










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales
Print version ISSN 0370-3908
Rev. acad. colomb. cienc. exact. fis. nat. vol.40 no.157 Bogotá Oct./Dec. 2016
https://doi.org/10.18257/raccefyn.403
doi: http://dx.doi.org/10.18257/raccefyn.403
1 Compensation International Progress S. A. -Ciprogress Greenlife-, Bogotá, Colombia
2 UNELLEZ-Guanare, Programa de Ciencias del Agro y el Mar, Herbario Universitario (PORT), Mesa de Cavacas, Estado Portuguesa, Venezuela
3 Grupo de Investigación en Biodiversidad y Conservación, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia
Abstract
Rapatea isanae, from the upper Isana [Içana] river, Guianía Department, Colombia, is described and illustrated, and its morphological relationships with allied species are discussed. This taxon is remarkable for Rapatea in its small stature (15-30 cm tall) and leaves, and it is only the second species with white petals in a genus that otherwise has only yellow petals. It is most closely R. spruceana, with the base of the leaf blade gradually tapering from the intergrading into the petiole, and the bractlet of the spikelet and sepal shape. However, this new species differs from R. spruceana in its shorter size, sparse verrucose on the lower surface, the length of the attenuate portion of the involucral bracts, and the shape and color of the petals. It also has similarities to R. longipes and R. modesta in its ventricose sheath-leaf, inflorescence shape, and bractlets equal in length. A previously described species from Colombia is restablished (i.e., R. modesta), and one variety is elevated to the rank of species (i.e., R. paludosa var. sessiliflora to R. sessiliflora). An updated key to the all twenty five species of Rapatea, and an abstract in Kuripaco are also provided.
Key words: Rapatea; sect. Longipes; Colombia; Isana [Içana] river; upper Rio Negro; Rapateaceae.
Resumen
Rapatea isanae, una nueva especie del alto río Isana, Departamento del Guianía, Colombia, es descrita, ilustrada, y sus relaciones morfológicas con las especies afines son discutidas. Esta especie es notable en el género Rapatea por el tamaño de las plantas (entre 15 y 30 cm de altura), las hojas muy pequeñas, y los pétalos blancos, siendo la segunda especie con este carácter en un género en el cual predominan los pétalos amarillos. Por la base de las hojas transformándose gradualmente en un pecíolo, la morfología de las bractéolas de las espiguillas y los sépalos, R. isanae está relacionada con R. spruceana. Por las vainas basales de las hojas ventricosas, la forma convexa de las inflorescencias y las bractéolas del mismo tamaño con R. longipes y R. modesta, respectivamente. Sin embargo, esta nueva especie difiere de R. spruceana por ser plantas muy pequeñas, el envés esparcidamente verrugoso, el largo de la porción atenuada de las brácteas involúcrales y la forma y color de los pétalos. Se restablece una especie previamente descrita para Colombia (i.e., R. modesta), y una variedad es elevada al rango de especie (i.e., R. paludosa var. sessiliflora a R. sessiliflora). Adicionalmente se presenta una clave actualizada de las 25 especies aceptadas de Rapatea y un resumen en Kuripaco.
Palabras clave: Rapatea; sect. Longipes; Río Isana [Içana]; Alto Río Negro; Colombia; Rapateaceae.
Abstract in Kuripaco. Supplementary information 1, http://www.raccefyn.co/index.php/raccefyn/article/downloadSuppFile/403/1749
Introduction
Rapatea Aubl. is the most widely distributed genus in the Rapateaceae (Boom, 1994). It currently comprises about 25 species of terrestrial herbs from 15 cm to 2 m tall, occurring in Panama, Colombia, Venezuela, Trinidad, Guyana, Suriname, French Guiana, Ecuador, Peru, Brazil and Bolivia (Monteiro, 2011). Species of Rapatea occur along streams, in the edges and shady understory of flooded (Rangel-Ch., 2008) and terra firme forests, riparian granitic outcrops (Giraldo-Cañas, 2008), and in the oligotrophic, seasonal flooded white-sand savannas associated with black-water rivers (Berry, 2004; Huber, 2005; 2006). Like the rest of the family, most of the species of Rapatea have radiated successfully in the Amazon basin and on the Guiana Shield (Ducke, 1938; Berry et al., 1995; Givnish et al., 2000, 2004; Bouchenak-Khelladi et al., 2014; Fernández-L. et al., 2016). Few species are found outside of this region, such as R. paludosa Aubl., the most widespread species of the genus (from Panama, Trinidad, and Venezuela to Bahia state in Brazil). Furthemore, Rapatea pycnocephala Seub. is known in central and southeastern Brazil (Mato Grosso do Sul: Xavantina; Bahia), and R. spectabilis Pilg. has been collected south to southern in Bolivia (Beni); R. elongata G. K. Schultze is common in northeastern of Colombia, an area that harbors some of the wettest forests in the world (Arellano-Peña & Rangel-Ch., 2004). This region is known as the biogeographical Chocó or "Chocó biogeográfico", and it is a broad strip of land with the highest biodiversity found between the Pacific Ocean and the slopes of the Western Cordillera (Cuatrecasas, 1989; Rangel-Ch., 2004). Rapatea is rare in the Andes, however two populations of R. muaju García-Barr. & L. E. Mora had been detected in Ecuador from collections made in the last 25 years in the foothills ("Piedemonte Andino"), the lower mountains (380-930 m) of Morona-Santiago Department, (see: http://www.tropicos.org/Name/27200122?tab=specimens), and in "Cerro Kaputna" in the Cordillera del Cóndor (Neill, et al., 2007). This region forms part of the "sub-Andean cordillera" made up principally of Mesozoic and early Tertiary sediments of sandstones and limestones (Rodríguez-R., et al., 2013) that form the border between Ecuador and Peru. The vegetation of these Cordillera have a phytogeographical connection with the Guayana Shield in northeastern South America (Neill, 2005). As a consequence, an important number of plant genera once thought to be endemic to the sandstone mountains (Tepuis) of the Guayana region have been found to occur disjunctly on the sandstone portions of the Andean cordilleras, but not elsewhere in the Andean region (Berry & Riina, 2005; Aymard & Campbell, 2008).
