Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Desde hace dos siglos naturalistas y geólogos, como von Humboldt, Wallace y Darwin, han emprendido viajes alrededor del mundo con el propósito de entender el origen, la distribución y la diversificación de las especies a lo largo de gradientes latitudinales y altitudinales, y de observar los patrones que han permitido el desarrollo de importantes teorías en los campos de la ecología y la evolución (Fischer, et al., 2011). Sus registros han servido para documentar cómo la diversidad de las especies aumenta a medida que disminuye la latitud, lo que se refleja en el Neotrópico, donde se alberga el 37 % de las plantas del mundo (90.000 a 110.000 especies de plantas con semillas), con una diversidad mayor que la de las dos regiones del Paleotrópico combinadas (Antonelli & Sanmartín, 2011). Desde el punto de vista de la altitud, se han descrito cuatro tendencias principales de distribución de las especies a lo largo de un gradiente: el patrón de disminución de riqueza, es decir, del número de especies al aumentar la elevación; el patrón de meseta baja, con una gran riqueza en la porción consecutiva baja del gradiente (hasta los 300 m), que decrece al aumentar la altitud; el patrón de meseta baja con pico de riqueza en la mitad del dominio, en donde la riqueza es alta por debajo de los 300 m, pero el pico se alcanza en la mitad del gradiente y de ahí decrece; y, por último, el patrón de pico en altitud media, con un máximo de diversidad unimodal en elevación intermedia. (superior a 300 m) con un 25 % adicional de especies comparada con la base y la cima de la montaña (McCain & Grytnes, 2010).
Estos patrones de distribución altitudinal son mediados por distintos factores, clasificados por Antonelli & Sanmartín (2011) en dos grandes grupos: bióticos (adaptación al suelo, interacción con polinizadores, dispersión y herbívoros, conservación del nicho) y abióticos (relieve, energía disponible, tiempo, precipitación, temperatura, área, estacionalidad, tipo de suelo, frecuencia y grado de perturbación, cambios hidrológicos y geológicos). Estos factores varían a lo largo de una montaña de manera no aleatoria y, por lo tanto, predecible, y su interacción determina el patrón de distribución de la riqueza de especies observado en cada caso (Lomolino, 2001; McCain & Grytnes, 2010). De hecho, en un estudio realizado con base en una recopilación de datos de Gentry (1982), Phillips & Miller (2002) aseguraron que los análisis estadísticos multivariados evidenciaban que los niveles de diversidad podían pronosticarse con una alta confiabilidad a partir de diversas variables ambientales (los sitios con mayor riqueza son las áreas ecuatoriales, bajas y húmedas), siendo la precipitación el factor abiótico más estrechamente correlacionado con la diversidad de los bosques tropicales (Gentry, 1982; Kreft & Jetz, 2007).
El Chocó biogeográfico es una región interesante para desarrollar este tipo de estudios, ya que es una ecorregión de la América tropical con una gran diversidad, concentración de especies endémicas y pérdida rápida de hábitat, lo que la convierte en uno de los veinticinco puntos calientes de biodiversidad en el mundo, por lo cual muchos esfuerzos se han orientado hacia la conservación del 24,2 % que aún se conserva de la extensión original de esta zona (Myers, et al., 2000). La mayoría de los especialistas coinciden en que esta zona abarca desde la provincia del Darién, al occidente de Panamá, hasta la provincia de Manabí, en la costa noroccidental de Ecuador y a lo largo de toda la costa colombiana del Pacífico. El límite occidental está definido claramente por el océano Pacífico y, según algunos autores, el límite oriental está demarcado por las estribaciones occidentales de la cordillera de los Andes, en tanto que otros lo sitúan en la línea divisoria de aguas de los Andes occidentales en Colombia y Ecuador (Otero, et al., 2009).
En Colombia, esta región ocupa territorios de los departamentos del Chocó, Antioquia, Valle del Cauca, Risaralda, Cauca y Nariño, con una extensión aproximada de 113.000 km2, lo que representa casi el 70 % de toda la ecorregión y el 10 % del territorio colombiano (Otero, et al., 2009). Esta zona se caracteriza por presentar elevaciones desde el nivel del mar hasta aproximadamente los 3.500 m, niveles muy variables de precipitación y temperatura, con medias anuales de 9.000 mm y 27 ºC, lo que permite la existencia de una gran diversidad de ecosistemas como manglares, terrazas aluviales, bosques pluviales tropicales, y bosques premontanos y montanos (Kelleher, et al., 1995). Desde la perspectiva de la diversidad y composición florística, esta ecorregión es una de las menos conocidas del país y el promedio de nuevas especies vegetales por descubrir es mucho más elevado que el de otras (Gentry, 1986; Bernal, et al., 2015); sin embargo, presenta altas tasas de deforestación por la tala indiscriminada de árboles, el cambio del uso de la tierra, la contaminación doméstica, la minería ilegal y los problemas sociales (Rangel & Lowy, 1993; Bernal, et al., 2015).
