










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
Print version ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.36 no.139 Bogotá Apr./June 2012
GEOLOGÍA
* Servicio Geológico Colombiano, Subdirección de Geología Básica, Bogotá. Email: mpardo@ingeominas.gov.co
RESUMEN
En 1994 se realizó el hallazgo de un cráneo de Mastodonte juvenil en zona rural del Municipio de Turbaná. El presente reporte describe el ejemplar y expone de manera resumida el criterio de los diferentes autores acerca de la nomenclatura y posición sistemática de los mastodontes suramericanos. La pieza es asignada provisionalmente a la especie Stegomastodon waringi (Holland, 1920), hasta que un pronunciamiento oficial de la Comisión Internacional de Nomenclatura Zoológica resuelva y aclare la validez de los criterios taxonómicos y la prevalencia de los diferentes nombres usados a través del tiempo para clasificar los restos de este grupo de la megafauna Suramericana.
Palabras clave: Mastodonte, Stegomastodon, Haplomastodon, taxonomía, megafauna.
ABSTRACT
In the year 1994 was the discovery of a juvenile mastodon skull in the rural area of Turbaná town. This report describes the sample and briefly summarized the approach of different authors concerning the nomenclature and systematic position of the South American mastodons. The specimen is tentatively assigned to the specie Stegomastodon waringi (Holland, 1920), until an official statement from the International Commission of Zoological Nomenclature resolve and clarify the validity of the taxonomic criteria and the prevalence of the different names used over time to classify the remains of this group of South American megafauna.
Key words: Mastodons, Stegomastodon, Haplomastodon, taxonomic, megafauna.
Introducción
En el primer semestre de 1994 al sur de la ciudad de Cartagena de Indias se realizó de manera fortuita el hallazgo de un cráneo de mastodonte juvenil en sedimentitas de la Unidad Detrítica de La Popa. En ese mismo año El Museo Geológico Nacional José Royo y Gómez del Servicio Geológico Colombiano (antes INGEOMINAS), adelantó la prospección del hallazgo Paleontológico encabezado por la Dra. María Páramo Fonseca, dichas labores permitieron confirmar que los restos fósiles correspondían a un fragmento craneal de mastodonte juvenil denominados hasta entonces restos de Mastodonte de Turbaná (preliminarmente UC-1 o IPN-43). En ese momento el INGEOMINAS indicó la relevancia del hallazgo y solicitó a los propietarios del predio la donación de la pieza a una entidad museística regional; así en el año 2004 el señor álvaro Durango donó formalmente a la Universidad de Cartagena la pieza paleontológica, la cual fue albergada y cuidada en el Claustro de San Agustín de la Universidad de Cartagena, bajo la custodia del profesor Wilson Blanco, profesor titular del programa de Historia de la Universidad de Cartagena. Posteriormente la pieza fue examinada en el año 2005 por el paleontólogo Carlos Villarroel y el Arqueólogo Gonzalo Correal, quienes en su momento y con base en las observaciones realizadas clasificaron la pieza dentro del género Haplomastodon. En Octubre de 2009 se suscribió formalmente el convenio interadministrativo celebrado entre la Universidad de Cartagena y el Museo Geológico Nacional del Instituto Colombiano de Geología y Minería INGEOMINAS, en el marco del cual se trasladaron los restos fósiles a los laboratorios del Museo en la ciudad de Bogotá, con el fin de realizar las respectivas labores de curatoría y preparación de las piezas para su adecuada conservación y posterior exhibición en los predios de la Universidad de Cartagena. La preparación de los restos estuvo a cargo del Señor Gerardo Vargas Quintero, técnico del laboratorio 1 de macrofósiles del Museo Geológico Nacional de INGEOMINAS y de Mauricio Pardo Jaramillo, geólogo-paleontólogo de la misma institución y autor del presente reporte.
La historia de la taxonomía o clasificación de los mastodontes Sur Americanos es bastante compleja ya que esta ha sufrido constantes revisiones y clasificaciones, aun cuando los criterios para su determinación han sido básicamente los mismos, como lo son la forma y presencia o no de banda de esmalte en las defensas, la morfología y manera de desgaste en los molares y la forma general del cráneo. Resultado de estas variadas maneras de entender la taxonomía de este grupo fósil son las diferentes corrientes que existen en la actualidad para clasificar los proboscídeos suramericanos, sin que hasta el momento la Comisión Internacional de Nomenclatura Zoológica se haya pronunciado oficialmente al respecto.
