculados y los de estambres no geniculados (Mendoza-Cifuentes & Fernández-Alonso, 2010). Este último grupo constituye el clado C de este trabajo y comprende cinco de los 15 géneros que constituyen la tribu; no obstante, estos cinco géneros poseen el 80% de las especies de Merianieae (Mendoza-Cifuentes & Fernández-Alonso, 2010).
Clados relacionados con Graffenrieda y Meriania
Considerando que la mayor parte de las especies de Centronia se asocian ya sea a Graffenrieda o a Meriania, es importante comentar aspectos de las diferencias entre estos dos géneros de acuerdo con los resultados de este análisis cladístico.
Grafenrieda. Los subclados basales dentro del clado C involucraron principalmente a especies del género Graffenrieda (clados E, H, J). El nivel de resolución del árbol de consenso estricto no permite establecer mayor comentario sobre este género, no obstante, Graffenrieda tiene caracteres estables del cáliz y los estambres (Mendoza-Cifuentes & Fernández-Alonso, 2010), así como del ovario y de los pétalos, que permiten separarlo de otros géneros de Merianieae. Entre estos caracteres se encuentran: el poro de la antera orientado ventral o apicalmente (53/0-2); las anteras arqueadas hacia el dorso (63/1), placentas triangulares en corte transversal y con rudimentos seminales sólo en la superficie dorsal (83/1, 84/1), y estambres con un haz vascular del conectivo dorsal no bifurcado (58/1). El anterior conjunto de caracteres, junto con los pétalos diminutos a medianos y de coloración blanca o rosada, sólo se encuentra en Graffenrieda y permite diferenciarlo principalmente de Meriania. Como se comenta párrafos arriba, cuatro especies de Centronia comparten algunos de estos caracteres con Graffenrieda, lo que sugiere su posible transferencia en este último género, no obstante, el árbol de consenso estricto no muestra suficiente resolución al respecto y es necesario un análisis con mayor muestro.
Meriania. Por otro lado, el género Meriania se muestra más derivado en el árbol de consenso estricto con respecto a Graffenrieda, y se encuentra a partir del clado L (Fig. 10). Este clado presenta SB de 2 y 11 caracteres de soporte, dos de ellos sinapomórficos: dimorfismo estaminal (51/2), y anteras con poros dorsales (53/1). Los bajos valores del SB, el anidamiento de especies de los géneros Adelobotrys, Axinaea y Centronia dentro de este clado, y la gran variabilidad en los caracteres de los estambres y el cáliz (Mendoza-Cifuentes & Fernández-Alonso, 2010) permite sugerir que Meriania no es un grupo natural. Considerando futuros arreglos o depuraciones de este género, algunos de los caracteres importantes que pueden sustentar a Meriania son: anteras con poros dorsales (53/1) y pétalos de tamaño grande a gigante (30/2-3). El caracter de la placenta con rudimentos seminales en todas las superficies (84/ 0) fue estable en todas las especies de este género en este análisis, pero aun falta mayor evaluación en otras especies.
Aspectos biogeográficos
Las consideraciones biogeográficas que pueden deducirse del árbol de consenso estricto no son muy amplias dado el grado de resolución dentro del clado C. En la parte basal de este clado son coincidentes grupos del Escudo Guayanés y zonas de los Andes asociadas a la Cordillera del Cóndor en los límites entre Ecuador y Perú. En la parte más derivada se encuentran grupos de los Andes del norte (Fig. 10). Todas las especies de Centronia no asociadas a Meriania se encuentran en el Escudo Guayanés y Cordillera del Cóndor, mientras que las especies que se asocian a Meriania se restringen a los Andes excepto por C. ruizii. Esta asociación geográfica respalda aun más los distintos orígenes de grupos de especies de Centronia y su artificialidad taxonómica.
El grupo con mayor número de especies de Centronia, clado Q, se concentra en el norte de los Andes, en especial en la Cordillera Oriental de Colombia y su continuación en Venezuela entre los 1200 m hasta zonas de páramo. En Ecuador este grupo solo está representado en el norte, cerca de la frontera con Colombia, y vuelve a aparecer en el norte de Perú (C. haemantha). Esta asociación geográfica confirma la necesidad de transferir las especies de Centronia de los Andes del norte al género Meriania (Mendoza- Cifuentes & Fernández-Alonso, 2012).
