











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
El género Symplocos se compone de unas 500 especies distribuidas principalmente en áreas tropicales del mundo (Souza & Lorenzi, 2005). Las plantas de este género son importantes por su contenido de metabolitos secundarios, entre los cuales se han identificado flavonoides, lignanos, fenoles, esteroides, alcaloides, iridoides, terpenoides y antraquinonas (Badoni, et al., 2010). Estos compuestos son de interés por su actividad biológica antioxidante, antimicrobiana, antitumoral y antigenotóxica, entre otras. Por ello muchas especies del género tienen potencial para aprovechamiento en química y medicina (Badoni, et al., 2010; Patel, et al., 2012; Farooq, et al, 2017; Kar & Kumar, 2017). En China se ha reportado el uso de Symplocos paniculata como planta ornamental para paisajes urbanos, así como la utilización del aceite de los frutos para la producción de biodiesel (Liu, et al., 2017).
En Colombia se registran ocho especies de Symplocos (Bernal, et al., 2011), entre las que se encuentra Symplocos theiformis, especie nativa que se distribuye en los bosques húmedos andinos entre los 2.450 y los 3.724 m de los departamentos de Antioquia, Boyacá, Cundinamarca y Santander (Kelly, 2018). Concretamente en los bosques altoandinos del noroccidente medio de Antioquia, se ha reportado como una especie más o menos abundante, que crece al interior de robledales y bosques secundarios (Toro & Vanegas, 2002). Se le llama "té de Bogotá" debido a sus cualidades astringentes y tónicas, similares a las del té y el café (Pérez, 1996).
En estudios de propagación por vía sexual y asexual en S. theiformis se ha encontrado que hasta un 96 % de las semillas se ve afectado principalmente por insectos himenópteros de la familia Eupelmidae (Córdoba, et al., 2010) y se ha confirmado que el fruto posee un endocarpio lignificado en donde reposan de 3 a 4 semillas (Cabezas, 2016). En cuanto a la viabilidad, el 74 % de las semillas solo tiene un embrión viable, el 14 %, dos embriones viables y el otro 12 %, tres embriones viables (Córdoba, et al., 2010). Cabezas (2016) determinó una viabilidad del 36,4 % mediante la prueba de tetrazolio. Se ha registrado un porcentaje de germinación de la especie de 1,7 % con la aplicación de concentraciones bajas de AG3 (200 ppm) e iluminación continua bajo condiciones de invernadero, usando sustrato compuesto por tierra en un 40 %, turba en un 40 % y cascarilla en un 20 %, lo cual fue corroborado mediante un modelo logístico en el que el tratamiento con la mejor respuesta de germinación incluyó la presencia de luz y el uso de ácido giberélico (AG3), con una probabilidad de ocurrencia de 0,045 (Cabezas, 2016). En cuanto a la siembra de semillas bajo condiciones in vitro en medio Murashige & Skoog (MS) con AG3 (1,5 mg/l), después de dos meses de evaluación se presentó un 5 % de germinación y el enraizamiento se logró en el medio MS con 1,5 mg/L de ácido indolbutírico (AIB) y 1 mg/L de 6-N-bencilaminopurina (BAP) (Linero, 2006). Los resultados tampoco han sido buenos con la propagación asexual por esquejes (Cabezas, 2016), aunque Córdoba (2007) menciona que la especie se puede propagar vegetativamente por acodo aéreo y terrestre. Estos antecedentes evidencian que S. theiformis es una especie con requerimientos de propagación específicos, tanto por vía sexual como asexual, y que no se sabe nada sobre la tolerancia a la desecación de sus semillas.
Además, S. theiformis es una especie de distribución restringida en ecosistemas altoandinos y, dada la presión actual que soporta dicho ecosistema, es urgente estudiar su conservación y propagación y contar con la información requerida para una posible alimentación de las poblaciones in situ y su conservación ex situ.
En el presente trabajo se propuso evaluar la tolerancia a la desecación y la dormancia fisiológica en semillas de S. theiformis mediante la aplicación de los siguientes cuatro tratamientos a semillas escarificadas: T1, contenido de humedad inicial (CHi); T2, contenido de humedad reducido a 10 %; T3, contenido de humedad reducido al 5 %, y T4, AG3 en 400 ppm, bajo condiciones de invernadero.