Los mastodontes suramericanos son bunodontos, es decir que sus molares poseen montículos separados y redondeados para triturar sus alimentos. Estos montículos presentan una compleja estructura que consta de un patrón seriado de hileras transversales o lofos. Cada lofo posee dos cúspides principales con forma de cúpula (conos o colinas) y cada una de estas cúpulas puede o no exhibir de manera adyacente cúspides más pequeñas (cónulos o conoletes) lo cual origina diferentes figuras en la superficie oclusal del molar a medida que este se desgasta.
Cabrera (1929) reconoce 3 géneros de mastodontes para Argentina: Cuvieronius OSBORN, 1923, Notiomastodon CABRERA, 1929 y Stegomastodon POHLIG, 1912, diferenciando Notiomastodon de Stegomastodon por tan solo la persistencia de una banda lateral de esmalte sin torsión en las defensas de Notiomastodon, rasgo que es reconocido por autores posteriores en individuos juveniles de Haplomastodon HOFFSTETTER, 1950 y Stegomastodon (Prado et al., 2005., Alberdi y Corona, 2005., Ferretti, 2008.). Posteriormente Hoffstetter (1950) describe Haplomastodon basado en material de Ecuador, considerándolo un subgénero de Stegomastodon, luego, en 1952 el mismo autor elevó este rango a género, basándose en la ausencia de un foramen transversal en el atlas, carácter que luego sería desvirtuado por otros autores (Simpson y de Paula Couto, 1957; Ficcarelli et al., 1993, 1995). Así, Hoffstetter considera válidos los géneros Cuvieronius, Stegomastodon y Haplomastodon y estima que Notiomastodon es en realidad una variación de Stegomastodon. Simpson y de Paula Couto (1957) reconocen la presencia en Sur América de los géneros Cuvieronius, Stegomastodon y Haplomastodon y guardan sus reservas en cuanto al género Notiomastodon, al que consideran, al igual que Hoffstetter, una variación de Stegomastodon. Con respecto a las diferencias entre Haplomastodon y Stegomastodon afirman, que la estructura de los molares en Stegomastodon es más compleja, sin embargo, la alternancia entre pretritos (cúspide con cónulos accesorios) y postritos (cúspides sin cónulos accesorios) en cada lofo de los molares es más marcada en estos dos géneros que en Cuvieronius; siendo los molares de Stegomastodon menos bunodontos que los de Haplomastodon y Cuvieronius. Ficcarelli et al. (1993, 1995) consideran válidos para Sur américa tres géneros de mastodontes: Cuvieronius, Haplomastodon, representado por la especie H. waringi (HOLLAND, 1920) y Stegomastodon, representado por la especie S. platensis (AMEGHINO, 1888). Alberdi y Prado (1995), proponen un esquema simplificado, incluyendo Haplomastodon y Notiomastodon dentro del género Stegomastodon, reduciendo a dos el número de géneros reportados en Sur América con tan solo 3 especies: Cuvieronius hyodon (FISCHER, 1814), Stegomastodon waringi y Stegomastodon platensis, consideran pues, que las diferencias asignadas entre los restos de Stegomastodon y Haplomastodon son tan reducidas que no se deben considerar genéricas. Ferretti (2008) hace una completa revisión de la historia de la sistemática de los proboscídeos suramericanos, aceptando en cuanto a los géneros, la revisión que en su momento hicieron Simpson y Paula Couto (1957), no sin anotar que esta nomenclatura sigue en controversia. Ferretti (2009, 2010) propone designar Haplomastodon chimborazi (PROAÑO,1922) de Ecuador nombre válido y Neotipo de H. waringi, argumentando diversas razones que son rebatidas por Pasenco (2010), Lucas (2010) y Demirjian (2010) en sendas publicaciones del Boletín de Nomenclatura Zoológica.
