











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Las quebradas de cabecera son ecosistemas extremadamente heterogéneos, no solo por sus condiciones biofísicas particulares, sino por la biodiversidad que comúnmente resguardan, por lo que su comunidad de macroinvertebrados acuáticos suele presentar una gran variabilidad espacio-temporal (Gomi, et al., 2002; Gordon, et al., 2004; Clarke, et al., 2008; Bae, et al., 2016). En algunos estudios se ha evidenciado la relación entre los factores geomorfológicos e hidroclimatológicos y la distribución de las especies de organismos (Ramírez & Pringle, 1998; Malmqvist, 2002; Gordon, et al., 2004; Ríos-Pulgarín, et al., 2016). También se ha reportado la relación entre los mesohábitats (rápidos o remansos) e, incluso, la heterogeneidad de sustratos con la colonización de los macroinvertebrados acuáticos con mayor diversidad en los sustratos rocosos y de hojarasca (Tomanova, et al., 2006; Domínguez, et al., 2009; Chará, et al., 2010).
Para evaluar la estructura trófica, es decir, la manera como los macroinvertebrados acuáticos dependen del recurso alimentario, es necesario comprender las relaciones entre los procesos físicos y los biológicos (Vannote, et al., 1980). En las microcuencas de cabecera la entrada de materia orgánica gruesa en partículas suele ser la principal fuente de energía porque hay una estrecha relación con el ecosistema terrestre (Ríos-Touma, et al., 2011; Meza-Salazar, et al, 2012), por lo que en estos tramos se observa una gran diversidad de macroinvertebrados acuáticos asociada con la alta heterogeneidad de hábitats (Jacobsen, et al., 2008; Torres-Zambrano & Torres-Zambrano, 2016).
En Colombia se han estudiado las relaciones entre la disponibilidad de recursos y la estructura de los grupos funcionales alimenticios (GFA), así como la diversidad específica asociada con los hábitats (Rodríguez-Barrios, et al., 2011; Chará-Serna, et al., 2012; Vásquez-Ramos & Reinoso, 2012; Longo & Blanco, 2014; Torres-Zambrano & Torres-Zambrano, 2016; Villada-Bedoya, et al., 2017) de los ríos del trópico con intervenciones antrópicas. Sin embargo, también es necesario estudiar las quebradas de las cabeceras y los nacimientos de sistemas fluviales para comprender y ampliar el conocimiento sobre la dinámica ecológica de nuestros ríos (Ramírez & Gutiérrez-Fonseca, 2014), así como los procesos que ocurren entre los sistemas terrestres y acuáticos (Heino, et al., 2003; Clarke, et al., 2008; Lorion & Kennedy, 2009; Bae, et al., 2016).
Específicamente en la cuenca del río Otún se han hecho estudios como los de Bernal, et al. (2006) y Chará-Serna, et al. (2010) en los que se caracterizaron los macroinver-tebrados acuáticos presentes en quebradas andinas tomando como referente para el análisis los microhábitats. En la zona de estudio, Walteros, et al. (2016) evaluaron la calidad ecológica de la microcuenca Dalí con base en algunos indicadores estructurales y funcionales de interés.
Si bien hay estudios sobre el papel de los macroinvertebrados acuáticos en los ecosistemas andinos, es necesario explorarlo en las corrientes de cabecera, donde han sido evidentes algunos efectos de perturbación. Un estudio crucial para esta investigación es el propuesto por Lorion & Kennedy (2009), quienes compararon los macroinvertebrados acuáticos en corrientes de cabecera con rangos altitudinales similares donde eran evidentes los efectos de la deforestación en la franja forestal ribereña. En el caso de la microcuenca Dalí, sin embargo, actualmente se evidencia un proceso de sucesión ecológica natural.