Materiales y métodos
Los frutos de S. theiformis se recolectaron a 3.257m en el páramo El Romeral (N4° 27' 52,8" - 074° 13' 9,13'), ubicado en el municipio de Sibaté, Cundinamarca, Colombia. El espécimen recolectado se ingresó al Herbario del Jardín Botánico "José Celestino Mutis" de Bogotá (JBB22208). Dada la dificultad de encontrar poblaciones naturales de esta especie, se hizo un seguimiento fenológico a dos individuos en un pequeño parche de bosque con frutos sanos (sin afectaciones fitopatógenas) hasta el momento en que se pudieron recolectar manualmente (desgranado) los frutos en el punto de madurez ideal, es decir, cuando pasan de color verde a morado y a negro (Figura 1a).

Figura 1 A) Escala de madurez de los frutos de S. theiformis. B) Izquierda, semilla sin escarificar; derecha, semilla escarificada (retiro de parte del endocarpio lignificado) de S. theiformis
En el laboratorio los frutos se lavaron con agua y se separó manualmente el mesocarpio de las semillas, las cuales se extendieron sobre papel periódico a temperatura ambiente y a la sombra por 48 horas. Para determinar el contenido de humedad inicial (T1:CHi), se pesaron tres grupos de ± 0,5 g (6 semillas), se colocaron en crisoles y estos se llevaron al horno (Binder) a 103 °C durante 17 horas, se dejaron enfriar y se pesaron para determinar el porcentaje de humedad de cada lote (International Seed Testing Association, ISTA, 2017). Para disminuir el contenido de humedad al 5 % (T2:CH5%) y al 10 % (T3:CH10 %), se seleccionaron las semillas, se pesaron y, posteriormente, se pusieron en bolsas de tela dentro de un frasco de vidrio con una relación semilla: sílica de 1:2, bajando su contenido de humedad al 5 y al 10 % de forma gradual. Con el fin de determinar el peso requerido para alcanzar el contenido de humedad se aplicó la fórmula PFS= PIS * [(100- CHi)/ (100 CHo)], donde PFS corresponde al peso final de las semillas, PIS a su peso inicial, Chi al contenido de humedad inicial, y CHo, al contenido de humedad objetivo (Rao, et al., 2007).
Posteriormente, se hizo un corte en el endocarpio ligni-ficado (Figura 1b), en la parte apical del fruto (escarificación manual); después, las semillas T1, T2, T3 se expusieron durante 22 horas a la hidratación en agua y un último lote a hidratación con 400 ppm de AG3 (T4) A continuación, se llevó a cabo la siembra colocando las semillas de manera vertical y con el corte hacia arriba; se utilizó un sustrato a base de sustracoco y turba rubia Klasmann® (70:30), y se mantuvieron bajo condiciones de invernadero expuestas a una temperatura promedio de 7 °C durante la noche y de 33 °C en el día. Se aplicó un riego moderado cada dos a tres días, o cada ocho días según las condiciones ambientales, durante aproximadamente 10 meses; cada ocho días, se registró la germinación, considerada como la emergencia de plántulas, y también se verificó cada 8 días el crecimiento de 30 plántulas.
Con los datos obtenidos se calculó el porcentaje de germinación (PG) con la fórmula PG = (N/NS) x 100, donde N es el número de semillas germinadas y NS el número total de semillas (William, 1991). El tiempo medio de germinación (TMG), se calculó mediante la fórmula TMG = Σi=1 ni x ti / Σi=1 ni, donde ti es el tiempo en días transcurrido para la germinación en el i-ésimo día; ni es el número de semillas germinadas en el i-ésimo día y k, la última germinación (Tompsett & Pritchard, 1998). La tasa de crecimiento relativo según la altura (TCRA) se calculó con la fórmula Mt - M (t-1) / t, donde Mt es la altura final; M (t-1) la altura inicial; t el tiempo (días) transcurrido entre las mediciones (Hastwell & Faccelli, 2003), en tanto que el porcentaje de supervivencia (PS) se estableció mediante la fórmula PS= (NT/NE) x 100, donde NT es el número de plántulas trasplantadas y NE, el número de plántulas establecidas.
Análisis estadístico. Dado el poco material con el que se contaba, se empleó un diseño completamente al azar. Se sembraron tres réplicas de 20 semillas por tratamiento para un total de 240; se evaluó la normalidad mediante la prueba de Shapiro-Wilks (modificado) (p≤0,05) y, para determinar las diferencias entre los tratamientos, se hizo un análisis de varianza (ANOVA), utilizando el programa estadístico INFOSTAT, versión 2014 (Di Rienzo, et al., 2008).
