










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
versión impresa ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.36 no.141 Bogotá oct./dic. 2012
BIOQUÍMICA
*Â Grupo Insight. Universidad Militar Nueva Granada. Centro de Investigaciones de la Clínica del Country
RESUMEN
El antígeno EBA-181 del Plasmodium falciparum se une al glóbulo rojo sugiriendo un papel en la invasión por parte del merozoito. Se aplicó una metodología previamente desarrollada con base en la probabilidad y la entropía, a la caracterización de péptidos de alta unión de EBA-181 al glóbulo rojo. Se calcularon los valores de probabilidad, sumatoria de probabilidad y entropía para las 79 secuencias no sobrelapadas de la proteína; también se efectuaron los cálculos para péptidos teóricos análogos construidos a partir de los péptidos de unión. Las secuencias específicas comprobadas experimentalmente como de alta unión presentaron valores dentro de los rangos asociados al macroestado unión, diferenciando las secuencias que se unen de las que no con un acierto en el 100% de los casos estudiados.
Palabras Clave: Probabilidad, entropía, alta unión, EBA-181, eritrocito, merozoito.
ABSTRACT
EBA-181, antigen of Plasmodium falciparum, binds to red blood cells, suggesting a role in the merozoite invasion. Â It was applied a previously developed methodology based on probability and entropy, to the characterization of high-binding EBA-181 peptides to red blood cells. Values of probability, probability summation and entropy were calculated for the 79 not overlapped sequences of the protein; the calculations were also made for analogous theoretical peptides, built from the binding peptides. The specific sequences experimentally proved as high binding ones had values into the ranges associated to the binding macrostate, differentiating the sequences that bind and not bind with a right guess of 100% in the studied cases.
Key Words: Probability, Entropy, High binding, EBA-181, erythrocyte, merozoite.
Introducción
La malaria es una enfermedad parasitaria que se presenta especialmente en áreas tropicales constituyendo en el mundo una de las principales causas de morbimortalidad (World Health Organization, 2009; Shuman, 2010; Ministerio de Protección Social, 2006). Un significativo número de los individuos que padecen de malaria son contagiados por el Plasmodium falciparum. La invasión periódica del Plasmodium a los eritrocitos es un proceso que requiere el reconocimiento, unión, orientación e internalización del merozoito dentro del eritrocito; cada una de estas acciones es mediada por interacciones receptor ligando, que caracterizan el ciclo eritrocítico de invasión de la malaria (Aikawa et al., 1978; Chitnis et al., 2000). La proteína del Plasmodium falciparum EBA-181 (o Erytrhocyte Binding Antigen-181), es una proteína de 1567 aminoácidos, que tiene una estructura primaria característica de los miembros de la familia de proteínas ebl. Â La proteína EBA-181 se encuentra en micronemas y se expresa simultáneamente a la EBA-175, a la que además parece manifestar una función análoga, así como también a EBA-140 (Adams et al., 2001; Gilberger et al., 2003). Â La expresión de EBA -181 difiere a lo largo de las cepas de P. falciparum. Su estructura primaria y el hecho de que se une a los glóbulos rojos, sugiere que desempeña un papel en la invasión del merozoito. Â La interrupción selectiva del gen EBA-181 no ha tenido ningún efecto sobre el fenotipo de invasión del parásito, lo que sugiere que podría actuar como un ligando alternativo (Mayer et al., 2004).
Para cuantificar la posibilidad de ocurrencia de un evento es empleada una medida matemática denominada probabilidad (Feynman et al., 1998a). En algunos sistemas complejos, como el gaseoso, el análisis individual de cada partícula no permite establecer su estado dinámico, pues tienen un número de moléculas del orden del número de Avogadro (1023), por lo tanto se debería emplear 1023 ecuaciones diferenciales. Surgieron  teorías nuevas para solucionar este problema: la termodinámica y la mecánica estadística. La medida proporcional al logaritmo del número establecido de microestados propios de un sistema en estado de equilibrio para que desde el punto de vista macroscópico, éste permanezca, está dada por la definición de la Entropía dada por Boltzmann en el contexto de la teoría cinética de los gases (Feynman et al., 1998b). Después de la termodinámica se fundamentó la mecánica estadística, con las probabilidades de las distribuciones microscópicas (Feynman et al., 1998b).