Lucas (2009, 2010) aconseja conservar el uso del nombre Mastodon waringi HOLLAND, 1920, tal como fue establecido en su momento, pero designando un Neotipo, ya que el holotipo de Pedra Vermelha (Brasil) consta de material no diagnóstico. Mothé, Avilla y Winck (2010), estudian varios aspectos de los Gonfoterios del rico yacimiento de Aguas de Araxá en Brasil y con respecto a la nomenclatura acogen el uso del nombre Stegomastodon waringi para clasificar los mastodontes del mismo yacimiento que en 1957 Simpson y de Paula Couto catalogaron como Haplomastodon waringi. En estudios paleoecológicos publicados por Dominato, Mothé, Costa da Silva y Avilla (2011) sobre los mismos restos de Aguas de Araxá, usan el nombre Haplomastodon waringi, sin embargo un poco después Asevedo, Winck, Mothé y Avilla (2011) hacen un estudio con los restos de este mismo yacimiento (Aguas do Araxá) acerca de la paleo-dieta de estos animales, pero esta vez los clasifican, posiblemente siguiendo en parte la propuesta de Madden (1984), como Notiomastodon platensis, respaldando esta última opción está en prensa el trabajo de Mothé et al (2011) en dónde se revisan las colecciones de los principales museos del Cono Sur y proponen el uso de este nuevo nombre para denominar los mastodontes Suramericanos diferentes de Cuvieronius hyodon, añadiendo una nueva forma de ver la intrincada historia taxonómica del grupo.
Materiales y Métodos
El cráneo examinado en el presente trabajo ( IGM p881179) será resguardado en calidad de comodato en la Universidad de Cartagena, ( Bolívar ), en un lugar especial dispuesto por la facultad de Ciencias Humanas de dicha Institución; por su parte la Curatoría y mantenimiento de la pieza será responsabilidad del Museo Geológico Nacional 'José Royo y Gómez'.
Se ha seguido la propuesta de Alberdi et al. (2002) para la descripción general de los molares y el registro de los dadatos morfológicos y morfométricos, ya que este es el trabajo que mejor ilustra con dibujos y esquemas las características dentales y craneales a medir. Para el registro de estos datos se utilizaron los instrumentos usuales para dicha tarea: un calibrador digital de 6 pulgadas y una cinta métrica.
Las características de la pieza de estudio indican que se trata de un individuo juvenil, por esta razón las medidas tomadas tienen primordialmente la finalidad de describir el ejemplar, no sobra anotar que dichos datos deben ser tratados con sumo cuidado a la hora de hacer análisis estadísticos comparativos con otros restos craneales de Gonfoterios, pues el individuo exhibe molares deciduales y como es de suponer no ha completado su crecimiento.

Repositorio: El espécimen estudiado reposará en el Claustro de San Agustín de La Universidad de Cartagena (Bolívar), haciendo parte del Inventario Paleontológico Nacional del Museo Geológico Nacional 'José Royo y Gómez' De INGEOMINAS; bajo el número de catálogo IGM p881179.
Descripción
El material estudiado consta de un cráneo de un individuo juvenil, con dos molares deciduos trilofodontos (dM2 y dM3), izquierdos y derechos respectivamente y los primeros molares definitivos (M) completamente formados e incluidos en el maxilar. El cráneo sufrió una fractura dorsiventral en la parte media del lado derecho del maxilar, que distorsiona y "adelanta" aparentemente la posición del dM3 derecho, esta fractura desplaza ligeramente hacia la izquierda todo el premaxilar; así mismo la parte distal del arco zigomático derecho (proceso zigomático temporal) esta fracturada y desplazada haciendo difícil reconocer la fosa temporal; evidencias de dicho fracturamiento son los numerosos fragmentos óseos sueltos que se encuentran en la matriz, así como el resquebrajamiento del material óseo. De la arcada yugal (proceso zigomático del maxilar) solo se conservan las uniones de éstas con el maxilar, preservándose un área un poco mayor de la arcada yugal izquierda. El premaxilar exhibe el alveolo derecho de la defensa bastante deteriorado mientras que el alveolo izquierdo conserva la raíz de la misma. La orientación de los dos lados del premaxilar (y la parte más anterior del maxilar) junto con los alveolos de las defensas es casi paralela. La incisura dental es estrecha.