Rapatea is distinguished from other genera in the family by its laterally compressed inflorescences, which are subtended by two conspicuous involucral bracts that contain a capitate or elongate assemblage of flowers (Stevenson, et al., 1998; Stevenson, 2004). The anthers are tipped with a cochleariform appendage (Oriani & Scatena, 2013), and the carpels are uniovulate with basal placentation, with seeds that are oblong and exappendiculate, with longitudinal striations. A revision of the genus was published by Maguire (1965), who recognized 18 species and placed them in three sections (Elongata, Longipes, and Paludosa). Maguire employed three characters to separate the species: the base of leaf blades, the length of the bractlets of the spikelets, and the shape of the inflorescence receptacle. Since then, four new species and two varieties have been described (Maguire, 1979; Steyermark, 1988, Steyermark, et al., 1989; Boom, 1994). The genus has also been treated for "Flora de Venezuela" (Maguire, 1982), Flora of Ecuador (Berry, 1999), Flora of the Venezuelan Guayana (Berry, 2004), the checklist of the plants of the Guiana Shield (Berry, 2007), and lately in the "Catálogo de Plantas y Líquenes de Colombia" (Berry, 2016).
This new species increases to 12 the number of Rapatea species known from Colombia, and 25 for the genus. Five of these species are restricted to the Río Negro basin in the Amazonian region of Brazil, Colombia and Venezuela (R. angustifolia Spruce ex Körn., R. circasiana García-Barr. & L. E. Mora, R. isanae Aymard & Arellano-Peña, R. longipes Spruce ex Körn., and R. spruceana Körn.). This region harbors an array of plant families species that tolerate the nutrient-poor soils habitats derived by sandstone substrates such as the white sand savannas surrounded by Amazonian Caatinga ("Campinarana florestada"), as well as open areas like the Bana ("Campinas" in Brazil) vegetation, most often found near black-water rivers (Huber, 1988; Kubitzki, 1989; 1990; Berry & Riina, 2005).
Taxonomy. Rapatea isanae Aymard & Arellano-Peña, sp. nov. (Figures 1, 2, 3, y 4).
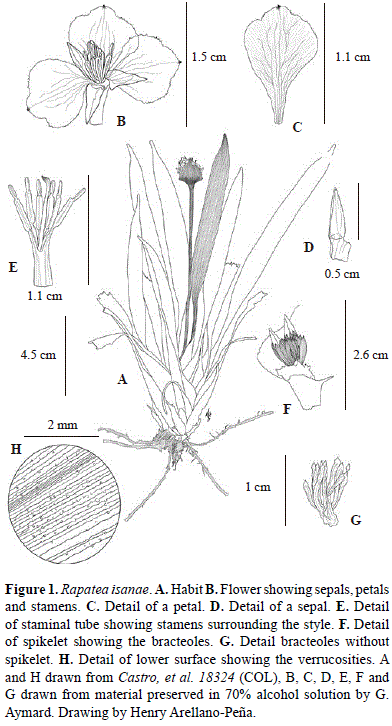



Type: COLOMBIA. Guianía: Panapaná, Corregimiento de Venado, alto río Isana, alrededores de la Comunidad Punta Tigre, 1° 4' 25.5048'' N; 69° 43' 58.0296'' W, 185 m, 7 de Mayo 2014, F. Castro-Lima, G. Aymard C., V. Minorta-C., H. Arellano-P., L. Niño, A. Lozano, M. González & C. Villegas 18324 (holotype: COL; isotype: COAH).
Diagnosis: Rapatea isanae differs from the morphologically most similar species R. spruceana Spruce ex Körn. in its shorter stature (15-30 cm tall), leaf of the lamina is shorter (8-13 cm long), the lower surface of the leaf sparse verrucose, the involucral bracts covered by shorter glandular trichomes in the basal portion outside, the attenuate portion 1-5 mm long, spikelets shortly pedicellate, bractlets not ciliate at the margins, petals white, broadly obovate, and the apical appendage of anthers brown.
Herb acaulescent, perennial, erect, 15-30 cm tall. Leaves 3 to 15 per plant, leaf sheaths reddish, glabrous, conduplicate, equitant and distichous, subcoriaceous to chartaceous, elongate, with about 50 distinct nerves, 5-9 cm long x 0.8-10 cm wide; base of leaf blades gradually passing into the petiole, petioles slender, 1.5-10 cm long, glabrous, narrowled toward the apex; leaf blades subcoriaceous, lanceolate, 5-13 cm long x 1-1.3 cm wide, glabrous on both sides, sparsely verrucose along the veins on the lower surface, with 12-15 secondary veins, the base attenuate, the apex long-acuminate, acumen ca. 1 cm long. Inflorescence 1-3 per plant, peduncle 10-15 cm long, 1-1.5 mm thick, glabrous, canaliculate, somewhat broadened and medially compressed at the summit below the head, head compressed, broadly ovate, subtended by 2 involucral bracts red ca. 1.5 x ca. 1.5 cm, coriaceous, covered by short glandular trichomes on the basal portion outside, glabrous inside, multinerved (ca. 100 nerves), receptacle convex, ca. 5 mm x 5 mm, the attenuate apex of the involucral bracts 1-5 mm long, spikelets 12-15 per inflorescence, pedicels ca. 1 mm long, bractlets 18-20 per spikelet, linear-oblanceolate, 7-10 mm x ca. 1 mm, 3-nerved, membranaceous, glabrous on both sides, shortly acuminate. Flower: sepals coriaceous, lanceolate-acuminate, 3 or 4-veined, 4-6 mm x ca. 2 mm, glabrous on both sides, bases membranous and basally connate into a tube 4-5 mm long, upper halves free; petals white, blade widely-obovate, membranous, 8-11 mm x 5-6, shortly apiculate at the apex, margins erose, 7-10-veined, the basal claws ca. 3 mm long, connate for 1-2 mm at the base, glabrous on both sides; stamens 6, the filaments 1.5-2.4 mm long, inserted near the base of the corolla tube, staminal tube ca. 3.5 mm long, anthers extrorse, white, 3-3.4 mm long with a brownish apical cochleariform linear-oblong appendage ca. 1.5 mm long, attached by a short, sigmoid base; ovary conical to subcapitate, ca. 2 mm x ca. 2 mm, style equaling the stamens, 9-10 mm long, the stigma curved, acute. Fruits and seeds not seen.