En este contexto, el propósito del presente estudio fue evaluar el efecto de la heterogeneidad de microhábitats en la riqueza de taxones y de los grupos funcionales alimenticios en los macroinvertebrados acuáticos presentes en la microcuenca Dalí. Las preguntas de investigación fueron las siguientes: ¿cómo varía la riqueza taxonómica y de los grupos funcionales alimenticios en los diferentes tramos muestreados en la quebrada Dalí y en sus tributarios? y ¿cómo determina la heterogeneidad de microhábitats acuáticos la estructura funcional alimenticia de los macroinvertebrados en la microcuenca? Se esperaba que, en la parte media de la microcuenca, donde es evidente el proceso de sucesión ecológica natural y existe una interacción con el complejo de humedales, la riqueza de taxones y la abundancia de macroinvertebrados acuáticos según los grupos funcionales alimenticios fuera similar a la composición y la estructura trófica del tramo de referencia, reconociendo las excelentes condiciones biofísicas y la alta heterogeneidad en el tramo muestreado.
Materiales y métodos
Area de estudio
La microcuenca de la quebrada Dalí se encuentra en la vereda San José, municipio de Santa Rosa de Cabal, Risaralda, y pertenece a la cuenca alta del río Otún (Figura 1). Tiene una extensión de 430 ha, con un rango altitudinal entre los 1.750 y los 2.700 m, es decir, se ubica en la franja de bosque andino. La corriente principal de la quebrada Dalí se considera de segundo orden, en tanto que sus dos tributarios son de primer orden (Walteros, et al., 2016).
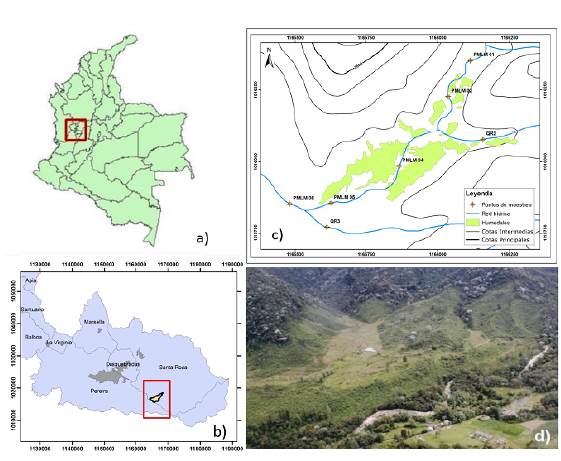
Fuente: SIG-UTP (2017) y Aguas y Aguas de Pereira (2012).
Figura 1 Microcuenca Dalí, cuenca alta del río Otún. a) Ubicación de las estaciones de muestreo en Dalí y sus tributarios. b) Referencia con el departamento de Risaralda. c) Ubicación de las estaciones de muestreo en Dalí y sus tributarios. d) Fotografía aérea de la microcuenca Dalí (parte media, zona de planicie inundable).
En su parte alta esta microcuenca presenta un relieve montañoso de pendiente mayor a 60 %, con predominio de bosque secundario natural y ripario. En la parte media, la quebrada Dalí recorre un valle en el que se encuentra un complejo de humedales del tipo de pantano arbustivo, el cual drena a esta fuente. Por último, en la parte baja de la quebrada reaparece la pendiente y hay una densa protección del bosque de ribera, lo que hace difícil su acceso.
Debe destacarse que en esta microcuenca se evidencia actualmente un proceso de sucesión natural de más de 10 años tanto en sus corrientes (principalmente la Dalí y un tributario) como en la franja intermedia de valle, donde se encuentra el complejo de humedales, proceso que se ha dado luego de las actividades ganaderas y forestales que se desarrollaron allí treinta años atrás (Parques Nacionales Naturales de Colombia-PNNC, 2007; Walteros, et al., 2016).
Muestreo
Entre el 2012 y el 2016 se monitorearon cinco estaciones en la quebrada Dalí (PMLM01, PMLM02, PMLM04, PMLM05 y PMLM06) y dos en los tributarios reconocidos por la nomenclatura QR2 y QR3 (Figura 1). Se hicieron dos muestreos por años en los picos de la temporada de lluvias y de la época seca. La estación de referencia (PMLM01, sin intervención antropogénica alguna) se seleccionó con ayuda del protocolo propuesto por Acosta, et al. (2009).