Las definiciones de macroestado y microestado han sido utilizadas con el objetivo de caracterizar el estado de sistemas como el gaseoso, y se ha dicho que la primera indica una distribución general del sistema, mientras que la definición de microestado señala las distintas configuraciones posibles para estas distribuciones. Shannon expresó que el concepto de la entropía de la mecánica estadística corresponde al encontrado en la teoría de la información, al buscar la forma de cuantificar la cantidad de información que contiene cualquier mensaje, con su correspondiente análisis como medida objetiva de la cantidad de información que posee un sistema (Frodden et al., 2004).
La probabilidad y la entropía fueron aplicadas para el desarrollo de una nueva metodología de caracterización de péptidos de alta unión al eritrocito (Rodríguez, 2008a); se analizaron la totalidad de las secuencias de 20 aminoácidos componentes de la proteína de superficie del merozoíto MSP-1, encontrando para las secuencias comprobadas experimentalmente de unión, valores de probabilidad, sumatoria de probabilidad y entropía dentro de los rangos asociados al macroestado de unión, y valores fuera de estos rangos para las secuencias de no unión, diferenciando acertadamente la pertenencia de las secuencias a cada uno de los macroestados en el 100% de los casos (Rodríguez, 2008a). Â Estos resultados son importantes pues evidenciaron un orden físico y matemático del fenómeno de unión, lo cual es importante para posteriormente realizar predicciones de unión ligando receptor, cruciales para la biología molecular. Desde esta perspectiva es necesario aplicar esta metodología a otras proteínas para comprobar su alcance. En este contexto, el propósito de este trabajo es la caracterización matemática de la unión de péptidos de la proteína EBA-181 al receptor del glóbulo rojo, mediante la aplicación de la metodología mencionada con base en la teoría de la probabilidad y la ley de la entropía.
Definiciones
Macroestado de unión: Representa a las secuencias que muestran alta unión al receptor del glóbulo rojo (Rodríguez, 2008a).
Macroestado de no unión: Representa a las secuencias que no muestran alta unión al receptor del glóbulo rojo (Rodríguez, 2008a).
Microestado: Cada una de las secuencias compuesta por un total de veinte aminoácidos (Rodríguez, 2008a).
Tipo de secuencia: Grupo particular de microestados que presentan valores similares de probabilidad, sumatoria de probabilidad y entropía (Rodríguez, 2008a).
Probabilidad Laplaciana [P(Ai)]: Probabilidad de un aminoácido A en una posición específica i, en la que se establece la relación entre la frecuencia de aparición de dicho aminoácido en una posición específica y el total de aminoácidos (Feynman et al., 1998a).

Probabilidad de una secuencia [P(S)]: Resulta de la multiplicación de las probabilidades de los aminoácidos constituyentes por posición específica i (Feynman et al., 1998a).
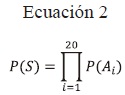
Sumatoria de probabilidad (SumP): Resulta de la suma de las probabilidades aisladas de cada aminoácido que conforma cada secuencia, en donde se cuantifican las probabilidades de aparición de un aminoácido particular en una posición específica.
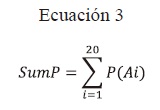
Entropía (S): Cuando se trata de sistemas que se componen de múltiples microestados con probabilidades divergentes, la entropía se define como (Frodden et al., 2004):
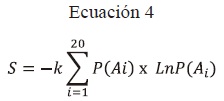
k = Constante de Boltzmann [1,38 × 10-23(J/K)].
S= Valor de la entropía.
P(Ai)= Valor de la probabilidad del aminoácido A en la posición i.
Materiales y métodos
Se estudiaron las 79 secuencias de 20 aminoácidos de longitud de la proteína EBA-181 reportadas por Vera-Bravo et al. (2005), sin ningún tipo de sobrelapamiento. Sólo de ellas 5 han sido comprobadas experimentalmente de alta unión al receptor del glóbulo rojo (Vera-Bravo et al, 2005), mientras que las 74 restantes no son de alta unión. A partir de estas secuencias conocidas como de alta unión se estableció la frecuencia de aparición de cada uno de los 20 aminoácidos existentes en las 20 posibles posiciones para las secuencias peptídicas, con la cual se construyó el espacio total de probabilidad, en el cual se determina la probabilidad laplaciana de las 5 secuencias comprobadas experimentalmente como de alta unión, mediante la ecuación 1. Un proceso idéntico se realizó con los 74 péptidos de 20 aminoácidos de no unión al receptor del glóbulo rojo, determinado de esta forma un espacio de probabilidad para el macroestado de no unión.