El clado K, en donde se encuentra Centronia ruizii y especies de Wurdastom, se encuentra en la planicie del Chocó biogeográfico y a lo largo del piedemonte occidental de los Andes hasta los 2000 m de altitud. Como se discute arriba, su presencia dentro del clado C obedece a efectos de muestreo, por lo que no se puede establecer alguna relación biogeográfica con estos resultados.
Variación y evolución de caracteres
A continuación se discuten algunos de los caracteres con mayores valores del IR y que pueden ser importantes taxonómicamente dentro del clado C de Merianieae. También se consideran algunos caracteres ampliamente utilizados dentro de la tribu.
Entre los caracteres vegetativos es importante resaltar la nerviación foliar. Los caracteres 14 y 15, con valores del IR de 0,54 y 0,72 respectivamente, mostraron correspondencia geográfica y taxonómica. La nerviación con sólo un par de venas secundarias y cercanas a la margen (broquidódroma) fue constante en todas las especies de Graffenrieda no andinas consideradas en el análisis, lo que indica que puede ser un caracter útil dentro de este género. En Meriania el carácter 14 es variable pero predomina la nerviación con varios pares de venas secundarias.
Caracteres del cáliz han sido utilizados clásicamente para diferenciar géneros y establecer secciones dentro de géneros en Merianieae (Mendoza-Cifuentes & Fernández- Alonso, 2010). Esta estructura es bastante plástica en la tribu y algunos de sus caracteres resultaron ser homoplásicos en este análisis. Los caracteres 36 al 40 corresponde al cáliz y sus valores del IR fluctuaron entre 0,0 y 0,32, excepto por el caracter 37 (forma de apertura de las caliptras). Este último caracter muestra ser plesiomórfico en su estado "apertura por fisuras irregulares". El caracter "presencia de caliptra" ha emergido varias veces en Merianieae (clado C) y no es útil para establecer diferencias genéricas en la tribu. Este caracter se encuentra en la tribu Cyphostileae (Lozano & Becerra, 1999b) y en varios géneros de Miconieae, Blakeeae y Henrietteeae (Mendoza & Ramírez, 2006; Penneys, 2007; Mendoza-Cifuentes & Fernández-Alonso, 2010; Penneys et al., 2010), lo que muestra que también es homoplásico al nivel de la familia.
El tamaño de los pétalos (30) mostró un IR de 0,64 y es un subrogado del tamaño de la flor. De acuerdo con los resultados, los pétalos grandes a gigantes es el estado derivado y caracteriza al grupo netamente andino de Meriania (clado O). Los pétalos de tamaño pequeño a mediano predominaron en las especies de Graffenrieda. Otro caracter de los pétalos es el cambio de coloración de rojo púrpura a violeta (47), con un IR de 1, siendo este cambio un caracter derivado que caracteriza al clado Q. El tipo de corola (42) también presentó un valor alto del IR y los resultados indican que las corolas tubulosas son un caracter derivado y en Meriania ha emergido en dos grupos distintos: en el grupo de Meriania tomentosa y en el grupo de especies asociado a Meriania peltata.
De caracteres de los estambres es importante resaltar la orientación del poro de la antera en estambres opositipétalos (53), con un IR de 1. En Graffenrieda este caracter varía en especies asociadas a los Andes, como G. cucullata, con poros apicales, pero en el resto de especies siempre es ventral. En Meriania, Adelobotrys y Axinaea, la orientación del poro es dorsal excepto por un pequeño grupo en Meriania con poros apicales (clado O). El caracter 55 es igualmente importante pues permite separar dos grandes grupos dentro de Merianieae. Este caracter es invariable dentro del clado C, donde se encuentran estambres no geniculados (55/0); no obstante, este estado de caracter también se encuentra en algunas especies de Blakea de la tribu Blakeeae (Penneys, 2007).
Los caracteres asociados a la placenta han sido poco utilizados en Melastomataceae, no obstante, trabajos como el de Schulman & Hyvönen (2003) y de Becquer (2008) documentan que poseen valor filogenético. Dentro de Merianieae clado C, los caracteres 83 y 84 de forma de la placenta y distribución de los rudimentos seminales, respectivamente, presentaron altos valores del IR y mostraron ser congruentes. El caracter 84 muestra una clara diferencia entre taxones como Graffenrieda y los géneros Meriania, Adelobotrys y Axinaea. De acuerdo con estos resultados las placentas con rudimentos sólo en dorso corresponde al estado plesiomórfico dentro del clado C.