Molares (Tabla 1)
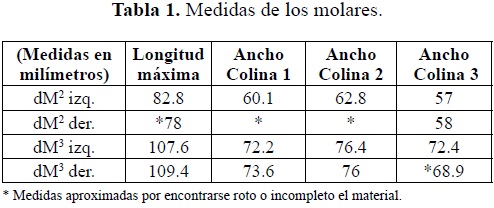
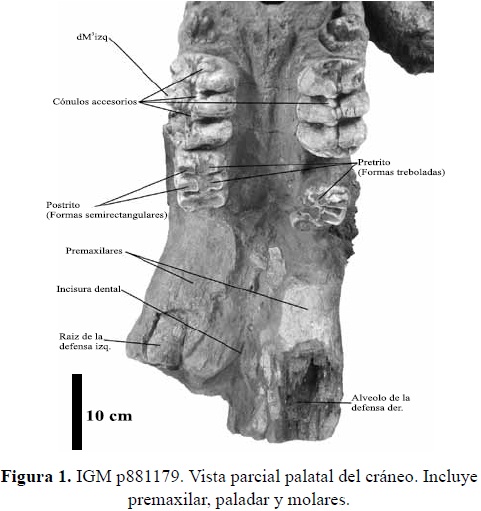
Tercer Molar superior deciduo izquierdo (dM3 izq.): En el lofo 1 la colina del lado bucal se encuentra rota completamente; posee un cónulo accesorio bilobulado en la parte mesial. El lofo 2 se encuentra completo, en la colina lingual se aprecian 3 cónulos accesorios dispuestos hacia la línea media del molar. El lofo 3 tiene la colina bucal rota en la parte posterior, faltando toda la corona hasta su base. Segundo molar superior deciduo derecho (dM2 der): En el lofo 1 falta todo el lado bucal (postrito). En el lofo 2 la colina lingual se encuentra rota (faltante) no pudiéndose apreciar su morfología. En el lofo 3 la cumbre de la colina bucal está desportillada.
Tercer molar superior deciduo derecho (dM3 der): El primer lofo presenta dos coneletes muy bajos y anchos en la parte delantera del molar. Los lofos 1 y 2 están prácticamente completos y sin desgaste, muy bien preservados, excepto que en la colina bucal del segundo lofo, muy cerca de su parte más alta, falta una pequeña porción del esmalte. Al lofo 3 le falta media colina del lado bucal, posee un cónulo accesorio posterior roto así como también falta la parte posterior lingual del talón de este molar.
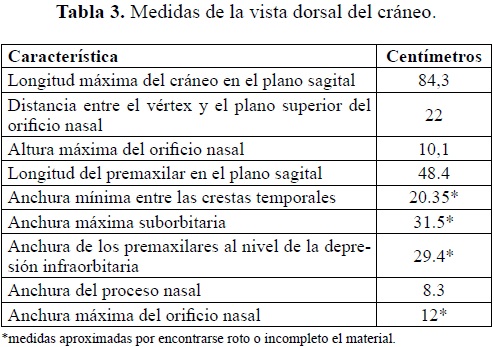
Vista ventral (Tabla 2)
El basicráneo en general incluyendo el esfenoides (Figuras 1 y 2) y la región occipital (Figura 3) se encuentran bien preservados. Palatino estrecho con quilla central suave, la cual se empieza a bifurcar 7,8 cm antes de la fosa de las coanas. Del pterigoides derecho e izquierdo solo se preservo la base la cual constituye a su vez los bordes laterales de la fosa de las coanas. El cóndilo occipital y la región occipital están bien preservados, en ésta última se aprecian claramente las fosas y la cresta del ligamento nucal.
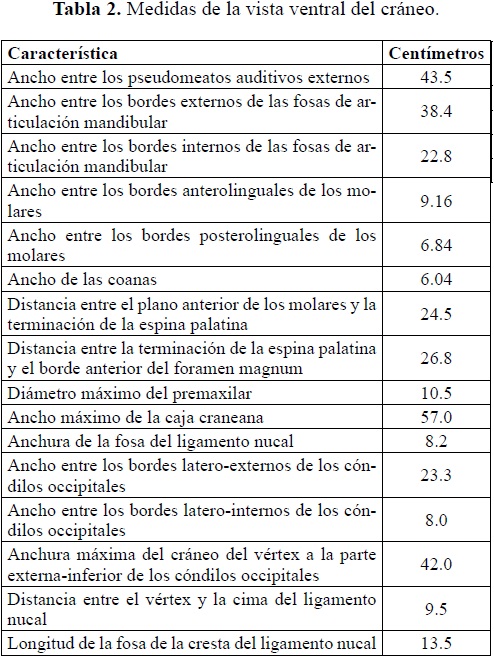
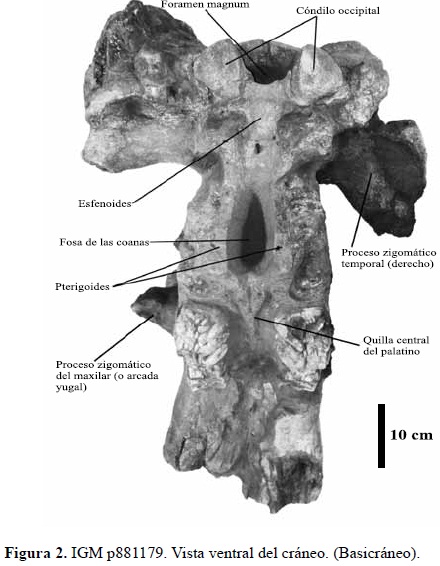

En general la superficie del cráneo en esta vista no está bien preservada, en varios lugares del cráneo se presentan fragmentos óseos no articulados dentro de una matriz limoarenosa, dicha situación dificultó la preparación de este lado de la pieza. La parte antero-superior del premaxilar derecho exhibe una fractura con desplazamiento de la parte bucal superior que da a esta parte del premaxilar un aspecto abultado, esta estructura está constituida en parte por hueso esponjoso, sin embargo todo el "promontorio" no corresponde a material óseo sino a sedimento areno-limoso que sirve de soporte a las partes de hueso desplazadas; dicha fractura continúa hacia la parte posterior y atraviesa de forma diagonal todo el premaxilar derecho.
