











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Las sabanas inundables abarcan más del 12 % del área de la cuenca del Orinoco en Colombia; se concentran en la depresión conformada por los departamentos de Arauca y Casanare (Mora-Fernández, et al., 2015) y proveen servicios ecosistémicos como el mantenimiento de la biodiversidad, el reciclaje de nutrientes, la formación de suelo, la producción primaria, la regulación hídrica y la provisión de belleza escénica (Mora-Fernández & Peñuela-Recio, 2013a; Lasso, et al., 2014).
La biodiversidad de las sabanas inundables de la Orinoquía colombiana está seriamente amenazada por el cambio de uso del suelo, la deforestación e introducción de pastos exóticos y el desarrollo de monocultivos extensivos (Ramírez, et al., 2011). Durante las últimas décadas ha crecido la demanda de tierra en el departamento de Casanare para expandir e intensificar actividades de alto impacto para la biodiversidad como la ganadería, el cultivo de arroz, el establecimiento de plantaciones de palma de aceite y la extracción de hidrocarburos, acelerando el deterioro de las sabanas inundables (Rippstein, et al., 2001; Mora-Fernández & Peñuela-Recio, 2013a; Mora-Fernández, et al., 2015). Aunque esto ha despertado el interés por documentar la flora del departamento, los registros de especies de plantas suelen ser más completos en municipios cercanos al piedemonte, donde hay más facilidades de acceso y movilidad, que en aquellos alejados, como San Luis de Palenque y Trinidad, en la cuenca baja del río Pauto (Córdoba, et al., 2011).
Los principales referentes para el estudio de la riqueza florística en el río Pauto son la flora de la cuenca del río Orinoco en Colombia (Cárdenas, et al., 2016; Sua-Tunjano & González-Giraldo, 2018), el catálogo de espermatófitos de la Orinoquía (Minorta-Cely & Rangel-Ch., 2014), la vegetación de sabana tropical de los Llanos de Colombia (Blydenstein, 1967), la flora del andén orinoqués (Castro-Lima, 2010), las plantas acuáticas de la Orinoquía (Fernández, et al., 2015; Madriñan, et al., 2017), el estado del conocimiento de las sabanas inundables en la Orinoquía colombiana (Mora-Fernández, et al., 2015), así como varios estudios sobre la composición y la estructura de los bosques ribereños en el río Yukao-Meta (Veneklaas, et al., 2005), en Puerto López-Meta (Carvajal, et al., 2007), y en la Reserva Tomo Grande-Vichada (Correa-Gómez & Stevenson, 2010). En Casanare se destacan los estudios sobre la flora del departamento (Córdoba, et al., 2011), las comunidades vegetales del bloque Cubiro (Pinzón-Pérez, et al., 2011), y las sabanas de Orocué (Romero-Duque, et al., 2018). En la cuenca baja del río Pauto algunos estudios han evaluado la salud ecosistémica (Mora-Fernández & Peñuela-Recio, 2013a) y han establecido la guía de campo de flora y fauna (Mora-Fernández & Peñuela-Recio, 2013b), así como la composición y estructura de los bosques ribereños (Cabrera-Amaya & Rivera-Díaz, 2016).
En el presente estudio se propuso analizar la flora de las sabanas y bosques de la cuenca baja del río Pauto en términos de riqueza, composición y distribución geográfica. La caracterización básica de la vegetación es el punto de partida para las decisiones sobre su conservación, uso y manejo sostenible, y para la evaluación del impacto ambiental de las actividades económicas, por tanto se requiere con urgencia en el departamento de Casanare y la Orinoquía colombiana. Asimismo, se elaboró un catálogo de las especies de plantas vasculares de la zona, con nuevos registros para la Cuenca del Orinoco de Colombia.
Materiales y métodos
Área de estudio
El estudio se realizó en los municipios de San Luis de Palenque y Trinidad (Casanare), en la cuenca baja del río Pauto, cuya área aproximada es de 2.874 km2 (Mora-Fernández, et al., 2011) (Figura 1). Según el sistema Thornwaite, la región presenta un clima tropical lluvioso unimodal de sabana, con una temperatura promedio de 26 °C, y una precipitación total anual de 2.048 mm distribuida entre la estación seca (diciembre-marzo) y la lluviosa (abril-noviembre), siendo mayo, junio y julio los meses más lluviosos (Mora-Fernández, et al., 2011). Según Goosen (1964) esta zona se subdivide en los paisajes de vegas, llanura eólica y llanura aluvial de desborde; estos paisajes fueron reinterpretados y denominados para efectos del estudio como vegas de ríos andinenses (ríos Pauto y Meta), sabanas inundables con influencia eólica, y sabanas inundables típicas (cuya inundación responde a la falta de infiltración del agua de las lluvias por la baja permeabilidad de los suelos arcillosos, y no al desbordamiento de los cursos de agua, Obs. Pers. Castro-Lima, 2012).