La forma de la semilla también es otro caracter importante para establecer el clado C de Merianieae. Sin embargo, dentro de este clado los caracteres como la forma del ápice o "cabeza" de la semilla y de la testa presentaron bajos valores del IR y variaciones dentro de subclados, que indican cierto nivel de plasticidad. Por ejemplo, la cabeza de las semillas en Adelobotrys y Meriania es variable entre cabeza alargada y cabeza corta.
Conclusiones
La evaluación filogenética de Centronia con base en caracteres morfológicos indica que el género es polifilético. Las especies del género se anidaron en siete lugares distintos del árbol de consenso estricto. De acuerdo con esto Centronia s. str. se reduce a C. laurifolia y C. reticulata; una especie debe ser transferida al género Wurdastom (C. ruizii), y 10 especies deben transferirse al géneroMeriania (C. sessilifolia, C. peruviana, C. haemantha, C. insignis, C. mutabilis, C. mutisii, C. brachycera, C. dichromantha, C. pulchra y C. eximia). Las especies C. neblinae y C. vaupesana se consideran de posición genérica incierta y para su plena resolución filogenética es necesario realizar un análisis cladístico completo del género Graffenrieda.
Centronia s. str. no posee caracteres sinapomórficos y el conjunto de caracteres que lo sustenta también se encuentra en Graffenrieda. No obstante, el nivel de resolución del árbol de consenso estricto no permite concluir aun sobre su inmersión o fusión con Graffenrieda.
Agradecimientos
Este proyecto (Núm. 201010011273) contó con la financiación de la Dirección de Investigación (DIB) de la Universidad Nacional de Colombia, sede Bogotá a quien expresamos nuestro agradecimiento. Del mismo modo, el segundo autor agradece al Ministerio de Ciencia e Investigación de España, el apoyo del Proyecto CGL 2010-19747 (Subprograma BOS) para la finalización de estre trabajo. Nuestros sinceros agradecimientos a las siguientes instituciones por la ayuda prestada para el desarrollo de este trabajo: Instituto de Ciencias Naturales de la Universidad Nacional de Colombia; Instituto Alexander von Humboldt; Field Museum de Chicago, Missouri Botanical Garden, New York Botanical Garden, Instituto Smithsonian; los herbarios COL, COAH, FMB, CUVC, HUQ, CAUP, QCA, QCNE, MO, F, NY, UPTC, US. A los colegas Fabian Michelangeli, Carmen Ulloa, Corine Vriecendrop, Tyana Wachter, Robin Foster, William Ariza, Gorky Villa, Wilson Quishpe y Janet Muñoz quienes contribuyeron de forma variada, suministrando imágenes, ejemplares o facilitando el apoyo logístico. A Fernando Alzate, Fabio González, Carlos Parra y a dos evaluadores anónimos por sus valiosas observaciones al manuscrito.
Referencias bibliográficas
Considera las referencias citadas en el Apéndice 2
Almeda, F. 1993. An evaluation of the mesoamerican species of Meriania (Melastomataceae: Merianieae). Proceedings of the California Academy of Sciences 48(7):141-152. [ Links ]
Becquer, E. R. 2008. Taxonomía y filogenia del género Pachyanthus (Melastomataceae: Miconieae). Universidad de la Habana, Jardín Botánico Nacional, Cuba. Tesis de Doctorado. [ Links ]
Clausing, G. & S. S. Renner. 2001. Molecular phylogenetics of Melastomataceae and Memecylaceae: implications for character evolution. American Journal of Botany 88(3):486-498. [ Links ]
Cogniaux, A. 1891. Melastomatacées. (p. 425). En: de Candolle (ed.), Monographie Phanerogamarum 7. Paris. [ Links ]
Don, D. 1823. An Ilustration of the natural family of plants called Melastomataceae. Memoirs of the Wernerian Natural History Society. Edinburgh. [ Links ]
Farris, J. S. 1989. The retention index and the rescaled consistency index. Cladistics 5:417-419. [ Links ]
Goloboff, P. 1993. Nona. Versión 2.0. Software and documentation by the author. Tucuman, Argentina. [ Links ]