Con el objetivo de determinar los patrones de distribución de las comunidades vegetales en un rango altitudinal, y relacionarlos con las variables climáticas de temperatura y precipitación, en este estudio se caracterizaron cinco comunidades vegetales del Chocó biogeográfico del Valle del Cauca según su fisionomía y su estructura; además, se tuvieron en cuenta otras localidades previamente muestreadas en el Chocó biogeográfico de Colombia y Ecuador.
Materiales y métodos
Área de estudio. El estudio se hizo en cinco localidades ubicadas a diferentes altitudes en el Chocó biogeográfico del Valle del Cauca, Colombia (Tabla 1), las cuales se eligieron por su buen estado de conservación, pues hacen parte del sistema de Parques Nacionales Naturales o reservas regionales.
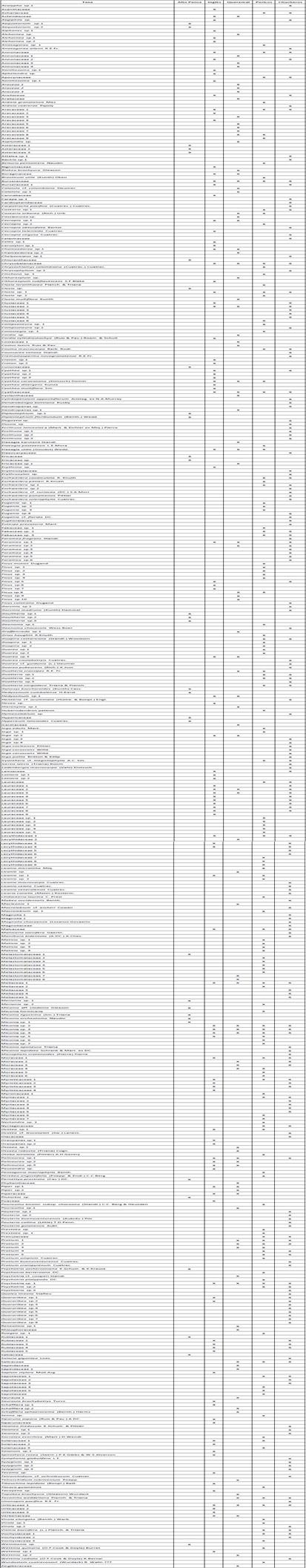
Tabla 1 Localidades muestreadas en este estudio (EE) y tomadas de la literatura (TL) ubicadas entre los 36 y los 3.650 m de altitud en el Chocó biogeográfico de Colombia y Ecuador (Gentry, 1988; Kappelle & Brown, 2001)
Asimismo, con el objetivo de mejorar la precisión de los modelos de regresión para la riqueza de especies, se utilizó la información de siete localidades de Colombia y Ecuador ubicadas dentro del Chocó biogeográfico, que cumplían con las mismas condiciones de muestreo de este estudio (ubicación geográfica, área muestreada y método de muestreo), y reportadas en los estudios de Phillips & Miller (2002) y Kappelle & Brown (2001).
Estructura y diversidad vegetal. Se utilizó el método de Gentry (1982), según el cual se traza un transecto de cinturón dividido en diez parcelas rectangulares consecutivas de 50 × 2 m, para un total de 0,1 hectáreas muestreadas en cada localidad. Se escogió el hábitat en el mejor estado de conservación posible para establecer el transecto. El muestreo incluyó toda la vegetación con diámetro a la altura del pecho (DAP) mayor o igual a 2,5 cm. Este método excluye la mayoría de hierbas y epífitas, pero incluye árboles, arbustos, lianas, hemiepífitas y hierbas grandes. Se midió la circunferencia a la altura del pecho (CAP) a 1,3 m de altura sobre el suelo usando una cinta métrica. Los tallos ramificados a menos de 1,3 m del suelo se midieron como individuos independientes. En los árboles con raíces tablares la medida se tomó por encima de estas. Se estimó la altura de cada individuo y se identificó hasta el mínimo nivel taxonómico reconocido en campo. Además, se consignaron los datos de hábito de crecimiento, presencia, color y olor de exudado, olor de la corteza y las hojas y color de la corteza, entre otros, lo que permitió el uso posterior de las claves taxonómicas. Se recolectó una muestra de herbario únicamente de las plantas que no fue posible identificar. Las muestras se prensaron y se conservaron en alcohol al 70 % hasta que fueron llevadas al herbario CUVC de la Universidad del Valle, donde fueron procesadas e identificadas con ayuda de la colección y de claves taxonómicas como la de Gentry (1996). Las muestras fértiles fueron debidamente ingresadas en la colección.