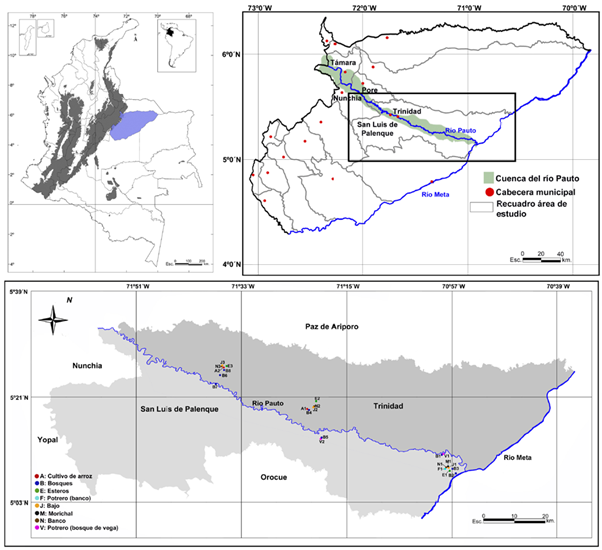
Figura 1 Ubicación de los puntos de muestreo en la cuenca baja del río Pauto (municipios de San Luis de Palenque y Trinidad, Casanare).
Las unidades de vegetación estudiadas se dividen por su fisionomía en bosques y sabanas y otros pastizales (denominadas en adelante como sabanas; ver anexo S1, figuras S1, S2 y S3https://www.racceiyn.co/index.php/raccefyn/article/view/1188/2893). Dichas unidades se definen a continuación con base en la literatura especializada (Sarmiento, et al., 1971; Sarmiento, 1990; Caro, 2006; Fernández, 2007; Baptiste & Ariza, 2008).
Sabanas. Se diferencian por su topografía, la estacionalidad en el contenido de agua del suelo y los tipos de suelos (Anexo 1, figura S1,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893) en: 1) bancos (sabanas secas o estacionales), es decir, zonas levemente elevadas de suelos arenosos bien drenados que no se inundan en la estación de lluvias y experimentan solo dos estaciones, seca y húmeda; 2) bajos (sabanas húmedas o hiperestacionales), depresiones que se inundan temporalmente por la precipitación durante la temporada lluviosa y experimentan cuatro estaciones, sequía, inicio de la saturación del suelo, inundación y disminución de la saturación del suelo; 3) esteros (sabanas semiestacionales), son depresiones de suelo arcilloso que nunca presentan déficit de agua, incluso en la estación seca, se inundan en la estación lluviosa y son habitados por plantas palustres y acuáticas; 4) banco transformado en potrero, con pastos introducidos para el mantenimiento del ganado; 5) cultivo de arroz en áreas manejadas en sabanas hiperestacionales o esteros con dominio de Oriza sativa; y 6) potreros inundables, antiguos bosques de vega transformados hoy en potreros que se inundan estacionalmente con los pulsos de inundación de los ríos andinenses.
Bosques. 1) Morichales, palmares estacionalmente inundables dominados por Mauritia flexuosa, típicos de sabanas abiertas y márgenes de cursos de agua (Anexo S1, Figura S2,https://www.racceiyn.co/index.php/raccefyn/article/view/1188/2893); 2) bosques de galería que se desarrollan en planicies aluviales y de desborde de ríos, caños y cañadas de las sabanas orinocenses (Anexo S1, Figura S2https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893); 3) bosques de vega que se extienden sobre los planos de inundación en valles aluviales de ríos andinenses (Anexo S1, Figura S3,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893), y 4) vegetación ruderal asociada con zonas abiertas alteradas cercanas a caminos y bordes de bosques, ríos y cañadas (Anexo S1, Figura S4,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893).
Fase de campo
El muestreo se hizo en el 2012 en dos salidas de campo, la primera en marzo durante la temporada seca, y la segunda en julio, en la temporada lluviosa. El detalle de la metodología empleada se presenta en el anexo S2, tabla S1, figura S5,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893.