Distribution and habitat. In the edges and in shaded places of oligotrophic, flooded white-sand savannas from the upper Isana [Içana] river, Guianía Department, Colombia, at elevations of 185 m.
Taxonomic notes. Because of the spikelets bractlets of the of equal length and the inflorescence receptacle convex or hemispheric, Rapatea isanae belongs to Rapatea sect. Longipes Maguire (Maguire, 1965). By its base of leaf blades gradually passing into the petiole, apex of leaf blades not abruptly narrowed, the petiole shorter than the blade, the attenuate portion of the involucral bracts short (< twice as long as the expanded broader basal portion), and bractlets linear-oblong, it is most similar to R. spruceana Körn. from the Río Negro basin in the Amazonian region of Brazil, Colombia and Venezuela (Berry, 2004). However, R. isanae differs from the latter in its shorter stature 15-30 cm tall (vs. 30-80 cm tall), leaf blades 8-13 cm long, the lower surface sparse verrucose (vs. 15-30 cm, glandular-punctate), peduncles 10-15 cm long (vs. 16-35 cm), involucral bracts covered by shorter glandular trichomes in the basal portion outside, and the attenuate portion 1-5 mm long (vs. glabrous, 10-45 mm long), spikelets shortly pedicellate (vs. sessile), bractlets not ciliate at the margins (vs. ciliate), petals white, widely-obovate (vs. yellow, widely-ovate), and the apical appendage of the anthers brown (vs. purple).
In its leaf sheath shape, the convex inflorescences, and the petal color, R. isanae also has affinities with R. longipes Spruce ex Körn. and R. modesta Maguire, however, it can be distinguished from these species by the characters indicated in Table 1.
Etymology. Rapatea isanae is named after the Isana river. This river drains into the Rio Negro; geographically, its basin is nested between the Xié (to the north) and the Vaupés (to the south) river basins. Politically, the upper portion is in Colombia and the lower portion in Brazil (where it is called Içana). The headwaters of the Isana river are considered sacred land for the Kuripaco and Baniwa Indigenous groups, a nation that belongs to the Arawak language family that number about 14.000 people who has lived for centuries in the upper Río Negro area (Cabrera-Becerra, 2010). This culture believes that the center of the world ("umbilicus"), where all the human races began, is found in the pristine region of the upper Isana (Alveiro Calero-Cayopare, pers. comm., 2016). This ancient heritage clearly demonstrates that forests also provide an array of cultural and spiritual benefits to the people that occupy them. Notwithstanding, these compensations are only recently being explicitly considered as ecosystem services (Daniel, et al., 2012).
Phenology. Known to flower in May, but information from other times of the year is lacking.
IUCN Red List category. According the IUCN criteria this species would be ranges in DD (data deficient category). In this grade, more information is required and acknowledges that future research will show that threatened classification is appropriate (IUCN, 2016). Currently, R. isanae is known from only two collections, but we expect this new species to have a wider distribution in the upper Río Negro basin, an area poorly known botanically (Aymard & Castro-Lima, 2015).
Additional specimens examined (Paratype):
Colombia. Guianía: Corregimiento de Venado, Comu-nidad Punta Tigre: same locality and date as the type. M. González, F. Castro-Lima, G. Aymard C., V. Minorta-C., H. Arellano-P., L. Niño, A. Lozano & C. Villegas V. 742 (COL); F. Castro-Lima, G. Aymard C., Vladimir Minorta-C., H. Arellano-P., L. Niño, A. Lozano, M. González & C. Villegas V. 18291 (COL).
Rapatea modesta Maguire, Bot. Mus. Leafl. 14: 112. 1950. - Rapatea longipes Spruce ex Körn. var. modesta (Maguire) Maguire, Mem. New York Bot. Gard. 12(3): 100. 1965. Type: COLOMBIA: Comisaría del Amazonas, TrapécioAmazónico, interior regions of trapecio between the Amazon and Putumayo watersheds, ca. 100 m, Nov 1945, R. E. Schultes 6900 (holotype: NY, isotype: GH).
Taxonomic history. Rapatea modesta was described by Maguire based on three specimens collected in 1945 by Richard Evans Schultes during his explorations to the "Trapecio Amazónico", a broad trapezoid strip of land 150 km long between the Putumayo and Amazon rivers. Maguire made a detailed description but did not mention an affinity to his new taxon. Afterwards he wrote to R. E. Schultes and remarked that R. modesta is most closely related to R. longipes, from which it differs in its smaller habit, broader, shorter, more conspicuously veined membranous leaf-sheaths, relatively broader and more numerously veined leaf blades, considerably larger heads, more acuminate spiked bractlets, and white rather than yellow petals (Schultes, 1950). In his monograph of the genus Rapatea, Maguire referred this species to a variety of R. longipes (i.e., R. longipes var. modesta (Maguire) Maguire), also providing a key to separate both varieties (Maguire, 1965). Examination of type specimen, the study of additional material, and Maguire's data clearly indicates that R. modesta should be restablished and treat as distinct species. Because of its bractlets of the spikelets of equal length and the inflorescence receptacle convex or hemispheric, this species belongs to Rapatea sect. Longipes Maguire, and by the base of leaf blades abruptly constricted above the summit of the sheath forming a long and slender petiole, R. modesta is most similar to R. longipes, however, these two species can be distinguished by the characteristics presented in Table 1.