Se estableció la fisionomía de cada localidad calculando los valores de riqueza, abundancia, frecuencia, densidad, área basal e índice de valor de importancia (IVI) de las especies. También se analizó su estructura o patrón espacial de distribución con clases diamétricas (DAP) y altura, utilizando las formulas descritas por Rangel & Velázquez (1997).
Análisis de datos. Se utilizaron los valores observados totales (especies y morfoespecies) y los estimadores de riqueza Chao 1, Jacknife 1 y Bootstrap para calcular la efectividad del muestreo con el programa EstimateS 9.1.0 (Colwell, 2013). Se construyó una curva de rarefacción en el programa PAST (Hammer, et al., 2001) para corregir las diferencias en la abundancia de plantas entre las localidades y comparar la riqueza teniendo en cuenta la localidad con menor número de individuos registrados (Gotelli & Graves, 1996; Chao, et al., 2016;Ibañez, et al., 2016). Se evaluó la diversidad alfa de cada localidad como el promedio de la riqueza de especies por transecto, y la diversidad beta como el promedio de todas las posibles comparaciones entre parejas del índice de Sørensen en los diez transectos y, por último, la diversidad gamma como el número total de especies por localidad. Estos parámetros se compararon entre localidades utilizando una prueba Anova de una vía y, posteriormente, una prueba de Tukey para determinar las localidades entre las cuales existían diferencias empleando el programa Statistica 7.0 (StatSoft, 2004).
Relación de la vegetación con los parámetros climáticos. En este análisis se utilizaron las cinco localidades muestreadas y las ocho registradas por Phillips & Miller (2002), así como las variables climáticas de precipitación media anual, variabilidad de la precipitación, temperatura media anual y variabilidad de la temperatura, tomadas de la base del WorldClim (Hijmans, et al., 2005) con resolución de 5 minutos. Se hicieron correlaciones lineales de Spearman entre la riqueza de especies y las variables climáticas para establecer la relación entre estos factores utilizando el programa Statistica 7.0 (StatSoft, 2004). Asimismo, se realizaron análisis de regresión para determinar la relación entre la riqueza de especies y las variables climáticas.
Resultados
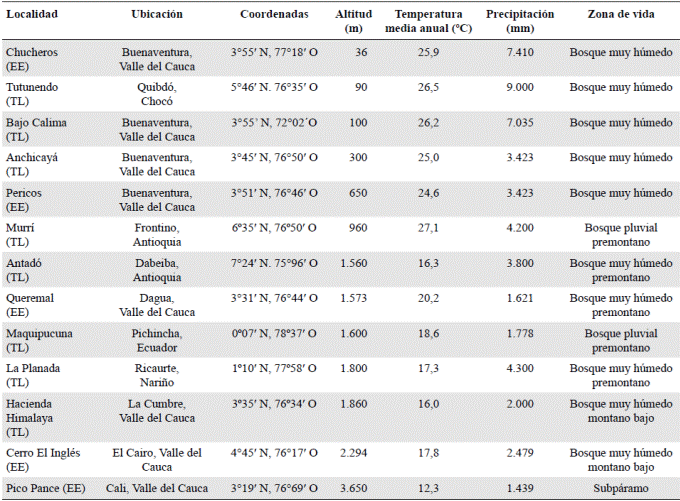
Caracterización fisionómica. La localidad Playa Chucheros presentó la mayor riqueza y abundancia, con 472 individuos de 201 especies (anexo 1), pertenecientes a 34 familias; a nivel de familia no se pudieron identificar 49 individuos de 37 morfoespecies diferentes. Por otra parte, en Queremal y Cerro El Inglés se registraron una riqueza y una abundancia similares, con 329 individuos de 98 especies y 31 familias en Queremal, y 371 individuos de 99 especies y 33 familias en Cerro El Inglés. En Queremal no se pudieron identificar 24 individuos de 20 morfoespecies y en Cerro El Inglés 12 individuos de nueve morfoespecies. La riqueza y la abundancia de las familias y las especies dominantes variaron entre las localidades (Tabla 2), aunque siempre predominaron Rubiaceae, Melastomataceae y Arecaceae, tanto con número de especies como de individuos. Myrtaceae y Sapotaceae solo se registraron en Playa Chucheros; Araceae fue la única familia que no se registró en Chucheros y Malvaceae en Queremal. Cyathea caracasana (Klotzsch) Domin (Cyatheaceae) y Elaeagia utilis (Goudot) Wedd. (Rubiaceae) se registraron tanto en Queremal como en Cerro El Inglés. En la Tabla 1S, https://www.raccefyn.co/index.php/raccefyn/article/downloadSuppFile/626/3026 se presentan las especies encontradas en todo el gradiente.