Resultados
Contenido de humedad, germinación y tiempo medio de germinación. El CHi de las semillas fue de 16,55 %. Bajo condiciones de invernadero, se observó una germinación lenta y asincrónica en el tiempo con todos los tratamientos, la cual se inició a los 97 días de la siembra y se estabilizó en el día 266, sin que se observara una nueva germinación con ninguno de los tratamientos (Figura 2).
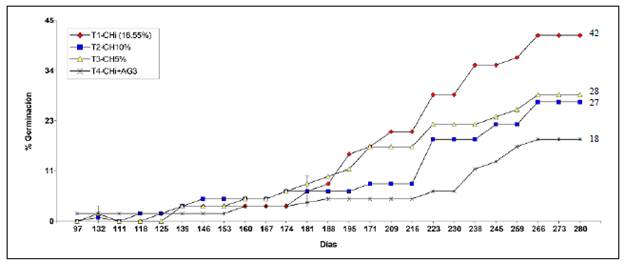
Figura 2 Germinación en el tiempo de S. theiformis con los tratamientos evaluados bajo condiciones de invernadero (n=20)
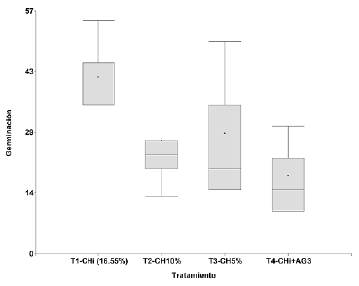
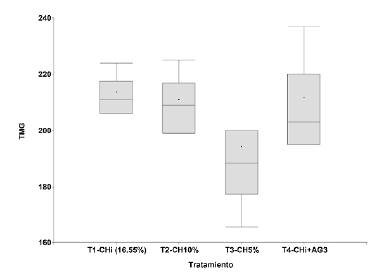
El análisis de varianza no mostró diferencias estadísticamente significativas entre los tratamientos para el porcentaje de germinación (p=0,2309) ni para el tiempo medio de germinación (p=0,4517) (Figuras 3 y 4); sin embargo, cabe destacar que las semillas frescas con el T1 y un CHi de 16,55 % fueron las que expresaron el mejor potencial germinativo (41,7 %); se observó un menor porcentaje de germinación con los tratamientos T2=CH10 % y T3=CH5 %, (26,7 y 28,3 %, respectivamente). El uso de 400 ppm de ácido giberélico en el T4 resultó inadecuado para inducir la germinación de esta especie, ya que con él se cuantificó el menor porcentaje de germinación (18,3 %). El tiempo medio de germinación en días con los tratamientos fue el siguiente: T1, 213,8; T2, 210,6; T3, 194,1, y T4, 211,5, con un rango de 194 a 214 días, lo que confirma que presentan una germinación lenta y asincrónica en el tiempo (Figura 4).
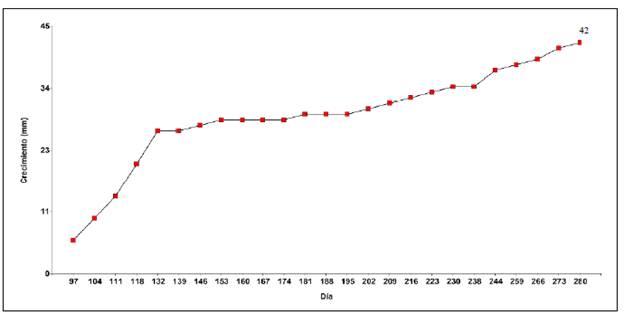
Crecimiento. La curva de crecimiento en el tiempo de la fase de plántula inicial de S. theiformis permite catalogarla como una especie arbórea de lento crecimiento, pues apenas alcanzó los 42 mm o 4,2 cm a los 280 de la siembra o a los 183 días de la germinación, lo que resultó en una tasa de crecimiento relativo según la altura (TCRA) de 0,12 mm/ día (Figura 5 y 6). En el seguimiento posterior al trasplante de las plántulas hasta los 363 días de vida, se observó un porcentaje de supervivencia de 92,1 %, momento en el que las plántulas tenían, en promedio, entre 9 y 10 cm de altura.