Evaluación de la calidad del hábitat
En cada estación de muestreo se definió un tramo de 20 m lineales, aproximadamente, donde se evaluó anualmente el hábitat mediante el índice de hábitat fluvial (IHF) adaptado para la región andina por Acosta, et al. (2009), el cual incluye los siguientes aspectos: inclusión de rápidos (IHF1), frecuencia de rápidos (IHF2), composición de sustrato (IHF3), régimen de velocidad y profundidad (IHF4), porcentaje de sombra (IHF5), elementos de heterogeneidad (IHF6) y cobertura vegetal acuática (IHF7).
Muestreo de los macroinvertebrados acuáticos
Para la recolección de los macroinvertebrados acuáticos se utilizó una red Surber que cubre un área de 900 cm2 por muestra tomada. Se muestrearon los microhábitats dominantes en cada corriente, es decir, sustrato rocoso (SR), orilla con vegetación (OV), hojarasca (Ho), sedimento fino (SF) y orilla con raíces (OR), con base en el protocolo propuesto por Silveira, et al. (2004). Se tomaron cuatro muestras en cada microhábitat.
El material biológico recolectado se preservó en alcohol al 90 % y se etiquetó en bolsas plásticas para su posterior análisis en el laboratorio. Se hizo la identificación taxonómica hasta el nivel de género con ayuda de claves especializadas (Manzo, 2005; Domínguez, et al., 2009; Flowers & De La Rosa, 2010; Springer, 2010; Prat, et al., 2012).
Análisis de los datos
Para conocer la riqueza y abundancia taxonómicas, se evaluaron de manera general y a escala espacial los tramos muestreados elaborando gráficos de barras. Se hizo un análisis a nivel de género para la riqueza de taxones y a nivel de órdenes para las abundancias relativas (Moreno, 2001).
Para la asignación de los grupos funcionales alimenticios (GFA) se tomaron como referencia las propuestas de Chará, et al. (2010) y Ramírez & Gutiérrez-Fonseca (2014), las cuales consideran los niveles de familia o género, y se estableció la siguiente clasificación: colectores (Co), recolectores (CoR), filtradores (Fil), depredadores (De), fragmentadores (Frag) y raspadores (Ras).
Se correlacionó la riqueza de taxones con el valor del índice de hábitat fluvial (IHF) usando el coeficiente de Pearson. Se hizo un análisis de varianza ANOVA para evaluar las diferencias entre las abundancias y la riqueza de organismos con los GFA en las estaciones y microhábitats. Las diferencias significativas (p<0,05) se sometieron posteriormente a una prueba de Tukey.
Con el fin de comparar las relaciones entre las estaciones con base en los microhábitats dominantes y las abundancias de organismos por grupos funcionales alimenticios, se hizo un análisis discriminante basado en el índice Bray-Curtis como medida de similitud. Este método de ordenación permitió visualizar las relaciones entre los grupos en un espacio multidimensional y evaluar las representaciones en dos dimensiones. Asimismo, para analizar las diferencias en la distribución de los GFA en los microhábitats dominantes, se hizo un análisis de similitud de una vía (ANOSIN) utilizando las estaciones de muestreo como factor. Para estos análisis se emplearon los programas RWizard v.1 (Guisande, 2014) y PAST 1.78 (Hammer, et al, 2001).
Resultados
Calidad del hábitat fluvial
Los tramos evaluados se caracterizaron por la dominancia del sustrato rocoso, sobretodo del tipo de cantos rodados y grava, y con la excepción del tributario denominado QR3, donde la dominancia es el sedimento fino del tipo de limo amarillo ocre. En las estaciones ubicadas en la zona media de la quebrada Dalí se registraron sedimentos finos provenientes del escurrimiento del complejo de humedales.
Aunque la diferencia altitudinal entre las estaciones no es marcada, predominó la frecuencia de rápidos en las estaciones PMLM01, PMLM02 y PMLM06, en comparación con las estaciones ubicadas en la zona media (PMLM04, PMLM05, QR2 y QR3), las cuales registran más zonas de remansos. Por lo tanto, los regímenes de velocidad y profundidad estuvieron mejor representados en las estaciones de la parte alta, encontrándose combinaciones de categorías de lentos y someros, rápidos y someros y rápidos y profundos, principalmente, en tanto que en la zona media de la microcuenca predominaron los regímenes lento y profundo y rápido y somero.