Tabla 2 Familias y especies vegetales con mayor índice de valor de importancia (IVI) en Playa Chucheros, Pericos, Queremal, Cerro El Inglés y Pico Pance, Valle del Cauca, Colombia
La mayoría de los individuos correspondió a árboles o arbustos, y solo se registraron diez lianas en Chucheros (2,1 %), ocho en Queremal (2,4 %) y cuatro en Cerro El Inglés (1 %). En Cerro El Inglés se registró el mayor valor de área basal, con 8,90 m2, seguido de Playa Chucheros con 5,98 m2, Pericos con 5,03 m2, Queremal con 3,70 m2 y Alto Pance con 0,929 m2. En cuanto a la densidad, frecuencia y cobertura, la familia Arecaceae presentó el mayor valor de importancia (IVI) en cuatro localidades (PlayaChucheros=19,13 %, Pericos=52,5 %, Queremal=25,71 % y Cerro El Inglés=15,12 %). Además, diferentes especies de esta familia se destacaron por registrar el mayor IVI (Playa Chucheros: Attalea sp., una con 15,14 %; Pericos: Wettinia quinaria con 9,2 %; Queremal: Wettinia sp., una con 16,34 %, y Cerro El Inglés: Socratea sp. 1 con 13,11 %). Las familias Rubiaceae y Melastomataceae también se registraron como una de las cinco más importantes en las cuatro localidades, con variaciones en cuanto a su importancia y representatividad (Tabla 2). Por otro lado, en Pico Pance las familias dominantes fueron Asteraceae (IVI=39,71), Ericaceae (IVI=26,11) y Melastomataceae (IVI=20,23).
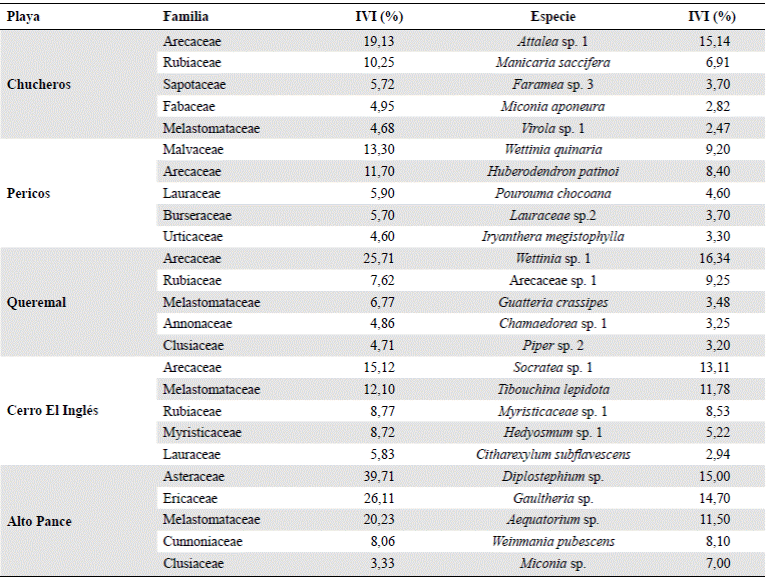
Según sus clases diamétricas, en las cinco localidades muestreadas se presentó un patrón similar, con una distribución en forma de “J” invertida (Figura 1 A), y en todos los casos la clase I contenía más de la mitad de los individuos (entre 68 y 82 %), presentando luego una drástica disminución, y la clase II entre 12 y 17 %; los valores continuaron descendiendo de modo que ninguna de las otras clases presentó más del 10 % de los individuos.