The label of on R. E. Schultes 6899* states (handwritten, but the label data) "red flowers" for R. modesta Maguire. Schultes observed that the red-flowered form grows together with the white-color form (Schultes, 1950). According to Maguire (1950, 1982), as well as hundreds of specimens examinated, red flowers have not been reported for the genus besides this Schultes's collection (Paul Berry, pers. comm., 2016). We are not questioning Schultes's field observations, but perhaps he was referring to the reddish involucral bracts, which can indeed be reddish, rather than to the petals themselves.
Additional specimens examined:
Colombia. Amazonas: TrapécioAmazónico, interior regions of trapecio between the Amazon and Putumayo watersheds, ca. 100 m, Nov 1946, G. A. Black & R. E. Schultes 46-361 (NY); ibid, flowers red, Nov 1945, R. E. Schultes 6899*(COL, GH, K, NY). P. N. N. Río Puré, Varillal de Aguanegra, 2° 02' 52.44''S; 69° 41' 33.96'' O, July 2010, A. Barona 46 (COAH); La Pedrera, La Tonina, 1° 29' S; 69° 29' O, 10 Oct 1994, D. Cárdenas, D. Giraldo C. & E. Yukuna 5750 (COAH).Vaupés: Río Apaporis, raudal Jirijirimo, Aug 1951, R. E. Schultes & I. Cabrera 13467 (GH); Río Piraparana, Cerro E-ree-eé-ko-mee-o-kee. 0° 19' 60'' N; 70° 30' 00'' W, Sep 1952, R. E. Schultes & I. Cabrera 17500 (COL); Río Piraparana, Misión San Miguel, Oct 1976, E. W. Davis 127 (COL).
Rapatea sessiliflora (Maguire) Aymard & Arellano-Peña, comb. et stat. nov.: Rapatea paludosa Aubl. var. sessiliflora Maguire, Bull. Torrey Bot. Club 75: 204. 1948. Type: SURINAME: Sipaliwini District: Tafelberg (Table Mountain), Grace Creek, 1 kilometer east of Savanna VIII, 03° 54' N; 56° 20' W, ca. 1000 m, 01 Sep 1944, B. Maguire 24587 (holotype: NY, isotypes: A, F, U, US).
Rapatea sessiliflora was described as a variety of R. paludosa based on two specimens (Maguire, 1948). The examination of type specimen and the additional material indicates that it should be treated as a separate species, which can be distinguished from R. paludosa by the following characters: slender plants (vs. stout), leaf blades 40-60 cm long. 1-2.5(3.5) cm wide (vs. 60-90 cm long, 4-12 cm wide), inflorescences 3-4 cm broad (vs. 4-8 cm broad), apex of the bracts ca. 10 cm long, 2.0-2.5(3.2) cm wide at the base (vs. 15-20 cm long, 4-6 cm wide), and spikelets sessile or with pedicels 0.5-3 mm long (vs. spikelets with pedicels 5-12(20) mm long).
Additional specimens examined. BRAZIL: Amazonas, In vicinibus Barra (Manaus), December-March 1850-51, R. Spruce s.n. (NY). Amazonas, vicinity of Manaus, Tarumã, Cachoeira baixa Tarumã, June 1967, G. T. Prance, B. S. Pena & J. F. Ramos 3859 (INPA, NY). SURINAME: base of Tafelberg Mountain, Coppename River headwaters, 106 m, July 1944, B. Maguire 24177 (NY); Tafelberg, Savanna landing strip, March 1961, K. U. Kramer & W. H. A. M. Hekking 3029 (NY, U); Sipaliwini District: Tafelberg National Park, western side of Lisa Creek, 3° 55' N; 56° 11' W, ca. 1000 m, June 1998, L. G. Lohmann 207 (MO, WIS); Tafelberg Mountain, Grace Creek, 1 km north of Grace fall, 3° 54' 32'' N; 56° 12' 44'' W, ca. 800-820 m, July 1998, T. Hawkins 1870 (MO, WIS). VENEZUELA: Bolívar: ríos Icabarú y Hacha, March 1955, L. A. Bernardi s.n (MER, NY).
Key to the species of Rapatea (Modified from Berry, 2004). Supplementary information 2, http://www.raccefyn.co/index.php/raccefyn/article/downloadSuppFile/403/1750.