La superficie de la parte posterior del premaxilar derecho no se preservó; debido a que justo en esa parte ya el hueso exhibe una compleja estructura neumática muy evidente en la parte lateral y posterior del premaxilar, lo cual influye en la preservación, pues los espacios vacíos en estas estructuras no permiten que los delgados huesos presentes soporten el peso del sedimento y los demás huesos craneales. La parte anterior de maxilar derecho se encuentra fracturada, es este sector se aprecian varios fragmentos no articulados soportados por matriz limoarenosa.
No se preservó tampoco parte alguna del material óseo del lado izquierdo del orificio nasal y del frontal, este último se encuentra fracturado sin desplazamiento y esta soportado por la matriz sedimentaria limoarenosa; la situación es diferente si se avanza hacia la parte distal o posterior del cráneo en dónde el parietal se encuentra muy fracturado, aquí los fragmentos de este hueso se encuentran desplazados entre si y están soportados al igual que el frontal por la matriz del interior del cráneo.
Discusión y Conclusiones
La propuesta de Lucas (2009, 2010), en opinión del autor de este trabajo es muy viable, en ella aconseja conservar el uso del nombre Mastodon waringi HOLLAND, 1920, tal como fue establecido en su momento, pero designando un Neotipo, ya que el holotipo de Pedra Vermelha (Brasil) consta de material no diagnóstico. Esta última propuesta sería igualmente aplicable en el caso de que se siguiera la opinión de Alberdi y Prado (1995) que considera Haplomastodon como sinónimo junior de Stegomastodon, pues la especie waringi como se mencionó anteriormente está definida con material incompleto, mal preservado y no diagnóstico. El planteamiento de Alberdi y Prado de agrupar en el género Stegomastodon los mastodontes suramericanos diferentes del género Cuvieronius, tiene fuertes cimientos en cuanto a la gran variabilidad de caracteres intraespecíficos observados en los restos fósiles de mastodontes, en dónde por ejemplo se observan aspectos morfológicos diferentes incluso en molares derechos e izquierdos del mismo individuo; ya Cabrera (1929), menciona la gran variabilidad intraespecífica presente en Elefantes Africanos de una misma población en Kenia, en donde tan solo por la forma de las defensas él habría podido distinguir cuando menos seis o siete tipos o formas muy diferentes entre sí. Con respecto a la propuesta de Mothé et al (2011) hay que considerar que la diagnosis del género Notiomastodon definida en su momento por Cabrera, establece "â¦incisivos superiores sencillamente encorvados en todas las edades, y con una banda lateral de esmalte,", característica que puede estar presente en individuos juveniles o adultos jóvenes.
Las razones expuestas anteriormente, así como las características de los molares del ejemplar en estudio conducen a optar por la propuesta de Alberdi y Prado en cuanto a la clasificación de la pieza. El principal carácter diagnóstico que nos permite ubicar al ejemplar dentro de la especie Stegomastodon waringi (HOLLAND, 1920) de acuerdo con lo expuesto por Alberdi y Prado, entre otros autores, es que los dos lados del premaxilar (y la parte más anterior del maxilar) junto con los alveolos de las defensas son casi paralelos, además que la incisura dental es estrecha.