Figura 1 Distribución de los individuos de la vegetación en Playa Chucheros, Queremal, Pericos, Pico Pance y Cerro El Inglés, Valle del Cauca, Colombia. a) Clases de diamétricas. b) Clases de altura
Según las clases de altura (Figura 1 B), en Playa Chucheros y Pico Pance se presentó una distribución de tipo Poisson, en la cual la clase II presentó más individuos que la clase I, con un decrecimiento posterior en las clases de la III a la VII. En Pericos y Cerro El Inglés la clase I incluyó más individuos, aunque los valores fueron similares a los de la clase II. Luego, las clases III y IV presentaron de nuevo valores similares para luego disminuir entre las clases V a VII. En Cerro El Inglés se dio un aumento en la última clase, en tanto que en Queremal se presentó una distribución del tipo de “J” invertida, muy similar a la de las clases diamétricas.
Riqueza de especies de árboles y parámetros fisionómicos. Ninguna de las cinco curvas de acumulación de especies se comportó asintóticamente. Los porcentajes de eficiencia del muestreo menores de 100 % (Tabla 3) evidenciaron que en las cuatro localidades aún hay especies por registrar. Según la curva de rarefacción (Figura 2), la riqueza fue considerablemente mayor en Playa Chucheros y Pericos, muy similar en las localidades de Queremal y Cerro El Inglés, y más baja en Pico Pance.
Tabla 3 Estimadores de riqueza y eficiencia de muestreo de la vegetación en Playa Chucheros, Pericos, Queremal, Cerro El Inglés y Pico Pance, Valle del Cauca, Colombia
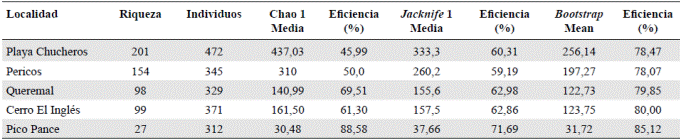
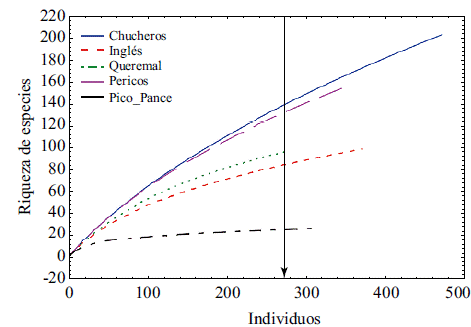
Figura 2 Curvas de rarefacción basadas en individuos de la vegetación registrados en Playa Chucheros, Pericos, Queremal, Cerro El Inglés y Pico Pance, Valle del Cauca, Colombia
Los estimadores de la riqueza de especies indicaron que en Queremal y Cerro El Inglés la riqueza fue parecida, en tanto que en Playa Chucheros estos valores se duplicaron según los estimadores Chao 1 y Jacknife 1, y se triplicaron con la herramienta Bootstrap (Tabla 3). La diversidad alfa en Playa Chucheros (30 especies) fue similar a la de Pericos (25 especies), pero significativamente mayor (Anova: p<0,05) que en Queremal (cerca de 16 especies), en Cerro El Inglés (cerca de 18 especies) y en Pico Pance (cerca de siete especies) (Tukey: p<0,05) (Figura 3). La diversidad beta no presentó diferencias significativas entre las cinco localidades (Anova: p=0,442).
Figura 3 Parámetros fisionómicos de la vegetación de Playa Chucheros, Pericos, Queremal, Cerro El Inglés y Pico Pance, Valle del Cauca, Colombia. a) Altura. b) Circunferencia a la altura del pecho (CAP). c) Diversidad alfa. d) Densidad de individuos. (Promedio para los diez transectos)
La densidad de individuos fue estadísticamente mayor (Anova: p<0,05) en la localidad de menor elevación (Chucheros), en tanto que los otros sitios presentaron una abundancia similar (Figura 3). El CAP fue mayor en El Cerro El Inglés, y más bajo en Alto Pance. Por el otro lado, la altura promedio de los árboles fue menor en Alto Pance, seguido de Queremal, en tanto que en las otras localidades las alturas fueron similares.
Relación de la riqueza de especies con los parámetros climáticos. El análisis de la riqueza en función de la altitud en las cinco localidades evaluadas en el estudio y en las otras reportadas en la literatura arrojó mayores valores de riqueza en las zonas bajas, con un pico en Bajo Calima, a los 100 m. La diversidad disminuyó drásticamente hasta Pico Pance, a 3.650 m (Figura 4). La temperatura y la precipitación media anuales, así como la altitud, se correlacionaron significativamente con la riqueza de la vegetación, pero no así la estacionalidad de la precipitación y la temperatura (Tabla 4). La variable que más se relacionó con la riqueza fue la altitud, seguida de la temperatura media y, por último, la precipitación. En Bajo Calima se registró la mayor riqueza (261 especies) y el menor valor de precipitación medio (7.035 mm), comparado con Playa Chucheros (201 especies y 7.410 mm). La riqueza se relacionó de forma lineal con la temperatura (R2=0,72, p<0,05) (Figura 4B), y de forma logarítmica con la precipitación (R2=0,86, p<0,05) (Figura 4C).