Durante el tiempo de monitoreo se observaron diferencias significativas tanto en la quebrada Dalí como en sus dos tributarios. En particular las estaciones PMLM04, PMLM05 y PMLM6 en la quebrada Dalí presentaron cambios significativos con respecto al porcentaje de cobertura del cauce y los elementos de heterogeneidad y de cobertura de la vegetación acuática (K-W: p<0,05), así como en el tributario QR3, aunque no tan pronunciados. En cuanto al porcentaje de sombra del cauce, se observaron diferencias a lo largo del tiempo de muestreo, particularmente en la zona media del cauce de la quebrada Dalí. En el 2012 se registró un cauce totalmente expuesto y en el 2016, condiciones de sombreado con ventanas en las estaciones PMLM04 y PMLM05 y de sombreado total en la estación PMLM06, razón por la cual el aporte de elementos de heterogeneidad, como la hojarasca y los troncos y ramas, también fue aumentando. En cuanto al tributario QR3, fue evidente el incremento de la cobertura de vegetación acuática, particularmente de Myriophyllum aquaticum (pinito de agua) enraizada en el sedimento fino y Cladophora (alga filamentosa), adherida al sustrato rocoso.
Composición de los macroinvertebrados acuáticos
Se recolectaron 34.144 individuos agrupados en 25 órdenes, 76 familias y 102 géneros. De los 147 taxones reportados, 101 se registraron en el microhábitat de sustrato rocoso, 97 en orilla con vegetación, 71 en hojarasca, 58 en el sedimento fino y 38 se asociaron con el de orilla con raíces (Figura 2). Este último sustrato solo fue dominante en el tributario QR2.
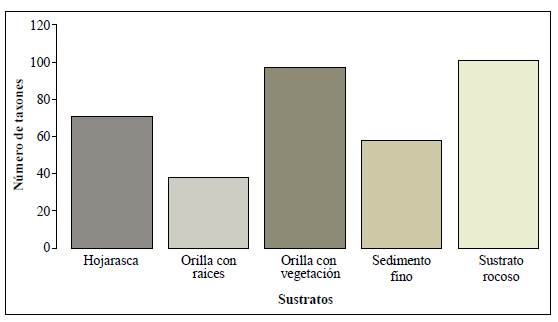
Figura 2 Riqueza taxonómica de macroinvertebrados acuáticos en cada tipo de los microhábitats en la microcuenca Dalí, cuenca alta del río Otún-Colombia
Los insectos acuáticos fueron el grupo dominante, con 58 familias distribuidas en 10 órdenes (Anexo 1,https://www.raccefyn.co/index.php/raccefyn/article/view/1085/2759). La mayor abundancia de individuos se registró en el orden Diptera, con un 26,5 % sobre el total de organismos recolectados, seguido de Ephemeroptera, con 23 %.
El microhábitat de sustrato rocoso fue el de mayormente representación, con un 45 %, seguido del de orilla con vegetación, con un 34 % y el de hojarasca, con un 11,5 %. El orden Coleoptera se destacó por su presencia en el sustrato rocoso, con 32 %, y Ephemeroptera con un 32,5 % en el de orilla con vegetación, mientras que Diptera fue más abundante en el de hojarasca, con un 46 %, y el de sedimento fino, con un 39,3 %. En el tributario QR2 el sustrato de orilla con raíces tuvo una proporción más homogénea y allí predominó la presencia de Coleoptera y Diptera (Figura 3).
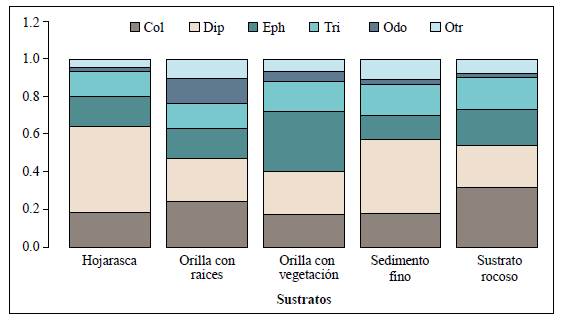
Figura 3 Composición de los órdenes más abundantes de macroinvertebrados acuáticos presentes en los diferentes microhábitats encontradod en la microcuenca Dalí, cuenca alta del río Otún-Colombia
Se encontraron diferencias significativas entre la riqueza de géneros por estaciones (p=0,007), así como de abundancias de individuos por microhábitats (p= 1,743e-08). No se registraron diferencias significativas en la riqueza de géneros por microhábitats (p=0,644).