Con el fin de realizar la caracterización matemática de cada uno de los macroestados, tanto de unión como de no unión, se determinan los valores de probabilidad a partir de la Ecuación 2, sumatoria de probabilidad de acuerdo con la Ecuación 3 y de entropía con  la ecuación 4, para la totalidad de los péptidos de EBA-181, tal como se efectuó en el trabajo previo (Rodriguez 2008a).
Tras determinar los valores característicos de cada macroestado, se crearon análogos teóricos de los péptidos de unión, y se realizaron los cálculos correspondientes de probabilidad, sumatoria de probabilidad y entropía, para así comparar el péptido de unión con cada uno de sus posibles péptidos análogos, al reemplazar cada uno de sus residuos constitutivos por Glicina, y con estas mutaciones teóricas ver la diferencia de los valores encontrados para cada una de ellas con respecto al péptido inicial.
Resultados
Para las 5 secuencias asociadas al macroestado de unión, las frecuencias de aparición de los aminoácidos se establecieron entre 0 y 3, correspondiendo este número mayor al aminoácido I en la posición 5 (Tabla 1). En el caso de las secuencias peptídicas asociadas al macroestado de unión la probabilidad laplaciana por posición específica se mantuvo entre 0 y 0,030, siendo el valor máximo asociado al aminoácido I, que tuvo mayor número de repeticiones en la posición 5 (Tabla 2).

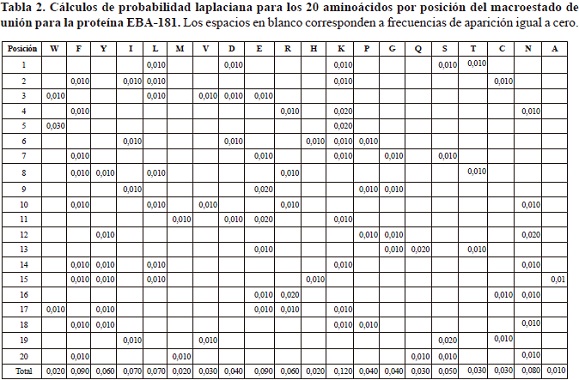
Paralelamente, las secuencias asociadas al macroestado de no unión presentaron frecuencias de aparición de los aminoácidos por posición específica que variaron entre 0 y 14, asociándose este valor mayor al aminoácido E en la posición 9 (tabla 3), el cual además corresponde a la mayor probabilidad laplaciana presentada para el macroestado de no-unión, cuyo valor fue 0,009 (tabla 4).
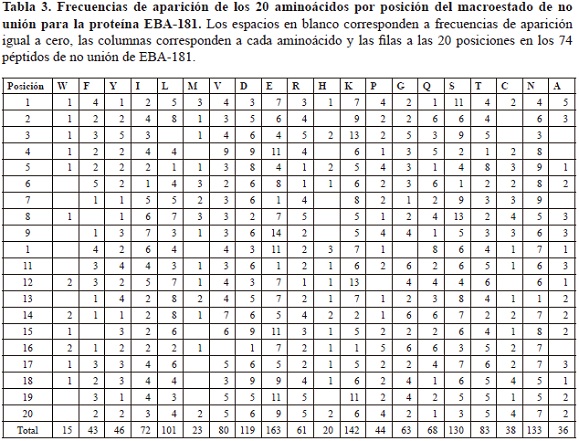
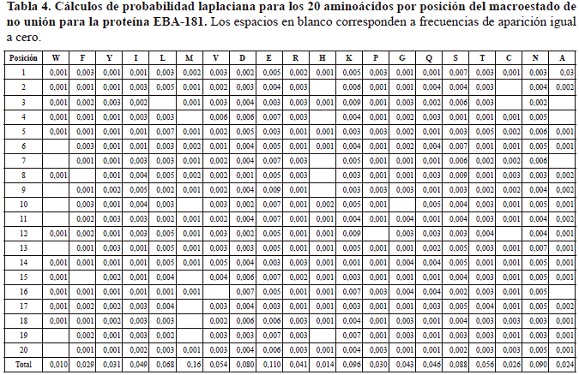
Los valores de probabilidad, sumatoria de probabilidad y entropía para el macroestado de unión variaron entre 8,00E-40 y 1,60E-39, 0,23 y 0, 026, y 1,53E-23 y 1,40E-23 respectivamente.