Hovenkamp, P. 2005. Branch Support. Nationaal Herbarium Nederland, Leiden University. Leiden (Fecha de consulta: 5/ Jun/2009. http://www.nationaalherbarium.nl/taskforcemo lecular/ PDF/Branch%20suppoerts.pdf). [ Links ]
Lozano, G. & N. Becerra. 1999a. Notas sobre el género Centronia (Melastomataceae) en Colombia. Revista de la Academia Colombiana de Ciencias 23:79-83. [ Links ]
Lozano, G. & N. Becerra. 1999b. Los géneros Allomaieta y Cyphostyla (Melastomataceae). Revista de la Academia Colombiana de Ciencias 23(86):5-18. [ Links ]
Macbride, J. F. 1941. Melastomataceae - Flora of Peru. Publications of the Field Museum of Natural History. Botanical series 13, pt. 4:327. [ Links ]
Mendoza-Cifuentes, H. & B. Ramírez. 2006. Guía ilustrada de géneros de Melastomataceae y Memecylaceae de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt; Universidad del Cauca, Bogotá D.C. 288 p. [ Links ]
______ & J. L. Fernández-Alonso. 2010. Evaluación de caracteres del cáliz y los estambres en la tribu Merianieae (Melastomataceae) y definición de homologías. Revista de la Academia Colombiana de Ciencias 34(131): 143-172. [ Links ]
______ & J. L. Fernández-Alonso. 2012. Novedades en Centronia y Meriana (Merianieae, Melastomataceae) y revisión taxonómica del "Grupo Brachycera". Anales del Jardín Botánico de Madrid -Entregado-. [ Links ]
Michelangeli, F. 2000. A cladistic analysis of the genus Tococa (Melastomataceae) based on morphological data. Systematic Botany 25(2):211-234. [ Links ]
______ ., D. S. Penneys, J. Giza, D. Soltis, M. H. Hils & J. D. Skean Jr. 2004. A preliminary phylogeny of the tribe Miconieae (Melastomataceae) based on nrITS sequence data and its implications on inflorescence position. Taxon 53(2):279-290. [ Links ]
Mora-Osejo, L. E. 1992. La evolución de la reproducción en la cormobionta y de las unidades de crecimiento y floración de las Angiospermae. Revista de la Academia Colombiana de Ciencias 18(70):311-322. [ Links ]
Nixon, K. C. 1999. WinClada. Version 1.00.08. Software and documentation distributed by the author. Cornell University, Ithaca, N.Y. [ Links ]
Peneys, D. S. 2007. Phylogeny and character evolution in the Blakeeae (Melastomataceae). Tesis de Doctorado, University of Florida. 176 p. [ Links ]
______ , F.A. Michelangeli, W.S. Judd & F. Almeda. 2010. Henrietteeae (Melastomataceae): A new neotropical berry-fruit tibe. Systematic Botany 35(4):783-800. [ Links ]
Renner, S. S. 1993. Phylogeny and classification of the Melastomataceae and Memecylaceae. Nordic Journal of Botany 13(5):519-550. [ Links ]
Schulman, L. & J. Hyvönen. 2003. A cladistic analysis of Adelobotrys (Melastomataceae) based on morphology, with notes on generic limits whitin the Tribe Merianieae. Systematic Botany 28(4):738-756. [ Links ]
Swofford, D. L. 2002. Paup* Phylogenetic Analysis Using Parsimony (*and other methods). Versión 4.0b10 para Macintosh. Sinuaer, Sunderland. [ Links ]
Triana, J. 1871. Les Melastomacées. Transactions of the Linnean Society of London 28:1-188. [ Links ]
Varassin, I. G., D. S. Penneys & F. A. Michelangeli. 2008. Comparative anatomy and morphology of nectar-producing Melastomataceae. Annals of Botany 102:899-909. [ Links ]
Wallnöfer, B. 1996. A revision of the genus Alloneuron Pilg. and segregation of Wurdastom gen. n. (Melastomataceae). Annalen Naturhistorischen Museums in Wien 98B Supplement: 447-462. [ Links ]
Wurdack, J. J. 1973. Melastomataceae (Memecyleae by T. Morley). P 1-819. En: T. Lasser (ed.). Flora de Venezuela. No. 8. Instituto Botánico, Ministerio de Agricultura y Cría, Caracas. [ Links ]
______ . 1980. Melastomataceae. En: G. Harling & B. Sparre (eds.). Flora of Ecuador. No. 13. Univ. Göteborg & Riksmuseum, Stockholm. 406 p. [ Links ]
Recibido: abril 13 de 2011. Aceptado para su publicación: diciembre 2 de 2011.