1. Leaf blades 0.5-2.5 cm wide....................................2
2(1). Receptacle plane-convex; bractlets strongly gradate............R. linearis Gleason (Amazonian of Brazil, Colombia, Guyana)
2. Receptacle shallowly convex, at least hemispheric or elongate, bractlets equal in length........................................3
3(2). Leaves blades orange punctate below, inflorescence elongate, base of the involucral bract deeply cordate........R. xiphoides Sandw. (Guyana: Kaieteur savannas)
3. Leaves blades if punctate, without orange dots, inflo-rescence shallowly convex or at least hemispheric, base of the involucral bract truncate, rounded or subcordate............4
4(3). Base of leaf blades abruptly widened at the base above the petiole; apex of leaf blade abruptly narrowed; petiole as long as the blade or longer...................R. longipes Spruce ex Körn. (Río Negro basin: Amazonian of Brazil, Colombia and Venezuela)
4. Base of leaf blades gradually passing into the petiole; apex of leaf blades not abruptly narrowed; petiole shorter than the blade.......................................................................5
5(4). Apex of the involucral bracts attenuate 1-45 mm long, < twice as long as the expanded broader basal portion; lower surface of the leaf blade glandular-punctate or sparse verrucose mainly on the lower surface.................................6
6. Plants 30-80 cm tall; lower surface of the leaf blade glandular-punctate; attenuate portion of the involucral bracts 10-45 mm long; peduncle of inflorescence 16-35 cm long, petals yellow.............R. spruceana Körn. (Río Negro basin: Amazonian of Brazil, Colombia and Venezuela)
6. Plants 15-30 cm tall, lower surface of the leaf sparsely verrucose; attenuate portion of the involucral bracts 1-5 mm long, peduncle of inflorescence 8-13 cm long; petals white..........R. isanae Aymard & Arellano-Peña (Río Negro basin: Amazonian Colombia)
5. Attenuate portion of the involucral bracts 20-80 mm long, 2-4 times longer than the expanded broader basal portion; lower surface of the leaf blade smooth (not glandular-punctate or sparsely verrucose)............................7
7(5). Attenuate portion of the involucral bracts 20-35 mm long, 2-3.5 times longer than the expanded broader basal portion; lower surface of the leaf blade densely and minutely papillate; bracteoles aristate...............R. yapacana Maguire (Venezuela, Amazonas state: savanna Yapacana)
7. Attenuate portion of the involucral bracts 35-80 mm long, 4-5 times longer than the expanded basal portion; lower surface of the leaf blade not minutely papillate; bracteoles acute, apiculate, or sharply acuminate................8
8(7). Plants slender; expanded basal portion of involucral bracts 7-12 mm wide; attenuate portion of involucral bracts 35-50 mm long, 4-5 mm broad; leaf blades 3-7 mm wide; peduncle 4-20 cm long, shorter than the leaves; plants growing in dense circular clumps of numerous individuals........R. angustifolia Spruce ex Körn. (Río Negro basin: Amazonian of Brazil, Colombia and Venezuela)
8. Plants stout; expanded basal portion of involucral bracts 20-25 mm wide; attenuate portion of involucral bracts 55-85 mm long, 9-14 mm broad; leaf blades 9-15 mm wide; peduncle 35-45 cm long, longer than, or equal to, the leaves; plants generally growing as solitary individuals........R. circasiana García-Barr. & L. E. Mora (Río Negro basin: Amazonian of Brazil, Colombia and Venezuela)
1. Leaf blades 3-14 cm wide........................................9
9(1). Leaf sheath short, 2-9 (-15) cm long; involucral bracts 1.5-8 cm long, apex narrowed, 0.5-3.5 cm long; heads 1-3 cm broad...........................................................10
10(9). Base of leaf blades gradually narrowed at the base to a union with the sheath, petiolar portion ca. 0.2 cm long; involucral bracts 6-8 cm long, narrowed apical part 3-3.5 cm long..................Rapatea sp. A (Colombia; Cauca)
10. Base of leaf blades abruptly widened at the base above the petiole; portion more than 1 cm long; involucral bracts 1.5-3.5 cm long, narrowed apical part 0.5-2.5 cm long......11
11(10). Leaf sheaths lanceolate, 3 or more times longer than broad; petiole as long as the blade, leaf blades 3-3.5 cm wide, oblong-linear to lanceolate; narrowed apical part of the involucral bracts 0.5-1 cm long; peduncle 12-25(30) cm long; petals yellow...................................R. longipes Spruce ex Körn. (Río Negro basin: Amazonian Brazil, Colombia and Venezuela)
11. Leaf sheaths strongly ventricose, 2-3 times longer than broad; petiole longer than the blade; leaf blades 3-6 cm wide, oblong-elliptic; apex 2-3 cm long; narrowed apical part of the involucral bracts 1-2.5 cm long; inflorescence 6-15 cm long; petals white.................................R. modesta Maguire (Amazonian Colombia)
9. Leaf sheath elongate, 15-30 cm long; involucral bracts 5-25 cm long, apical attenuate portion 3-22 cm long; heads 3-8 cm broad...........................................................12
12(9). Lamina base gradually narrowed to petiole region..............................................................................13
13(12). Petiolar region 0.5-3 cm long.........................14
14(13). Leaves (4-) 5-9 cm wide; leaves smooth throughout; involucral bracts cordate at the base; bracteoles acuminate, spotted (maculate) at the apex........R. spectabilis Pilg. (Brazil, Bolivia, Colombia, Ecuador and Peru)
14. Leaves 3-4 cm wide; lower surface of the leaves slightly scabridulous; involucral bracts truncate at the base; bracteoles cucullate-obtuse, without spots at the apex...... R. scabra Maguire (Venezuela, Amazonas state: Cerro Sipapo)
13. Petiolar region more than 3.5 cm long...................15
15(13). Leaves 4-12 cm wide, inflorescence 4-8 cm broad, involucral bracts 10-20 cm long, cordate, spikelets pedicellate (pedicels 10-20 mm long), widespread in the Neotropics (from Panama, Trinidad, Venezuela to Bolivia)................R. paludosa Aubl.