Para el macroestado de no unión los valores de probabilidad se encontraron entre 2,00E-22 y 1, los de sumatoria de probabilidad oscilaron entre 0 y 0,12, y los valores de entropía se encontraron en el rango comprendido entre 6,36E-25 y 1,27E-24 (tabla 5), encontrándose además un valor de 0, correspondiente a una de las secuencias peptídicas de no unión, que además de este valor específico de entropía, presentó valores bien diferenciados dentro del rango de valores de no unión, siendo la secuencia FRKQRFTSMHYIAGGGIIAL, cuyos valores respectivos de probabilidad y sumatoria de probabilidad fueron 1 y 0.
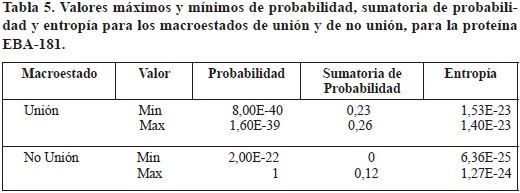
Los valores de probabilidad, sumatoria de probabilidad y entropía de las secuencias peptídicas comprobadas experimentalmente como de alta unión se mantuvieron dentro del rango asociado al macroestado de unión, que se mantiene en un rango diferenciado del hallado para el macroestado de no unión.
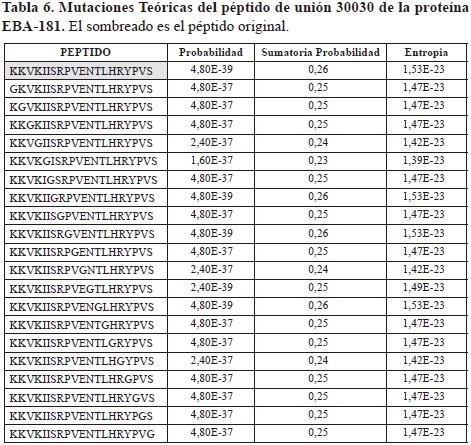
Para las 20 secuencias teóricas del péptido de unión 30030, los valores de probabilidad oscilaron entre 4,80E-39 y 1,60E-37, siendo el primero de ellos equivalente al hallado en el péptido original; los valores de sumatoria de probabilidad estuvieron entre 0,24 y 0,26, siendo este ultimo el obtenido en el péptido original, y los valores de entropía se encontraron entre 1,53E-23 y 1,39E-23, correspondiendo el primero de ellos al hallado para el original. Los valores calculados para las mutaciones teóricas de este péptido, se muestran como ejemplo de estos resultados teóricos en la Tabla 6. Â Para las 18 secuencias teóricas generadas como análogas al péptido 30031 los valores de probabilidad, sumatoria de probabilidad y entropía oscilaron entre 6,00E-40 y 2,00E-38, entre 0,2 y 0,23, y entre 1,40E-23 y 1,25E-23 respectivamente. Para las 19 secuencias teóricas análogas del péptido 30045, los valores de probabilidad se encontraron entre 8,00E-40 y 4,00E-38, los valores de sumatoria de probabilidad estuvieron entre 0,21 y 0,23, y los valores de entropía entre 1,40E-23 y 1,30E-23. Para las 19 secuencias teóricas del péptido 30051, los valores se encontraron respectivamente entre 8,00E-40 y 1,60E-37, entre 0,22 y 0,24, y entre 1,45E-23 y 1,34E-23. Finalmente, para las 20 secuencias teóricas generadas para el péptido 30056, los valores de probabilidad oscilaron entre 4,80E-39 y 1,60E-37, los valores de sumatoria de probabilidad estuvieron entre 0,23 y 0,26, y los valores de entropía se hallaron entre 1,53E-23 y 1,39E-23. Â La variación en la cantidad de secuencias teóricas generadas para cada péptido de unión se debe a la cantidad de glicinas (G) que presenta cada uno en su secuencia; por ejemplo, para el péptido 30031 (SFLNIKKYGRKGEYLNRNSF) se generaron 18 secuencias, ya que este presenta dos glicinas en su secuencia, en las posiciones 9 y 12, de manera que estas posiciones no son reemplazadas. Los valores encontrados en todos los cálculos para los péptidos teóricos análogos generados se encontraron dentro de los rangos hallados de unión y en ocasiones tomaron valores un poco menores a los hallados para dicho rango, pero en ningún caso se encontraron dentro de los rangos hallados para el macroestado de no-unión.