En la prueba de Tukey se encontraron diferencias significativas, especialmente en los sustratos de hojarasca y orilla con vegetación, sustrato fino y orilla con vegetación, sustrato rocoso y hojarasca y sustrato rocoso y sedimento fino.
En general, se evidenció una correlación positiva entre la riqueza de taxones y el índice de habitat fluvial (r2=0,923).
Grupos funcionales alimenticios
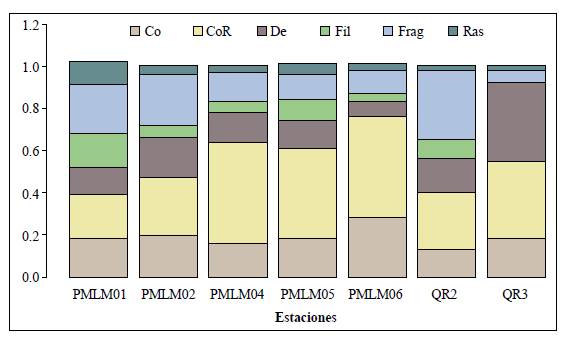
Los recolectores fueron el grupo funcional alimenticio dominante, con un 36,4 % (N: 12425, S: 28), seguido de los fragmentadores, con un 17,8 % (N: 6086, S: 16) y los colectores, con un 17,4 % (N: 5952, S: 24). Los depredadores (13,7 %, N: 4667, S: 60), filtradores (8,2 %, N: 2821, S: 9) y los raspadores (6,1 %, N: 2104, S: 16) presentaron los valores más bajos.
El porcentaje de individuos por GFA varió según las estaciones (Figura 4), encontrándose mayor representación de fragmentadores en la estación de referencia (PMLM01) y el tributario QR2, en tanto que los recolectores fueron los más abundantes en las estaciones ubicadas en la parte media de la microcuenca (PMLM04, PMLM05 y PMLM06), superando el 40 % del total de individuos. En cuanto al tributario QR3, los depredadores dominaron. Los raspadores fueron abundantes en la estación de referencia (PMLM01), y esta presentó la mayor equidad entre los grupos tróficos funcionales reportados. No se registraron filtradores ni raspadores en la estación QR3.
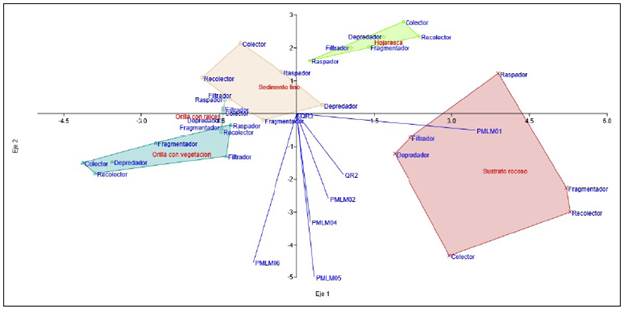
El análisis discriminante reveló que la estación PMLM01 era muy heterogénea, con dominio del microhábitat de hojarasca y abundancia de organismos en los GFA. Por lo tanto, se observó una separación fuerte y positiva entre este punto de referencia y los demás. Por el contrario, se evidenció un grado de separación con las estaciones ubicadas en la parte media de la quebrada Dalí (PMLM04, PMLM05 y PMLM06), donde, si bien se registraron organismos pertenecientes a todos los GFA, estos no fueron tan abundantes. También se evidenció en el tributario QR3 el dominio del sedimento fino y la abundancia de organismos depredadores, en tanto que el tributario QR2 fue el único con dominancia del microhábitat de orilla con raíces. En general, el análisis no reveló un patrón de asociación particular, sin embargo, es importante evidenciar que separó la estación de referencia, la PMLM01, y con un poco más de distancia, el tributario QR2, el cual presentó mejores condiciones biofísicas (Figura 5). Dicha separación se ve respaldada en el eje 1 por los dos microhábitats más dominantes en este estudio, es decir, el sustrato rocoso y el de hojarasca. El eje 1 (p: 6,14) explicó el 62 % de la varianza de abundancia de organismos en los GFA, y el eje 2 (p: 2,37), el 24 %. La correlación de esta clasificación fue del 87 %.