15. Leaves 3-4 cm wide, inflorescence ca. 3 cm broad, involucral bracts 8-10 cm long, involucral bracts rounded or deeply cordate at the base, spikelet sessile or pedicels not longer than 3 mm (Brazil, Guayana Shield of Suriname and Venezuela)..........................................................................16
16(15). Involucral bracts deeply cordate at the base; spikelet bractlets 5(-7) nerved at the apex........................R. pycnocephala Seub. (Brazil: Bahia, Goiás, Maranhão, Mato Grosso, Pará, Piauí, Rondônia)
16. Involucral bracts rounded at the base; spikelet bractlets 3-nerved at the apex......................R. sessiliflora (Maguire) Aymard & Arellano-Peña (Brazil-Amazonas, Suriname and Venezuela)
12. Base of leaf blades abruptly constricted above the summit of the sheath...........................................................17
17(12). Floral receptacle elongate...............................18
18(17). Leaf blades strongly rugulose; inflorescences 5-14 cm long......................................................................19
19(18). Leaves blades coriaceous, narrower-lanceolate, ca. 3.5 cm long; petiole 8-10 cm long: head 5-8 cm long, bracteoles scarious-membranaceous...................R. rugulosa Maguire (Brazil: Amazonas state)
19. Leaves blades membranaceous, linear-lanceolate, 6-10 cm wide; sessile, head 6-12 cm long, bracteoles coriaceous...................R. membranacea Maguire (Guyana: Pakaraima mountains)
18. Leaf blades smooth; inflorescences 4-8 cm long.....20
20(18). Leaf blades 1.5-3 cm wide, with 14-16 nerves, orange punctate; receptacle 1.5-2 cm; involucral bracts 15-25 x 1.5-2 cm, cordate, inflorescence ovoid-spherical, 3.5-4 x 3-3.5 cm...................R. xiphoides Sandw. (Guyana: Kaieteur Savannas)
20. Leaf blades 3-5 cm wide, with 10-12 nerves, if punctate, without orange dots; receptacle 5-19 cm long; involucral bracts 20-37 x 3-5 cm, rounded or attenuate, inflorescence oblong, 7-8.5 x 5-6 cm.......... R. elongata G. K. Schultze (Brazil and Colombia)
17. Floral receptacle convex or hemispheric...............21
21(17). Leaves blades oblong, abruptly short-acuminate at the apex, constricted at the base into an evident petiole of 1-20 cm long.................................................................22
22. Leaves blades coriaceous, petiolar portion 10-20 cm long; bracteoles rigid, 3-7-nerved, short-acuminate.........R. muaju García-Barr. & L. E. Mora (Brazil-Acre, Colombia, Ecuador and Peru)
22(21). Leaves blades membranaceous; petiolar portion 1-10 cm long; bracteoles membranaceous, 3-5-nerved, long-aristate.......................................................................23
23(22). Leaves blades ca. 50 x ca. 10.5 cm; base subcordate to oblique; petiolar portion 8-10 cm long; penducle of the inflorescence 18-25 cm long; bracteoles 5-nerved........................R. saulensis Boom (French Guiana and Suriname)
23. Leaf blades 16-21 x 4-8 cm; base inequalateral; petiolar portion 1-3 cm long; penducle of the inflorescence 4-5 cm long; bracteoles 3-5-nerved.................R. undulata Ducke (Amazonian of Brazil, Colombia and Peru)
21. Leaf blades lanceolate, gradually acuminate at the apex, cordate or attenuate at the base, with a shorter petiole, 5-20 mm or less of long....................................................24
24(21). Leaf blades smooth; inflorescences elongate-hemispheric; bracteoles not all gradate..............................25
25(24). Leaf blades 33-38 x 5-6 cm, leaf sheaths 10-14 cm long, ventricose......R. ulei Pilg. var. ulei (Amazonian of Brazil, Guyana, French Guiana and Peru)
25. Leaves blades 40-60 x 10-14 cm, leaf sheaths 20-30 cm long, elongate....................................................26
26(25). Inflorescence uni-capitate..............R. ulei Pilg. var. latifolia Maguire (Brazil, Amazonas state)
26. Inflorescences multicapitate...................R. ulei Pilg. var. latifolia Maguire fm. multicapitata Maguire (Brazil, Amazonas state)
24. Leaves blades glandular-papillate; inflorescences shallow-hemispheric to sub-hemispheric; bracteoles evi-dently gradate.....................................................................27
27(24). Peduncle 3-14 cm long; involucral bracts broadly deltoid, nearly as broad as long, 6-10 cm long..................28
28(27). Lower surface of leaf sheath densely brown-pubescent with elongate, crisp-villosulous, loose, multi-cellular hairs; upper surface of leaf with short, divaricate, pointed hairs irregularly scattered on the veins in addition to the minutely papillate surface; spikelets 16-20 mm long (excluding the sepals); lowest bracteoles 12-13 mm long; anthers ca. 5 mm long; filaments glabrous.......R. steyermarkii Maguire (Guayana Shield of Guyana and Venezuela)
28. Lower surface of leaf sheath brown-dotted between smaller, pale punctuations; upper leaf surface densely verruculose-papillate throughout; spikelets 23-25 mm long (excluding the sepals); lowest bracteoles 17-18 mm long; anthers ca. 10 mm long; filaments ciliolate.................R. aracamuniana Steyerm. (Venezuela, Amazonas state: Cerro Aracamuni)
27. Peduncle 20-50 cm long; involucral bracts lanceolate, longer than broad, 10-25 cm long...............................29
29(27). Bracteoles stiff, obviously different in size, the lower ones much less than 1/2 the length of the spikelet; leaf blades 25-75 cm long, 6-11 cm wide; involucral bracts 10- 20 cm long; lower leaf surface with the primary veins acutely elevated, separating the broad, flat depressions; primary veins of upper leaf surface shallowly raised, not presenting a plaited appearance, 0.5-1 mm apart................R. fanshawei Maguire (including R. fanshawei var. minor) Guayana Shield of Guyana and Venezuela
29. Bracteoles sub-membranaceous, of nearly equal length; leaf blades ca. 130 cm long, 9-12 cm wide; involucral bracts ca. 25 cm long; lower leaf surface with shallowly convex surfaces separated by shallow sulcations, lacking acutely elevated primary veins; primary veins of upper surface conspicuously elevated and presenting a plaited appearance, 1-2 mm apart............R. chimantensis Steyerm. (Venezuela, Bolívar state: Chimantátepui)
Acknowledgments
We gratefully acknowledge the logistic support provided by Compensation International Progress S.A.-Ciprogress Greenlife- staff: Germán Bernal-G., Alicia Micolta-C., Daniel S. Bernal-L., and the Kuripaco indigenous group in the course of a botanical expedition to the Cuyarí and Isana rivers sponsored by the "Flor de Inírida" REDD+ project of the Guayana-Amazonian transition region. We also thank, Orlando Cordubí and Lino Flores for their field assistance, the Kuripaco people of the communities of Cejal, Amanadona and Campo Alegre (Cuyarí river) for their extraordinary effort carrying hundreds of kilograms of equipment and food along ancestral trails between the cañoNakén and ríoCuyarí, and the "raudal" Yuruparí along the ríoCuyarí. To Paul E. Berry (MICH) for reviewing many times the manuscript, for his constructive feedback, and for his help improving the key to species of Rapatea. We also thank Lisa M. Campbell (NY) for reviewing the manuscript, to Gustavo Romero-G. (AMES) for reviewing the manuscript and for his help in finding updated literature, and to Carlos Parra-O. (COL) and Dairón Cárdenas-L. (COAH) for their herbarium assistance. Finally, we wish to thank Albeiro Calero Cayopare, a native expert in the Kuripaco language (from Tonina, a village along the Guianía river) for his help in translating the abstract into Kuripaco, and for providing information about the upper Isana River.