Discusión
Este es el primer trabajo en el que a partir de una caracterización matemática del fenómeno de unión para la proteína EBA-181 mediante la probabilidad y la entropía, partiendo del análisis de la totalidad de secuencias peptídicas no sobrelapadas de 20 residuos de la proteína, se diferenciaron rangos de valores de probabilidad, sumatoria de probabilidad y entropía asociados a los macroestados de unión y de no unión, diferenciando las secuencias que se unen de las que no se unen con un acierto del 100% en los casos estudiados. Se crearon análogos teóricos de los péptidos de unión, reemplazando cada uno de sus aminoácidos constitutivos por glicina, a fin de comparar sus valores encontrados de probabilidad, sumatoria de probabilidad y entropía, con respecto a los hallados en el péptido original de unión desarrollando una primera metodología predictiva de este método experimental de ensayo y error. Este método de péptidos teóricos análogos es una metodología nueva en esta disciplina, que está siendo refinada para establecer predicciones útiles.
Este trabajo se basa en un estudio previo realizado por Rodríguez (2008a), en el que se desarrolló la metodología para la caracterización de péptidos de alta unión de la proteína MSP-1; esta metodología fue aplicada posteriormente a la caracterización de péptidos de las proteínas EBA-140 (Rodriguez et al., 2009a), y MSA-2 (Rodriguez et al., 2009b), obteniendo el mismo nivel de acierto en todos los casos, equivalente al 100%.  En estos trabajos se encontraron para todas las proteínas estudiadas rangos de valores diferenciados respectivos a los cálculos de probabilidad, sumatoria de probabilidad y entropía  asociados a los macroestados de unión y de no unión; en el presente trabajo se hallaron valores dentro de rangos claramente diferenciados para todos los cálculos realizados, aclarando que para el caso de la entropía se encontró además un valor de 0 para una de las secuencias de no unión estudiadas, la secuencia FRKQRFTSMHYIAGGGIIAL, cuyo valor de sumatoria de probabilidad también fue de 0, y cuyo valor de probabilidad correspondió al más alto valor encontrado, siendo equivalente a 1.
Esta metodología evidencia la existencia de un orden físico y matemático subyacente al fenómeno de unión implicado en la invasión al glóbulo rojo por el merozoito. Estos trabajos, realizados desde una perspectiva acausal, permiten una comprensión físico-matemática a partir de la cual sería posible generar predicciones que superen la efectividad inherente a la metodología de ensayo y error actualmente empleada, simplificando el proceso y economizando recursos. Â Desde esta perspectiva se han desarrollado teorías de predicción para el fenómeno de unión de péptidos, que pueden ser útiles para facilitar la escogencia de péptidos a usar en el desarrollo de vacunas. La primera se basa en las leyes de probabilidad, combinatoria y entropía (Rodríguez 2008b), y fue aplicada a 100 secuencias específicas de péptidos teóricos, promiscuos, sintéticos y naturales que se unen a moléculas de HLA clase II y 61 péptidos que no se unen; se desarrolló una predicción de tipo teórico para el conjunto apropiado de secuencias que debe tener un péptido para ser presentado por el HLA clase II, con la que se acertó en el 100% de los casos comprobados experimentalmente. La segunda fue desarrollada con la teoría de la probabilidad (Rodriguez et al., 2010), a partir de la cual se realizó una inducción con 45 péptidos específicos, calculando valores predictivos para los péptidos de unión y de no unión al glóbulo rojo; se realizaron medidas de desempeño de las predicciones respecto a los hallazgos experimentales, obteniendo como resultado una sensibilidad del 95% y una especificidad del 90%.