El ejemplar presenta los segundos y terceros molares deciduos, los cuales se identifican como tal por varias razones, la primera de ellas es ser trilofodontos, la segunda es que los molares incluidos en la zona maxilar poseen más de 3 lofos y la tercera es que un primer diente deciduo dM1 debería haber sido bilofodonto (de acuerdo con Mothé et al, 2010). Se decidió llamar deciduos a los molares expuestos del mastodonte debido a que al ser trilofodontos serán reemplazados tarde o temprano por molares definitivos con más de tres lofos. (Aunque también sería válido llamarlos M1 y M2 si se considera que el Molar totalmente incluido en el maxilar corresponde con el M3).
Sin embargo, tal y como se describió en la introducción, la taxonomía de los mastodontes suramericanos es un tema aún por resolver y a la hora de identificar y ubicar taxonómicamente restos fósiles de proboscídeos, se debe tener en cuenta la intrincada historia taxonómica del grupo, y por supuesto, esperar a que la Comisión Internacional de Nomenclatura Zoológica se pronuncie en este caso y dictamine las directrices a seguir en la materia.
Agradecimientos
A la Dra. María Páramo por sus sugerencias y comentarios. A la Dra. María Teresa Alberdi, por el envío de bibliografía y sus valiosas observaciones. Al Dr. Marco Ferretti por resolver amablemente algunas preguntas hechas vía e-mail. Al señor Gerardo Vargas por la dedicación y ayuda en la curatoría y preparación de la pieza. A Gilberto Mendoza por su interés. Gracias también al Profesor Wilson Blanco de la Universidad de Cartagena y a José Enrique Arenas, director del Museo Geológico Nacional por su interés y las gestiones administrativas requeridas para el traslado del material a la ciudad de Bogotá.
La investigación se encontró amparada por los proyectos de inversión nacional 'BPIN Ampliación del Conocimiento Geológico y Recursos del Territorio Colombiano y 'BPIN Ampliación del Conocimiento Geológico y del Potencial de Recursos del Subsuelo de la Nación' Código 0043-00015- 0000.
Bibliografía
Alberdi, M. T., Prado J.L. 1995. Los Mastodontes de América del Sur. En: Alberdi, M.T., Leone, G., y Tonny, E.P. (eds.). Evolución Biológica y climática de la región pampeana durante los últimos 5 millones de años. Un ensayo de correlación con el mediterráneo Occidental. Monografías, Museo de Ciencias Naturales, CSIC, Madrid, 12: 277-292. [ Links ]
Alberdi M.T., Prado J.Ll, Cartelle C. 2002. El registro de Stegomastodon (Mammalia, Gomphotheriidae) en el Pleistoceno Superior de Brasil. Revista Española de Paleontología, 17(2): 217-235. [ Links ]
Alberdi, M. T., Corona-M, E. 2005. Revisión de los Gonfoterios en el Cenozoico tardío de México. Revista Mexicana de Ciencias Geológicas, v. 22. P. 246-260. [ Links ]
Asevedo, L., Winck, G. R., Mothé, D., Avilla, L. S. 2011. Ancient diet of the Pleistocene gomphothere Notiomastodon platensis (Mammalia, Proboscidea, Gomphotheriidae) from lowland mid-latitudes of South America: Stereomicrowear and tooth calculus analyses combined. EN PRENSA. Quaternary International (2011), doi:10.1016/j. quaint.2011.08.037. [ Links ]
Cabrera, A., 1929. Una revisión de los Mastodontes Argentinos. Revista del Museo de La Plata 32: 61-144. [ Links ]
Demirjian, V. 2010. Comment on the proposed conservation of usage of Mastodon waringi Holland, 1920 (currently Haplomastodon waringi; Mammalia, Proboscidea) by designation of a neotype. Bulletin of Zoological Nomenclature. V. 67. Issue 4, pp. 333. [ Links ]
Dominato, V. H., Mothé, D., Costa Da Silva, R., Avilla, L. 2011. Evidence of scavenging on remains of the gomphothere Haplomastodon waringi (Proboscidea: Mammalia) from the Pleistocene of Brazil: Taphonomic and paleoecological remarks. Journal of South American Earth Sciences 31 (2011) 171-177 [ Links ]