Conflicts of interest
The authors declare that they have no conflict of interest.
References
Arellano-Peña, H. y Rangel-Ch, J. O. (2004). Clima del Chocó biogeográfico/Costa pacífica de Colombia. Pp. 39-82. Colombia Diversidad Biótica IV. El Chocó biogeográfico/ Costa Pacífica. En: Rangel-Ch. J. O. (editor), Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Aymard, G. & Castro-Lima, F. (2015). A second tree species of Ampelozizyphus (Rhamnaceae), from the upper Cuyarí river basin, Guianía (Colombia). Harvard Papers in Botany 20: 161-166. [ Links ]
Aymard G. & L. Campbell. (2008). A new species of Caraipa Aublet (Clusiaceae) from the Venezuelan Andes and its biogeographical implications. Rodriguesia 59 (2): 1-6. [ Links ]
Berry, P. E. (1999). 211B. Rapateaceae. Flora of Ecuador 63: 37-46. [ Links ]
Berry, P. E. (2004). Rapateaceae. Flora of the Venezuelan Guayana 8: 413-472. [ Links ]
Berry, P. E. (2007). Rapateaceae. Checklist of the Plants of the Guiana Shield (VENEZUELA: Amazonas, Bolivar, Delta Amacuro; GUYANA, SURINAM, FRENCH GUIANA). Contributions from the United States National Herbarium 55: 173-174. [ Links ]
Berry, P. E. (2016). Rapateaceae. Pp. 2238-2241. Catálogo de Plantas y Líquenes de Colombia. En: Bernal, R., Gradstein, S. R. y Celis, M. (editores). Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. Colombia. http://catalogoplantasdecolombia.unal.edu.co [ Links ]
Berry, P. E. & Riina, R. (2005). Insight into the diversity of the Pantepui flora and the biogeographic complexity of the Guayana shield. Biol. Skr. 55: 145-167. [ Links ]
Berry, P. E., Huber, O. & Holst, B. K. (1995). Floristic analysis and phytogeography. Pp. 161-192. In: Flora of the Venezuelan Guayana, Vol. 1. (Introduction) Berry, P. E. & Holst, B. K. (Editors). Timber Press, Portland. [ Links ]
Boom, B. M. (1994). A new species of Rapatea section Paludosa (Rapateaceae) from French Guiana. Brittonia 46: 314-316. [ Links ]
Bouchenak-Khelladi, Y., Muasya, A. M. & Linder, H. P. (2014). A revised evolutionary history of Poales: origins and diversification. Botanical J. of the Linnean Society 175: 4-16. [ Links ]
Cabrera-Becerra, G. (2010). El Geral y la colonización en el alto Río Negro-Vaupés. Pp. 365-390. En: Perspectivas antropológicas sobre la Amazonia contemporánea. Chaves-Chamorro, M. & Del Cairo-Silva, C. L. (Editores). Instituto Colombiano de Antropología y Pontificia Universidad Javeriana. Bogotá, Colombia. [ Links ]
Cuatrecasas, J. (1989). Aspectos de la vegetación natural de Colombia. Pérez Arbelaezia. 2: 155-283. [ Links ]
Daniel, T. C., Muhar, A., Arnberger, A., Aznar, O., Boyd, J. W., Chan, K. M. A., Costanza, R., Elmqvist, T., Flint, G. C., Gobster, P. H., Grêt-Regamey, A., Lave, R., Muhar, S., Penker, M., Ribe, R. G., Schauppenlehner, T., Sokor, T., Soloviy, I., Spierenburg, M., Kaczanowska, K., Tam, J. & von der Dunk, A. (2012). Contributions of cultural services to the ecosystem services agenda. Proc. Natl Acad. Sci. USA 109: 8812-8819. [ Links ]
Ducke, A. (1938). A Flora do Curicuriarí, afluente do Rio Negro, observada em viagens com a comissão demarcadora das fronteiras do setor Oeste. In: Anais da Primeira Reuniao Sul-Americana de Botanica. Vol 3, parte 1: 389-398. Rio de Janeiro. [ Links ]
Fernández-Lucero, M., Madriñán S. & Campbell, L. M. (2016). Morphology and anatomy of Guacamaya superba (Rapateaceae ) and Schoenocephalieae with notes on the natural history of the Flor de Inírida. Harvard Paper in Botany 21: 105-123. [ Links ]
Giraldo-Cañas, D. (2008). Flora vascular de los afloramientos precámbricos (Lajas-Inselbergs) de la Amazonia Colombiana y áreas adyacentes del Vichada: I. Composición florística. Pp. 89-118. En: Diversidad Biótica VII (Vegetación, palinología y paleoecología de la Amazonía colombiana). Rangel-Ch. J. O (editor). Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Givnish T. J., Evans, T. M., Pires, J. C. Zjhra, M. L., Patterson, T. B., Berry, P. E. & Sytsma, K. J. (2000). Molecular evolution, adaptive radiation, and geographic diversification in the amphiatlantic family Rapateaceae: evidence from ndhF sequences and morphology. Evolution 54: 1915-1937. [ Links ]
Givnish, T. J., Millam, K. C., Evans, T. M., Hall, J. C., Pires, J. C., Berry, P. E. & Sytsma, K. J. (2004). Ancient vicariance or recent long-distance dispersal? inferences about phylogeny and south American-African disjunctions in Rapateaceae and Bromeliaceae based on ndhf sequence data. Int. J. Plant Sci. 165 (4 Suppl.): S35-S54. [ Links ]