La perspectiva física acausal también ha sido base para el desarrollo de metodologías predictivas en otras áreas de la medicina. En el campo de la epidemiología, se desarrolló una predicción de la dinámica temporal de la malaria en 820 municipios de Colombia para los años 2003 a 2007, a partir de la probabilidad y la entropía, estableciendo relaciones específicas que permiten determinar la correspondencia o no de la dinámica a un brote (Rodríguez, 2010a); este método puede ser útil como soporte para la planificación de intervenciones y la toma de decisiones en salud pública. También se han hallado resultados predictivos en el área de cardiología, donde a partir de las concepciones de los sistemas dinámicos y la aplicación de leyes de la probabilidad y la entropía, se desarrolló una metodología que permite predecir las diferencias de las dinámicas cardiacas agudas, crónicas, normales y estados de evolución entre normalidad y enfermedad, la cual es útil como herramienta de ayuda diagnóstica en la clínica (Rodríguez 2010b). Con base en la geometría fractal se desarrolló una generalización teórica con la que se determinó un total de 69.049 prototipos arteriales posibles normales y enfermos en el proceso de restenosis, a partir de una  simulación computacional basada en el concepto de Armonía Matemática Intrínseca aplicada a dimensiones fractales de los objetos componentes de las arterias (Rodríguez et al, 2010c). En el área de morfofisiología celular, la geometría fractal y la aplicación de los conceptos de Armonía Matemática Intrínseca celular y de variabilidad, sustentan el desarrollo de un nuevo diagnóstico de las alteraciones celulares preneoplásicas y neoplásicas en citologías cervicales (Rodríguez, 2011). La ocupación de atractores caóticos en el espacio fractal de Box-counting, ha permitido la diferenciación entre dinámicas cardiacas agudas de aquellas dentro de límites de normalidad y con enfermedades crónicas, metodología diagnóstica aplicada obteniendo un valor de 100% de sensibilidad y especificidad, así como un coeficiente Kappa de 1, en el estudio comparativo de pacientes diagnosticados con enfermedad aguda y normales (Rodríguez et al, 2011a).
El trabajo realizado evidencia órdenes físico-matemáticos subyacentes a los diversos fenómenos propios de la medicina, favoreciendo el desarrollo de métodos predictivos que no requieren de análisis estadísticos ni de grandes pruebas experimentales, como sucede específicamente con las pruebas de ensayo y error empleadas para el estudio del fenómeno de unión.
Agradecimientos
A la Universidad Militar Nueva Granada, especialmente a la Dra. Esperanza Fajardo, Directora de Investigaciones de la Facultad de Medicina, al Dr. Juan Miguel Estrada, Decano de la Facultad de Medicina y a la Vicerrectoría de investigaciones, en especial al Dr. Ricardo Cure Hakim, Vicerrector y a Henry Acuña, Jefe División de Investigación Científica, por su apoyo a nuestras investigaciones.
Este trabajo es resultado del proyecto de Investigación PIC MED-785, financiado por el Fondo de Investigaciones de la Universidad Militar Nueva Granada, por lo cual agradecemos su apoyo a nuestras investigaciones.
Al Centro de Investigaciones de la Clínica del Country, en especial a todas las personas que trabajan en el mismo, por su apoyo efectivo.
Dedicatoria
A nuestros hijos y a Joshua, ahijado del primer autor.
Referencias bibliográficas
Adams JH, Blair PL, Kaneko O, Peterson DS. 2001. An expanding ebl family of Plasmodium falciparum. Trend Parasitol. 17(6):297-9. [ Links ]
Aikawa M, Miller LH, Johnson J, Rabbege. 1978. Erythrocyte entry by malarial parasites. A Moving junction between erythrocyte and parasite. J Cell Biol. 77:72-82. [ Links ]
Chitnis CE, Blackman MJ. 2000. Host cell invasion by malaria parasites. Parasitol. 16:411-415. [ Links ]
Gilberger TW, Thompson JK, Reed MB, Good RT, Cowman AF. 2003. The cytoplasmic domain of the Plasmodium falciparum ligand EBA-175 is essential for invasion but not protein trafficking. J Cell Biol. 162(2):317-27. [ Links ]
Feynman RP, Leighton RB, Sands M. 1998a. Física, Probabilidad, Vol. 1. Addison-Wesley Iberoamericana, Wilmington, S. A. México, p. 6-1, 6-16. [ Links ]
Feynman RP, Leighton RB, Sands M. 1998b. Física. Leyes de la Termodinámica, Vol. 1. Addison-Wesley Iberoamericana, Wilmington, S. A. México p. 44-1, 44-19. [ Links ] [ Links ]
Mayer DC, Mu JB, Kaneko O, Duan J, Su XZ, Miller LH. 2004. Polymorphism in the Plasmodium falciparum erythrocyte-binding ligand JESEBL/EBA-181 alters its receptor specificity. Proc Natl Acad Sci U S A. 11(8):2518-23. [ Links ]
Ministerio de Protección Social. 2006. Guía de atención de la malaria. Bogotá [ Links ].