Fase de laboratorio y análisis de información
Los ejemplares recolectados se clasificaron mediante la consulta con expertos, la revisión de la literatura especializada, y la comparación con los exsiccata de las colecciones del Herbario COL. La clasificación de las familias sigue el sistema APG IV para las angiospermas (Chase, et al., 2016) y el sistema de Smith, et al. (2006) para los pteridófitos sensu lato. Los nombres científicos, sinonimias y autores se validaron empleando la herramienta web The Taxonomic Name Resolution Service (TNRS, 2019), el Catálogo de plantas y líquenes de Colombia (Bernal, et al., 2019). Se analizó la riqueza por familias, géneros y formas de crecimiento de cada unidad de vegetación y se elaboró un catálogo sistemático de la flora vascular con todas las especies registradas para la zona tanto en parcelas de vegetación como en colecciones generales (Anexo S6,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893). Las diferencias florísticas entre las unidades de vegetación se compararon mediante un análisis de similitud basado en el índice de Serensen usando el software PAST 2.04 (Hammer, et al., 2001). Este índice también se empleó para analizar la similitud de las especies nativas de la zona de estudio según su patrón de distribución geográfica en las regiones naturales de América con base en la propuesta de Morrone (2014). La información sobre la presencia de las especies se obtuvo de la base de datos del Missouri Botanical Garden (Tropicos.org, 2019). Por último, el número de especies amenazadas en la zona se analizó con base en los libros rojos de la Flora de Colombia (Calderón, et al., 2002, 2005; Calderón, 2006; García, et al., 2006; Cárdenas & Salinas, 2007; García, 2007), los listados CITES (CITES, 2019), y la lista roja de IUCN (iucnredlist.org, 2020).
Resultados
Riqueza florística
En toda la zona estudiada se registraron 701 especies, 421 géneros y 121 familias (Tabla 1, Anexo S3, Tabla S2,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893). En cuanto a las grandes unidades fisionómicas en las sabanas, las familias más diversas fueron Leguminosae sensu lato (43), Cyperaceae (42), Poaceae (38), Malvaceae (16), Rubiaceae y Asteraceae (12 cada una), Lamiaceae (10), Euphorbiaceae (8) Onagraceae y Melastomataceae (7 cada uno), que en conjunto representaron el 60 % de la riqueza de especies (Anexo S3, Tabla S3,https://www.raccefyn.co/index.php/raccefyrVarticle/view/1188/2893). Los géneros más ricos fueron Cyperus (13 especies), Aeschynomene, Hyptis, Rhynchospora y Eleocharis (9 cada uno), Ludwigia (7) y Spermacoce (6), con el 19 % de la riqueza (Anexo S3, Tabla S4,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893). Por otra parte, las familias más ricas en especies en los bosques correspondieron a Leguminosae (49), Malvaceae y Rubiaceae (26 cada una), Poaceae (19), Asteraceae (17), Apocynaceae (16), Euphorbiaceae (15), Sapindaceae (12), Moraceae (11), Melastomataceae y Piperaceae (10 cada una), las cuales representaron el 45 % de las especies (Anexo S3, Tabla S5,https://www.raccefyn.co/index.php/racceiyn/article/view/1188/2893). Los géneros Piper, Senna y Miconia (7 cada una), Casearia, Ficus, Heliconia, Inga, Passiflora, Sida y Tillandsia (5 cada una), son los más ricos y reunieron el 12 % de las especies (Anexo S3, Tabla S6,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893).
En general, en los bosques se registró una mayor riqueza (470 especies) que en las sabanas (325), aunque al detallar las unidades de vegetación, los bosques de vega del río Pauto tuvieron más especies (231), seguidos por los bajos (141) y los bancos (131) (Tabla 1). En contraste, los esteros, morichales, cultivos de arroz y el banco transformado en potrero fueron las unidades de vegetación con menor riqueza, cada uno con valores inferiores al 10 % del total de especies (Tabla 1).