Discusión
Germinación y tiempo medio de germinación. La germinación fue lenta y las primeras semillas germinadas se observaron a los 97 días de la siembra. Esta respuesta contrasta con lo reportado por Córdoba (2007) y Cabezas (2016), quienes registraron el inicio de la germinación entre los 15 y los 60 días de la siembra y los 30 y los 35 días, respectivamente; las diferencias encontradas pueden explicarse principalmente por las características genéticas de las especies, las cuales están relacionadas con el origen del material vegetal que provenía de diferentes fuentes de semillas, lotes e, incluso, individuos (Doria, 2010), así como por no tener en cuenta el estado de madurez del fruto ni la madurez fisiológica de las semillas usadas, pues se llegó a sembrar, incluso, una mezcla de semillas, lo que afectaría la respuesta germinativa. En este sentido, se conoce que en algunas especies el momento de máximo desarrollo de las semientes coincide fenológicamente con la madurez de consumo de los frutos, en tanto que otras lo hacen diacrónicamente (Bewley, et al., 2013), por lo tanto, seleccionar los lotes de frutos maduros homogéneos es importante para los estudios de germinación de las semillas.
Los frutos escarificados manualmente de S. theiformis bajo condiciones controladas presentaron un porcentaje de germinación de 18,3 a 41,7 %, lo que difiere de lo encontrado por Córdoba (2007) y Cabezas (2016), quienes reportaron un máximo de germinación en esta especie de 16 y 1,7 %, respectivamente, y asociaron esta respuesta con el bajo número de semillas con potencial de germinación encontrado en las pruebas de viabilidad, así como al hecho de que la fuente de semillas presentaba afectaciones por insectos, o a que el tiempo y las condiciones del ensayo (el uso de otros sustratos a base de tierra y cascarilla y la no escarificación manual del fruto), no permitieron establecer su máximo potencial germinativo.
Los resultados obtenidos en este estudio se asemejan a lo reportado por Ribeiro (2014) en Symplocos uniflora, cuya mayor germinación se dio con semillas provenientes de frutos maduros, aunque sin escarificar (30 %), almacenadas a temperatura ambiente y expuestas a la luz (18 %) y germinadas a una temperatura controlada de 25 °C (23 %).
Dormancia. Es probable que algún tipo de dormancia pueda estar afectando la germinación en S. theiformis, ya que en otras especies como S. uniflora se ha reportado la dormancia exógena asociada con el hecho de que el fruto tiene un endocarpio lignificado que impide o dificulta la entrada de agua a las semillas (Carvalho, 2009), estructura que también está presente en los frutos de S. theiformis (Cabezas, 2016). Pareciera, entonces, que la escarificación mecánica o el retiro de una parte del endocarpio en la parte apical del fruto, como se hizo en el presente estudio, puede constituir un pretratamiento efectivo para activar el proceso de germinación, lo que demuestra que hay una dormancia mecánica y descartaría la dormancia fisiológica.
Por otra parte, no hubo una respuesta positiva al uso del AG3 con 400 ppm; por el contrario, con este tratamiento se registró el menor porcentaje de germinación (18,3 %), lo que difiere de lo hallado por Cabezas (2016), en cuyo estudio el mayor porcentaje de germinación se registró con el uso de 200 ppm de AG3 (1,7 %), por lo que es probable que no se presente la dormancia fisiológica al usar frutos maduros recién recolectados que contengan semillas frescas, o que las concentraciones sean muy bajas para generar un efecto sobre la posible dormancia de las semillas, o que, por el contrario, las concentraciones de AG3 empleadas hayan generado un efecto inhibitorio sobre la germinación, como se ha reportado en estudios de otras especies (Rojas, et al., 2001; Bradford & Nonogaki, 2007; Suárez, et al., 2011). Lo anterior puede deberse a una baja viabilidad de las semillas (34 %), o a la presencia de, por lo menos, un embrión viable (74 %), como lo han reportado Córdoba (2007) y Cabezas (2016), respectivamente, condición que podría ser determinante para la germinación y la emergencia final de las plántulas.