Ferretti, M. P., 2008. A Review of South American Proboscideans. In: Lucas, S. G., Morgan, G. S., Spielmann, J. A., Prothero, D. R. (Eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin, 44, pp. 381-392. [ Links ]
Ferretti, M. P., 2009. Comment on the proposed conservation of usage of Mastodon waringi Holland, 1920 (currently Haplomastodon waringi; Mammalia, Proboscidea) by assignation of a neotype. Bulletin of Zoological Nomenclature. V. 66, Issue 4, pp. 358-359. [ Links ]
Ferretti, M. P., 2010. Anatomy of Haplomastodon chimborazi (Mammalia, Proboscidea) from the late Pleistocene of Ecuador and its bearing on the phylogeny and systematics of South American gomphotheres. Geodiversitas:32 (4). Publications Scientifiques du Muséum national d’Histoire naturelle, Paris.663-721. [ Links ]
Ficcarelli, G., Azzaroli, A., Borselli, V., Coltorti, M., Dramis, F., Fejfar, O., Hirtz, A., Torre, D. 1993. Stratigraphical and Paleontological aspects of Late Quaternary deposits in the Interandean Depression of Northern Ecuador. Journal of South American Earth Sciences, 6,3: 145-150. [ Links ]
Ficcarelli, G., Borselli, V., Herrera, G., Moreno-Espinosa, M., Torre, D. 1995. Taxonomic remarks on the South American Mastodons referred to Haplomastodon and Cuvieronius. Geobios, 28(6): 745-756. [ Links ]
Hoffstetter, R., 1950. Observaciones sobre los mastodontes de Sud América y especialmente del Ecuador. Haplomastodon, subgen. Nov. de Stegomastodon; Publicaciones Escuela Politécnica Nacional, 1950, v. I, p.1-49. [ Links ]
Hoffstetter, R., 1952. Les Mammifères Pléistocènes de la République de l'Equateur: Mémoires des Société Géologique de France, v. 66, p. 1-391. [ Links ]
Holland, W. J., 1920. Fossil mammals collected at Pedra Vermelha, Bahia, Brazil, by Gerald A. Waringi. Annals of Carnegie Museum, 13, 224- 232. [ Links ]
Lucas, S.G. 2010. Comment on the proposed conservation of usage of Mastodon waringi Holland, 1920 (currently Haplomastodon waringi; Mammalia, Proboscidea) by designation of a neotype. Bulletin of Zoological Nomenclature. V. 67. Issue 2. Pp. 181-182. [ Links ]
Madden, C. T., 1984. The Proboscidea of South America. Geological Society of America, abstracts with programs 12: 474. [ Links ]
Mothé, D., Avilla, L. S., Winck, G. R., 2010. Population structure of the gomphothere Stegomastodon waringi (Mammalia: Proboscidea: Gomphotheriidae) from the Pleistocene of Brazil. Anais da Academia Brasileira de Ciéncias 82 (4): 983-996. [ Links ]
Mothé, D., Avilla, L. S., Cozzuol, M., Winck, G. R. 2011. Taxonomic revision of the Quaternary gomphotheres (Mammalia: Proboscidea: Gomphotheriidae) from the South America lowlands. IN PRESS. Quaternary International (2011), doi:10.1016/j.quaint.2011.15.018. [ Links ]
Osborn, E. C., 1923. New subfamily, generic, and specific stages in the evolution of the Proboscidea. Amer. Mus. Novitates, No. 99, pp. 1-4. [ Links ]
Pasenko, M. R. 2010. Comment on the proposed conservation of usage of Mastodon waringi Holland, 1920 (currently Haplomastodon waringi; Mammalia, Proboscidea) by designation of a neotype (Case 3480). Bulletin of Zoological Nomenclature. V. 67. Issue 1. Pp. 96. [ Links ]
Pohlig, H., 1912. Sur une vieille mandibule de « Tetracaulodon ohioticum ». Blum., avec défense in situ. Bulletin de la Société Belge de Géologie, de Paléontologie et D'Hydrologie. Bruselles. Tome XXVI, pp. 187-193. [ Links ]
Prado, J. L., Alberdi, M. T., Azanza, B., Sánchez, B., Frassinetti, D., 2005. The Pleistocene Gomphotheriidae (Proboscidea) from South America. Quaternary International 126-128, 21-30. [ Links ]
Simpson, G. G., De Paula Couto, C., 1957. The Mastodons of Brazil. American Museum of Natural History 112 (2), 131-185. [ Links ]
Recibido: 26 de septiembre de 2011
Aceptado para publicación: 2 de junio de 2012