Huber, O. (1988). Guayana highlands versus Guayana lowlands, a reappraisal. Taxon 37: 595-614. [ Links ]
Huber, O. (2005). Diversity of vegetation types in the Guayana region: An overview. Biol. Skr. 55: 169-188. [ Links ]
Huber, O. (2006). Herbaceous ecosystems on the Guayana Shield, a regional overview. J. of Biogeography 33: 464-475. [ Links ]
IUCN. (2016). Guidelines for using the IUCN red list categories and criteria. Version 12. Prepared by the standards and petitions Subcommittee. IUCN, Gland, Switzerland and Cambridge, England. http://www.iucnredlist.org/documents/RedListGuidelines.pdf . [ Links ]
Kubitzki, K. (1989). Amazon lowland and Guayana highland. Historical and ecological aspects of their floristic development. Rev. Acad. Colomb. Cienc. Ex. Fis. Nat. 17 (65): 271-276. [ Links ]
Kubitzki, K. (1990). The psammophilous flora of northern South America. Mem. New York Bot. Gard. 64: 248-253. [ Links ]
Maguire, B. (1948). Plant Explorations in Guiana in 1944, chiefly to the Tafelberg and the Kaieteur Plateau-II. Bull. Torrey Bot. Club. 75: 189-230. [ Links ]
Maguire, B. (1965). Rapateaceae (The botany of the Guayana Highland-Part VI). Mem. New York Bot. Gard. 12: 69-102. [ Links ]
Maguire, B. (1979). Additions to the Rapateaceae. Acta Amazonica 9: 267-269. [ Links ]
Maguire, B. (1982). Rapateaceae. En: Luces de Febres, Z. & Steyermark, J. A. (editores). Flora de Venezuela 11 (2): 85-203. [ Links ]
Monteiro, R. F. (2011). Neotropical Rapateaceae. In: Neotropikey - Interactive key and information resources for flowering plants of the Neotropics. Milliken, W., Klitgård B. & Baracat, A (2009 onwards), Kew Gardens, UK. http://www.kew.org/science/tropamerica/neotropikey/families/Rapateaceae.htm [ Links ]
Neill, D. A. (2005). Cordillera del Cóndor: Botanical treasures between the Andes and the Amazon. Plant Talk 41: 17-21. [ Links ]
Neill, D. A. H. van der Werff, S. Churchill, T. Croat, M. Asanza, W. Quizhpe, C. Morales, D. Reyes, J. Manzanares, T. Katan, C. Kajekai, A. W. E. Rodríguez, J. Perea, J. Gruhn & P. Almeida. 2007. Inventario Botánico de la región de la Cordillera del Cóndor, Ecuador y Perú: Actividades y resultados científicos del proyecto, 2004- 2007. Informe Fundación Nacional de Ciencias (NSF), Proyecto # 0346679, USA, 47 p. [ Links ]
Oriani, A. & Scatena, V. L. (2013). The taxonomic value of floral characters in Rapateaceae (Poales-Monocotyledons). Plant Syst. Evol. 299: 291-303. [ Links ]
Rodríguez-R., E. F., D. Neill, E. Alvítez-Izquierdo, S. J. Arroyo-Alfaro & J. A. Núñez-Zapata. (2013). Estado actual de los estudios de la flora en la Cordillera del Cóndor y áreas adyacentes en el Perú, 2006. Sagasteguiana 1 (1): 101-120. [ Links ]
Rangel-Ch. J. O. (2004). La vegetación del Chocó biogeográfico de Colombia y zonas cordilleranas aledañas - síntesis. Pp. 769-815. En: Colombia Diversidad Biótica IV. El Chocó biogeográfico /Costa Pacífica. Rangel-Ch., J. O. (editor). Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Rangel-Ch. J. O. (2008). La vegetación de la región Amazónica - aproximación inicial. Pp. 1-57. En: Colombia Diversidad Biótica VII. Vegetación, palinología y paleoecología de la Amazonía Colombiana. Rangel-Ch., J. O. (editor). Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Schultes, R. E. (1950). Plantae austro-americanae VI. Plantarum principautes. Vallis amazoniae novarum vel minus cognitarum notae diversae. Bot. Mus. Leafl. 14: 109-157. [ Links ]
Stevenson, D. W., Colella, M. & Boom, B. M. (1998). Rapateaceae. Pp. 415-424. In: The families and genera of vascular plants Vol. 4. Kubitzki, K., Huber, H., Rudall, P. J., Stevens P. S. & Stützel T. (editors). Springer-Verlag, Berlin. Germany. [ Links ]
Stevenson, D. W. (2004). Rapateaceae. Pp. 477-479. In: Flowering Plants of the Neotropics. Smith, N. P., Mori, S. A., Henderson, A., Stevenson, D. W. & Heald, S. V. (editors). The New York Botanical Garden, Princeton University Press, Princeton. NJ. [ Links ]
Steyermark, J. A. (1988). Flora of the Venezuelan - IV. Ann. Missouri Bot. Gard. 75: 311-351. [ Links ]
Steyermark, J. A., Holst, B. K. & collaborators. (1989). Flora of the Venezuelan Guayana - VII. Contributions of the flora of the Cerro Aracamuni, Venezuela. Ann. Missouri Bot. Gard. 76: 945-992. [ Links ]