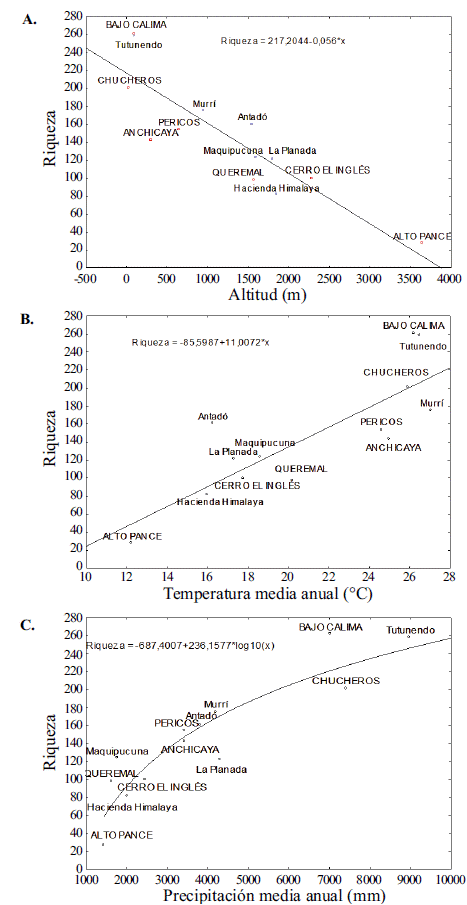
Figura 4 Riqueza de especies arbóreas en función de la altitud (A), temperatura (B) y precipitación (C) media anual en el Chocó biogeográfico de Colombia y Ecuador. La línea indica la tendencia de la relación en los análisis de regresión lineal y no lineal. Los cuadrados rojos corresponden a las localidades de este estudio y los círculos azules a las reportadas en la literatura.
Tabla 4 Correlaciones de Pearson entre la riqueza de especies y la altitud con las variables climáticas en 12 localidades entre los 36,8 y los 3.650 m de altitud en el Chocó biogeográfico de Colombia y Ecuador. * Correlaciones significativas. DA: densidad de árboles; AB: área basal; DE: desviación estándar
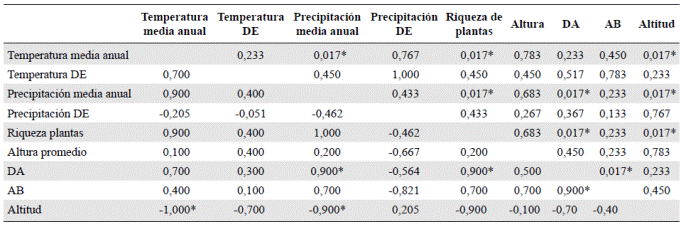
La temperatura presentó una correlación negativa perfecta con la elevación (Tabla 4), y la precipitación presentó una correlación menor. Por otra parte, la variación de la precipitación y de la temperatura no se correlacionó de manera significativa con la elevación. La riqueza de plantas presentó una correlación positiva con la elevación, la temperatura, la precipitación y la densidad de árboles, en tanto que esta se correlacionó positivamente con la precipitación y el área basal.
Discusión
La familia Rubiaceae fue la de mayor riqueza de especies y gran abundancia de individuos. Sin embargo, en las cuatro localidades ubicadas por debajo de 3.000 m el mayor IVI se registró en la familia Arecaceae por su gran abundancia de individuos y la extensión del área basal. La familia Melastomataceae también se sitúo entre los primeros cinco valores de IVI. Estas mismas tres familias también habían sido de gran importancia en la caracterización fisionómica de las dos localidades adicionales, Bajo Calima y Bajo Anchicayá (Phillips & Miller, 2002). Por el otro lado, en la localidad de Pico Pance las familias más importantes fueron Asteraceae y Melastomataceae, resultados que concuerdan con los hallados en el Chocó biogeográfico de Colombia, donde Rubiaceae se ha registrado como la familia más diversa, con 342 especies de 72 géneros, en tanto que Melastomataceae ocupa el tercer lugar, con 225 especies de 27 géneros, y Arecaceae, el decimocuarto lugar, con 86 especies de 30 géneros (Rangel-Ch & Rivera-Díaz, 2004; Kappelle & Brown, 2001).