Tabla 1 Riqueza florística de las unidades de vegetación de la cuenca baja del río Pauto
| Unidad de vegetación (Abreviación) | Familias | Géneros | Especies | % de especies |
|---|---|---|---|---|
| Bosque de vega río Pauto (BV-P) | 75 | 191 | 231 | 32,9 |
| Bajo (J) | 43 | 94 | 141 | 20,1 |
| Banco (N) | 37 | 88 | 131 | 18,7 |
| Vegetación ruderal (R) | 50 | 101 | 121 | 17,2 |
| Bosque de galería (BG) | 50 | 92 | 110 | 15,7 |
| Potrero inundable (V) | 38 | 75 | 98 | 14,0 |
| Bosque de vega río Meta (BV-M) | 38 | 67 | 70 | 10,0 |
| Esteros (E) | 26 | 41 | 54 | 7,8 |
| Morichal (M) | 36 | 46 | 53 | 7,5 |
| Cultivo de arroz (A) | 19 | 40 | 46 | 6,6 |
| Banco transformado en potrero (F) | 18 | 32 | 38 | 5,4 |
| Total | 121 | 421 | 701 | 100 |
Las hierbas fueron las formas de crecimiento con mayor riqueza (42 %), seguidas de los árboles (18 %), los sufrútices (16 %) y los arbustos (15 %), en tanto que las plantas acuáticas, trepadoras leñosas y herbáceas, epífitas, hemiepífitas, palmoides y hemiparásitas representaron, en conjunto, menos del 10 % de las especies (Tabla 2). En las grandes unidades fisionómicas las formas de crecimiento predominantes en las sabanas fueron las hierbas (211 especies), seguidas por las sufrútices (81) y acuáticas (53), que sumaron el 75 % de las especies. Los bosques albergaron el 70 % de las especies, la mayoría árboles (126), seguidos por hierbas (115) y arbustos (88). Se destaca que 52 especies no habían sido registradas previamente en la Orinoquía y constituyen novedades para esta región natural (Ver Anexo S7,https://www.raccefyn.co/index.php/racceiyn/article/view/1188/2893); asimismo, se registraron dos especies por primera vez en Colombia: Nymphaea gardneriana y Sagittaria platyphylla.
Tabla 2 Riqueza de especies según forma de crecimiento en las unidades fisionómicas y de vegetación de la cuenca baja del río Pauto. BG: bosque de galería, BV-M: bosque de vega río Meta, BV-P: bosque de vega río Pauto, M: morichal, R: vegetación ruderal, J: bajo, N: banco, F: banco transformado en potrero, A: cultivo de arroz, E: estero, V: potrero inundable
| Forma de | Bosques | Sabanas | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| crecimiento | BG | BV-M | BV-P | M | R | Total | J | N | F | A | E | V | Total | |
| Acuática | 1 | 1 | 2 | 0 | 6 | 9 | 21 | 7 | 0 | 3 | 31 | 8 | 53 | 59 |
| Árbol | 50 | 24 | 52 | 22 | 11 | 126 | 2 | 1 | 0 | 2 | 1 | 1 | 7 | 129 |
| Arbusto | 27 | 12 | 46 | 7 | 11 | 88 | 6 | 9 | 0 | 2 | 3 | 9 | 27 | 105 |
| Epífita | 9 | 2 | 10 | 3 | 1 | 23 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 24 |
| Hemiepífita | 1 | 0 | 4 | 1 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 7 |
| Hemiparásita | 0 | 0 | 1 | 3 | 2 | 4 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 5 |
| Hierba | 12 | 7 | 60 | 7 | 43 | 115 | 103 | 90 | 23 | 37 | 27 | 58 | 211 | 294 |
| Palma | 0 | 2 | 4 | 1 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 7 |
| Sufrútice | 3 | 4 | 22 | 3 | 15 | 44 | 22 | 39 | 16 | 3 | 7 | 21 | 81 | 111 |
| Trepadora herbácea | 5 | 6 | 21 | 0 | 17 | 42 | 2 | 8 | 1 | 0 | 0 | 10 | 19 | 54 |
| Trepadora leñosa | 12 | 13 | 23 | 6 | 19 | 53 | 2 | 3 | 0 | 0 | 0 | 3 | 8 | 57 |
| Total | 110 | 70 | 232 | 53 | 121 | 470 | 141 | 131 | 38 | 46 | 55 | 98 | 325 | 701 |
Diferencias florísticas entre las unidades de vegetación
Con respecto a las unidades fisionómicas, en las sabanas hubo diferencias florísticas entre las inundables (bajos, arrozales y potreros) y las no inundables como los bancos (Anexo S4, figura S6-A,https://www.racceiyn.co/index.php/raccefyn/article/view/1188/2893). Los análisis de similitud de especies separaron bajos y esteros naturales de potreros y arrozales (transformados) en las unidades de vegetación inundables. Por otra parte, en las sabanas naturales hubo una clara diferencia entre esteros. No obstante, los bancos más cercanos al río Meta (N2 y N3) presentaron poca similitud florística con los demás (Anexo S4, Figura S6-Ahttps://www.racceiyn.co/index.php/raccefyn/article/view/1188/2893), que, además de encontrarse más alejados de ese río, se asemejaban más a los bajos.