Discusión
Los resultados de este estudio corroboran el supuesto de que en las microcuencas de cabecera con procesos de sucesión ecológica natural como los reportados en la microcuenca Dalí, la riqueza de taxones y de grupos funcionales alimenticios es similar. Sin embargo, se observó la diferencia entre la estación de referencia y las estaciones de la parte media, donde se concentra la interacción entre el complejo de humedales y la quebrada Dalí. Puede afirmarse, por lo tanto, que los macroinvertebrados acuáticos son modelos eficientes para evaluar los impactos en corrientes alteradas (Lorion & Kennedy, 2009; Villada-Bedoya, et al., 2017).
Las óptimas condiciones del hábitat fluvial tanto en la quebrada Dalí como en el tributario QR2, son uno de los referentes claves para confirmar la heterogeneidad espacial, la cual estaría influenciada por las estaciones de la parte alta, en particular por la estación de referencia PMLM01, y puede ser determinante para la presencia de los taxones de invertebrados que habitan en estas quebradas, cuya capacidad de dispersión les permite colonizar rápidamente tramos de las corrientes después de haber presentado alguna perturbación (Gomi, et al., 2002; Malmqvist, 2002; Heino, et al., 2003; Clarke, et al., 2008).
Durante el tiempo de muestreo se evidenció que, aunque el tributario QR3 no presenta estas óptimas condiciones, sí ha pasado de tener un hábitat empobrecido a uno de mejores condiciones que facilitan el establecimiento de invertebrados acuáticos. Por lo tanto, esta corriente puede ser un buen referente para estudiar los efectos de la restauración fluvial a lo largo del tiempo.
Se registraron los patrones que determinan las características propias de las corrientes andinas, tal como lo describen Ríos-Touma, et al. (2011): la combinación de rápidos y remansos, la presencia de diferentes sustratos orgánicos e inorgánicos e, incluso, la de orillas con vegetación densa y heterogénea que incluye desde briófitos hasta árboles con buena cobertura de dosel. La presencia de estas características permite considerar a micro-cuencas como la Dalí como entornos altamente heterogéneos (Clarke, et al., 2008; Bae, et al., 2016) con una fauna única que puede influir aguas abajo (Vannote, et al., 1980).
Sin embargo, es necesario llevar el análisis hasta la escala de microhábitats para comprender las relaciones y la competencia por el recurso. En este sentido, se destacó la combinación de microhábitats de sustrato rocoso y de hojarasca en casi todas las estaciones, lo que asegura la abundancia y riqueza de organismos, así como la de los grupos funcionales alimenticios, preferentemente la de recolectores y fragmentadores, claves en la transformación de la materia orgánica que ingresa al sistema acuático (Heino, et al., 2003; Lorion & Kennedy, 2009; Ros-Touma, et al., 2009; Ramírez & Gutiérrez-Fonseca, 2014).
Se ratifica, entonces, que los grupos funcionales alimenticios están directamente asociados, no solo con el sustrato rocoso, sino también con la presencia de vegetación de ribera (Longo & Blanco, 2014), lo que fue evidente en este estudio, particularmente en los tramos donde la cobertura de dosel es plena, como en el caso de PMLM01 y QR2. Estos resultados coinciden con los de Walteros-Rodríguez & Paiba (2010) y Meza-Salazar, et al. (2012).