Tolerancia a la desecación. Al bajar el contenido de humedad de las semillas de S. theiformis de 10 a 5 %, se observó que su viabilidad se reducía, así como, posiblemente, su longevidad, por lo que se puede deducir que el potencial germinativo de las semillas de la especie disminuye cuando se reduce su contenido de humedad, lo que las clasifica como recalcitrantes (Hong & Ellis, 2002). En este sentido, en S. uniflora, que presenta semillas tetraloculadas, igualmente la viabilidad disminuye rápidamente y son consideradas recalcitrantes al almacenamiento (Lorenzi & Matos, 2008), por lo que no responden a ambientes secos y su viabilidad puede llegar, eventualmente, a cero (Hong & Ellis, 2002). Dichos resultados se pueden explicar porque la semilla proveniente de frutos maduros es pequeña, de color café claro y levemente turgente (Cabezas, 2016), y contiene estructuras que podrían verse directamente afectadas al bajar el contenido de humedad con la consecuente pérdida de turgencia y posible desecación del embrión, lo que, en últimas, afecta su viabilidad. Sin embargo, aunque la disminución prevaleció, se observó un leve aumento de germinación con el tratamiento CH5 % (28,3 %), frente al CH10 % (26,7 %), probablemente por las diferencias en el estado de deshidratación inicial de las semillas al alcanzar su estado de maduración, ya que se conoce que la fase final de maduración de algunas está acompañada por la deshidratación celular, la cual empieza con la pérdida de agua para el suministro vascular de la planta madre a la semilla (Bewley & Black, 1994), situación que favorece claramente a las semillas tolerantes a la desecación e, incluso, mejora su viabilidad y potencial de almacenamiento (Nkang, 2002). Dicha variación en la tolerancia a la desecación puede deberse a características intrínsecas de la planta y a las condiciones ambientales, lo que resulta en que la planta madre pueda producir semillas con un menor grado de recalcitrancia (Magnitskiy & Plaza, 2007).
Crecimiento. La humedad constante, el efecto del uso de sustratos de granulometría fina con alta capacidad de retención de humedad (turba y coco) frente a sustratos elaborados principalmente con tierra y cascarilla (Cabezas, 2016), probablemente potencializó las probabilidades de germinación de las semillas; estas condiciones se asemejan al patrón de regeneración natural de S. paniculata, especie que a pesar de que su dispersión se da en la estación seca, muestra picos de germinación en temporada lluviosa (Viswanath, et al, 2002). En esta línea, es claro que, a pesar de pertenecer a un mismo género o familia, cada especie requiere de condiciones ambientales (temperatura, luz, oxígeno y humedad) específicas para lograr la germinación de sus semillas (Carvalho & Nakagawa, 2000; Corbineau, et al., 2014).
Bajo condiciones de invernadero, las plántulas de S. theiformis obtenidas de la germinación requieren de especial cuidado; debe mantenerse una humedad moderada a baja en el sustrato, hacer revisiones fitosanitarias permanentes y trasplantar a un sustrato con buena materia orgánica y más nutrientes al llegar a los 5 cm de altura, momento en el cual se observan plántulas con un tallo de grosor bien establecido y acorde con su edad y, en promedio, dos pares de hojas verdaderas, lo que, al parecer, hace que su porcentaje de supervivencia bajo condiciones controladas sea mayor del 90 % después de 363 días de vida. En consecuencia, puede inferirse que se trata de una especie de lento crecimiento, sobre todo en la fase de plántula, condición que dificultaría su establecimiento y regeneración natural bajo condiciones naturales.
Al parecer, al igual que S. uniflora, la estrategia reproductiva de S. theiformis consiste en producir muchas semillas (Lorenzi & Matos, 2008), para así aumentar la posibilidad de germinación y establecimiento de plántulas. Sin embargo, es probable que las condiciones de su ambiente natural no permiten una alta tasa de regeneración por semilla, lo que impide que sea abundante en el ecosistema. Según Zhang, et al. (2005) en Symplocos laurina la propagación es predominantemente clonal, con un embotellamiento en la propagación sexual en la etapa del paso de la semilla al almacigo y de almacigo a individuo adulto.
Conclusiones
Para que la propagación sexual de S. theiformis tenga un alto potencial germinativo, se debe realizar la siembra inmediata de semillas provenientes de frutos completamente maduros con su humedad inicial y escarificando manualmente el endocarpio, debe sembrarse de forma inmediata. Se observó que los frutos presentaban una dormancia mecánica y se descartó la dormancia fisiológica en las semillas por los resultados obtenidos con el uso de AG3. Además, se comprobó que las semillas no respondieron adecuadamente a la desecación y fueron recalcitrantes. La especie tiene una germinación lenta y asincrónica en el tiempo, así como un lento crecimiento de las plántulas. Como estrategia de conservación, se recomienda mantenerla en un banco de plántulas in vivo, en colecciones vivas, o introducirla en bancos de germoplasma in vitro.