Solamente dos especies se registraron tanto en Queremal como en Cerro El Inglés: Cyathea caracasana, un helecho arborescente de amplia distribución, que se encuentra generalmente en zonas elevadas por encima de los 1.370 m de altitud (Murillo-A & Murillo-P, 2003), y la especie más común del género Elaeagia de la familia Rubiaceae en Colombia, E. utilis, que crece con frecuencia entre los 1.300 y los 2.000 m (Mora-Osejo, 1977).
En general, la estructura de la vegetación de las cinco localidades estudiadas se parece a la de los bosques neotropicales (Gentry, 1982; 1988; Kappelle & Brown, 2001), caracterizados por presentar altos valores de área basal y alta densidad, debido, principalmente, a la gran cantidad de individuos con DAP bajo (60 % de los individuos de las cinco localidades con DAP entre 2,5 y 11,21 cm). En estos bosques, con el dosel generalmente cerrado, las plantas aumentan su altura mucho más rápido que su diámetro, con el fin de alcanzar la energía solar y realizar todas sus funciones fisiológicas.
Con respecto al gradiente altitudinal de riqueza, Playa Chucheros y Bajo Calima fueron las localidades con mayor riqueza y abundancia vegetal de las siete localizadas en el Chocó biogeográfico del Valle del Cauca. Estos resultados coinciden con lo encontrado por Gentry (1988) y Rangel-Ch & Rivera-Díaz (2004) en el Chocó biogeográfico de Colombia, donde observaron la mayor riqueza y variedad de flora en la llanura aluvial y las colinas bajas (0-200 m de altitud), caracterizadas por húmedad y temperatura elevadas.
La diversidad de especies vegetales observada a lo largo del gradiente altitudinal disminuyó al aumentar la altitud, encontrándose los mayores valores en Playa Chucheros, Bajo Calima y Tutunendo, en tanto que empezó a disminuir en Bajo Anchicayá hasta alcanzar las zonas de montaña alta, donde la riqueza fue la más baja, comportamiento que corresponde al patrón de diversidad de especies decreciente con la altitud. Una de las causas potenciales que determinan este patrón es la relación entre especies y área (McCain & Grytnes, 2010), pues, a la manera de una pirámide, el área disponible disminuye al aumentar la altitud a lo largo de una montaña, así, en las zonas altas hay una menor cantidad de energía solar disponible, de recursos y de ambientes diferentes que servirían como refugio de varias especies, comparadas con las zonas bajas (Brown & Lomolino, 1998). Por otra parte, Whittaker (2007) comparó los sistemas montanos con base en la teoría de la biogeografía de islas, ya que están aislados de otros sistemas, por lo cual aumentan las tasas de extinción y disminuye el recambio de especies y, en consecuencia, el equilibrio se alcanza con menores valores de riqueza. El patrón decreciente de la riqueza de plantas se mantuvo en la curva de rarefacción, como lo sugiere Ibáñez, et al. (2016), lo cual indica que esta disminuyó de manera consistente a pesar de las diferencias en el número de individuos muestreados en cada localidad.
En este estudio las variables climáticas de temperatura y precipitación presentaron una significativa correlación con la riqueza de especies (Grime, 2006; Hamilton & Perrot, 1981; Huston & DeAngelis, 1994; Sánchez & López Mata, 2005; Fischer, et al., 2011; Gómez, et al., 2016; Ibáñez, et al., 2016), lo que implica que en ambientes montanos las bajas temperaturas y la precipitación son los factores limitantes para la mayoría de las especies (Gentry, 1986). La hipótesis propuesta para explicar este patrón es la del cambio climático gradual o estabilidad climática, la cual establece que en latitudes y altitudes altas el estrés climático produce una gran presión sobre la ecofisiología de muchas especies, limitando así su riqueza (Palmer, 1994; Rahbek, 1995; Vázquez & Givnish, 1998; Brown, 2001; Vetaas & Grytnes, 2002; Storch & Gaston, 2004). De esta manera, en elevaciones altas la baja temperatura y la precipitación funcionan como filtro ambiental que reduce la riqueza de especies (Vázquez & Givnish, 1998; Gómez, et al., 2016). En este caso, Pico Pance presentó los valores más bajos de temperatura y precipitación, en tanto que en Bajo Calima, Chucheros y Tutunendo se registró la diversidad más alta, así como la mayor temperatura y precipitación.