El predominio de órdenes como Diptera, Coleoptera y Ephemeroptera en los diferentes microhábitats también se ha observado en estudios similares como los de Walteros-Rodríguez & Paiba (2010); Chará-Serna, et al. (2012); Meza-Salazar, et al. (2012); Vásquez & Reinoso (2012); Silviera-Manzotii, et al. (2016); Walteros, et al. (2016) y Villada-Bedoya, et al. (2017). Estos órdenes suelen estar asociados a sustratos rocosos en zonas de rápidos, donde se crean ambientes heterogéneos en función de la cantidad de materia orgánica gruesa retenida entre las rocas (hojarasca) y la gran riqueza de flora perifítica (Vásquez & Reinoso, 2012), lo que incide positivamente en la disponibilidad de alimento y refugio para los macroinvertebrados acuáticos allí presentes.
Durante los cinco años de monitoreo en la microcuenca se pudo evidenciar que dominan los organismos recolectores, seguidos de los fragmentadores, tal como lo describen los trabajos previos de Bernal, et al. (2006), Chará, et al. (2010); Walteros, et al. (2016) y Villada-Bedoya, et al. (2017), quienes han realizado estudios en la zona central de los Andes. Resultados similares han sido reportados por Ríos-Touma, et al. (2009) pero en quebradas altoandinas con coberturas boscosas donde los recolectores fueron dominantes, y en un porcentaje importante también se reportaron fragmentadores.
Tal como lo evidencio el análisis discriminante, en la estación de referencia se registró, además, una gran abundancia de organismos asociada con la riqueza taxonómica y funcional según los GFA. Se considera que en este tramo se presenta el patrón heterogéneo que determina la comunidad de los macroinvertebrados acuáticos y su capacidad de colonizar otras zonas aguas abajo, lo que coincide con el concepto de río continuo (Vannote, et al., 1980). Esto, y el predominio de fragmentadores y colectores, refleja la importancia de dichos organismos en la descomposición de la materia orgánica gruesa y fina, recurso fundamental en las quebradas de cabecera, que suelen tener una densa cobertura boscosa y deberían denominarse ecosistemas heterotróficos (Chará, et al., 2010).
En cuanto a la estructura trófica de las estaciones ubicadas en la parte media (PMLM04, PMLM05 y PMLM06), se sugiere analizarla con precaución, pues durante estos cinco años se han evidenciado cambios en los procesos de sucesión de las coberturas vegetales, lo que probablemente esté condicionando la dominancia de los recolectores. Así, es probable que exista mayor disponibilidad de materia orgánica en partículas finas (MOPF) proveniente de las tasas de descomposición en los tramos altos, razón por la cual en estas estaciones es menor la proporción de fragmentadores (Greathouse & Pringle, 2006). Además, en la parte media de la quebrada Dalí disminuye la combinación de mesohábitats, por lo que el canal tiende a ser más profundo y rápido, con predominio de pastos e invasión de sus raíces, hábitats estos propicios para la presencia de diversos depredadores. Ramírez & Hernández-Cruz (2004) también encontraron que el ensamble de los macroinvertebrados acuáticos dependía de estas variables.
Según Chará-Serna, et al. (2015), las coberturas vegetales ribereñas pueden tener una influencia positiva o negativa en la comunidad y la estructura trófica de los macro-invertebrados acuáticos. En este estudio pareció ser positiva en las estaciones PMLM04 y PMLM05, donde con el transcurso del tiempo ha aumentado la densidad y la heterogeneidad de esta franja de protección, lo que, además de posibles cambios en el flujo hidrológico, estaría generando una menor variabilidad en la morfología del canal y un aumento de hábitats poco profundos. Pero en la estación PMLM06 esta condición parecería tener un efecto negativo, ya que en la franja protectora comienza a haber predominio de pastos de la familia Poaceae. Según Chará-Serna, et al. (2015), la presencia de este pasto conlleva una mayor incidencia de deslizamientos, inestabilidad del canal e invasión de la hierba.
Para concluir, los resultados sugieren la necesidad de ampliar los estudios, con el fin de conocer mejor los patrones y factores determinantes de la diversidad de macroinvertebrados en microcuencas de cabecera y suministrar información relevante para la planificación sistemática de la conservación y el diseño de reservas de protección de la biodiversidad ribereña (Clarke, et al., 2008).