Rodríguez J. 2008a. Caracterización física y matemática de péptidos de alta unión de MSP-1 mediante la aplicación de la teoría de la probabilidad y la entropía. Arch alerg inmunol clin. 39(2):74-82. [ Links ]
Rodríguez J. 2008b. Teoría de unión al HLA clase II: Teoría de probabilidad, combinatoria y Entropía aplicadas a secuencias peptídicas. Inmunología; 27:151-166. [ Links ]
Rodríguez J, Correa C, Prieto S, Puerta G, Vitery S, Bernal P, Soracipa Y, Botero D. 2009a. Aplicación de la probabilidad y la entropía a la proteína EBA-140. Caracterización matemática de péptidos de alta unión. Inmunología. 28(2):65-73. [ Links ]
Rodríguez J, Correa C, Prieto S, Cardona D, Vitery S, Puerta G, Soracipa Y, Bernal P. 2009b. Caracterización física y matemática de péptidos de alta unión de MSA-2 aplicación de la teoría de la probabilidad y la entropía. Rev Acad Colomb Cienc. 33(129):549-557. [ Links ]
Rodríguez J, Bernal P, Prieto S, Correa C. 2010. Teoría de péptidos de alta unión de malaria al glóbulo rojo. Predicciones teóricas de nuevos péptidos de unión y mutaciones teóricas predictivas de aminoácidos críticos. Inmunología. 29(1):7-19. [ Links ]
Rodríguez J. 2010a. Método para la predicción de la dinámica temporal de la malaria en los municipios de Colombia. Rev Panam Salud Pública. 27(3):211-8. [ Links ]
Rodríguez J. 2010b. Entropía Proporcional De Los Sistemas Dinámicos Cardiacos: Predicciones físicas y matemáticas de la dinámica cardiaca de aplicación clínica. Rev Colomb Cardiol. 17:115-129. [ Links ]
Rodríguez J, Prieto S, Correa C, Bernal P, Puerta G, Vitery S, Soracipa Y, Muñoz D. 2010c. Theoretical generalization of normal and sick coronary arteries with fractal dimensions and the arterial intrinsic mathematical harmony. BMC Medical Physics. 10:1 http://www.bio-medcentral.com/1756-6649/10/1 [ Links ]
Rodríguez J. 2011. Generalización fractal de células preneoplásicas y Cancerígenas del epitelio escamoso cervical. Una nueva Metodología de aplicación clínica. Rev. Fac Med. 18(2): 173-181. [ Links ]
Rodríguez J, Prieto S, Bernal P, Soracipa Y, Salazar G, Izasa D, Cruz LE, Correa C. 2011a. Nueva metodología diagnóstica de la dinámica geométrica cardiaca. Dinámica cardiaca caótica del Holter. Rev Acad Colomb Cienc 35(134):5-12. [ Links ]
Shuman E. 2010. Global Climate Change and Infectious Diseases. N Engl J Med. 362:12. [ Links ]
Vera-Bravo R, Valbuena J, Ocampo M, Garcia J, Rodríguez L, Puentes A, López R, et al. 2005. Amino terminal peptides from the Plasmodium  falciparum  EBA-181/JESEBL  protein  bind  specifically to erythrocytes and inhibit in vitro merozoite invasion. Biochimie 87:425-436. [ Links ]
World Health Organization, United Nations. World malaria report 2009. Geneva, Switzerland. WHO Press 2009. [ Links ]
Recibido: 27 de julio de 2012
Aceptado para publicación: 16 de octubre de 2012