La precipitación y la temperatura se correlacionaron fuertemente con el número de especies en el gradiente altitudinal. En una revisión de 45 localidades en 11 países, Gentry (1986, 1988) aseguró que la tendencia más predecible es la fuerte relación de la curva de precipitación y riqueza de especies vegetales, la cual alcanza una asíntota (saturación) en cerca de 250 especies con 4.000 mm de precipitación anual. Esto concuerda con lo observado en esta investigación, ya que en Playa Chucheros y Bajo Calima, con los promedios de precipitación anual más altos (7.410 y 7.035 mm, respectivamente), se presentan valores de diversidad cercanos a esta predicción. Después de este punto crítico de 4.000 mm de precipitación, es posible que otros factores entren a tener un papel más determinante en la riqueza de las especies. Se podría explicar que las 60 especies adicionales de Bajo Calima son producto de la diversidad de ambientes por área en zonas de mayor relieve, lo que genera diferentes inclinaciones, exposición a la luz, sustratos geológicos y, por lo tanto, variedad de microambientes que se utilizan como nichos de otras especies de plantas (Vázquez & Givnish, 1998; Fischer, et al., 2011), en comparación con zonas planas como Playa Chucheros. Por el otro lado, Gentry (1988) demostró que las curvas de acumulación de especies vegetales en varios sitios representativos alcanzaban la asíntota en la mitad del esfuerzo de muestreo en bosques de poca diversidad, en tanto que en sitios con gran diversidad la curva apenas se niveló, y, por lo tanto, el valor poco variable de 250 especies después de los 4.000 mm de precipitación anual podría ser un efecto de las limitaciones del método de muestreo. Debido a ello, se sugiere ampliar el área de muestreo, lo cual llevaría, probablemente, a revelar diferencias significativas en las riquezas de estos sitios. Sin embargo, hay que tener en cuenta que la diversidad de especies es producto no solo de la precipitación sino de la interacción del conjunto de factores bióticos, abióticos e históricos característicos de cada localidad.
Por ejemplo, la gran riqueza de especies en bosques húmedos del Neotrópico se ha relacionado con las interacciones con frugívoros especializados y nichos de dispersión más restringidos y mejor sintonizados, ya que se encuentra la misma cantidad de especies que utilizan la anemocoria como mecanismo de dispersión en bosques secos y húmedos, por lo que la dispersión de las especies adicionales provenientes de bosques húmedos casi siempre se da mediante la dispersión por zoocoria (Gentry, 1986; Correa-Gómez, et al., 2013), como en las familias más importantes en las zonas muestreadas: Arecaceae, Melastomataceae y Rubiaceae con frutos carnosos y de gran importancia ecológica.
La densidad de árboles se relacionó con la riqueza de plantas, el área basal y la precipitación, lo cual puede deberse a que a medida que aumenta la precipitación, aumenta la productividad primaria o disponibilidad de energía (hipótesis de la relación de energía y riqueza) (McCain & Grytnes, 2010; Wright, 1993; Wright, et al., 1993), lo cual permite un mayor número de individuos que aumenta la probabilidad de un mayor número de especies. Por otro lado, el mayor número de individuos está relacionado con una mayor área basal.
La ventaja de determinar estos patrones es que se puede predecir con cierta confiabilidad la tendencia de la riqueza de plantas, su composición e, incluso, los comportamientos fenológicos al analizar las variables de una localidad determinada. Para lograr una mayor confiabilidad en estos análisis se debe estandarizar el método de muestreo, el área muestreada y la definición de riqueza de especies.
Conclusión
En conclusión, el patrón altitudinal de las comunidades vegetales en el Chocó biogeográfico del Valle del Cauca evidenció que la riqueza de especies vegetales era mayor en zonas altitudinales bajas y disminuía al aumentar la altitud. En Bajo Calima se registró un punto máximo de diversidad, con 261 especies de plantas vasculares en 0,1 hectáreas a 100 m de altitud. Además, la temperatura fue el factor más estrechamente asociado con la riqueza de especies. Se recomienda repetir este tipo de estudios en el tiempo y en la misma localidad para poder comprender cómo cambian los ecosistemas y cómo se están viendo afectados por el hombre y por las rápidas variaciones climáticas, como el aumento en la temperatura media global que se predice para este siglo. Este tipo de estudios ayudará a comprender fenómenos como la tasa acelerada de extinción, la migración de especies hacia zonas altitudinales altas, el cambio en la composición de comunidades y en su fisiología y fenología. También sería importante emprender esfuerzos de muestreo en diferentes épocas del año para aumentar la resolución de la caracterización florística de las zonas y, eventualmente, encontrar especies aún no descritas, con el propósito de plantear, ordenar, sistematizar y fortalecer planes de manejo de estas importantes áreas.