Hubo una similitud menor al 50 % entre todos los bosques ribereños, lo que evidencia el bajo número de especies compartidas entre unidades de vegetación. En los bosques los análisis evidenciaron una mayor afinidad florística entre puntos más cercanos (sin importar si se trataba de bosques de galería o de vega), pues por un lado se agruparon las unidades boscosas influenciadas por el río Pauto, y por el otro, las cercanas al río Meta. Por ejemplo (Anexo S4, Figura S6-B,https://www.raccefyn.co/index.php/racceiyn/article/view/1188/2893), uno de los bosques de vega (B6) presentó menor similitud con otros bosques de vega que con dos bosques de galería (B4, B8), y todos ellos se encuentran distanciados del cauce principal del río Pauto.
Patrones de distribución de las especies
A nivel de las sabanas inundables en Colombia, 486 especies de las registradas en el presente trabajo lo están en Arauca, Casanare y Meta. De ellas, 126 se han encontrado en los tres departamentos, 361 están registradas en el departamento del Meta, 333 en Casanare, y 227 en Arauca; además, se registraron en este estudio 228 especies enumeradas previamente para el departamento del Vichada, donde predominan las sabanas no inundables. Por otro lado, ninguna de las especies encontradas es endémica de Colombia, sin embargo, Cipura gigas Celis, Goldblatt & Betancur (Iridaceae) y Diclidanthera bolivarensis Pittier (Polygalaceae) son taxones restringidos a la Orinoquía de Colombia y Venezuela (Aymard, 2017).
La mayoría de las especies es de amplia distribución en el norte de Suramérica, incluidos Brasil y las Guayanas hacia el oriente, e incluso Panamá y Costa Rica hacia el norte en algunos casos. Muchas están distribuidas en las subregiones biogeográficas Brasilera y Chaqueña y en el caso de las sabanas se distribuyen más especies en los dominios Brasilero sur (provincia Rondônia), Pacífico (provincias Sabana, Guatuso), Brasilero norte (provincias Pantepui, Guayana tierras-bajas), Mesoamericano (provincias Mosquito, Veracruz) y Chaqueño (provincia Cerrado) (Anexo S4, Figura S7-A,https://www.raccefyn.co/index.php/raccefyn/article/view/1188/2893). En los bosques hay más especies compartidas en los dominios Pacífico (provincia Sabana), Brasilero norte (provincias Pantepui, Imerí, Guayana tierras-bajas, Napo), Brasilero sur (Rondônia, Ucayali, Madeira) y Chaqueña (provincia Cerrado) (Anexo S4, Figura S7-B,https://www.raccefyn.co/index.php/raccefynarticle/view/1188/2893). Por otro lado, son muy pocas las especies que se distribuyen hasta las regiones neártica o andina (regiones templadas de América del norte y del sur, respectivamente), aunque allí hay más especies de sabanas que de bosques.
Especies amenazadas
De las 701 especies registradas en este trabajo, solo 14 (2 %) habían sido evaluadas previamente en cuanto a su estado de conservación según los libros rojos nacionales y las listas rojas internacionales. Se encontraron tres especies amenazadas: Cedrela odorata (en peligro, EN), Syagrus sancona y Tapura acreana (vulnerables, VU). Otras diez están citadas en los apéndices del CITES (CITES, 2019), diez se encuentran en el apéndice II (especies no necesariamente amenazadas cuyo comercio debe controlarse para evitar que su uso sea incompatible con la supervivencia), y una en el apéndice III (especies protegidas por lo menos en un país, y con solicitud de ayuda para controlar su comercio). Por otra parte, 15 de las especies registradas están incluidas en la lista de especies priorizadas en el marco de la Estrategia Nacional para la Conservación de Plantas en la Orinoquia (Instituto Humboldt, 2013) (Ver Anexo S5,https://www.raccefyn.co/index.php/racceiyn/article/view/1188/2893).
Discusión
Riqueza florística
Tanto la composición florística como las familias con mayor riqueza en la región del Pauto se asemejan a las registradas en otros estudios realizados en la Orinoquía que incluyeron vegetación boscosa y de sabana (Córdoba, et al., 2011; Minorta-Cely & Rangel-Ch., 2014; Cárdenas, et al., 2016; Romero-Duque, et al., 2018). La riqueza de especies observada en el presente estudio es relativamente alta comparada con otras zonas de sabana inundable en Arauca y Casanare (Córdoba, et al., 2011; Romero-Duque, et al., 2018). Por otra parte, hubo una mayor riqueza de especies en los bosques que en las sabanas, a pesar de que estas constituyen la principal cobertura vegetal de la región (Minorta-Cely & Rangel-Ch., 2014; Romero-Duque, et al., 2018). Al contrastar el número de especies registradas aquí para la cuenca baja del río Pauto con el de otros trabajos de la Orinoquía colombiana (Minorta-Cely & Rangel Ch., 2014; Cárdenas, et al., 2016; Minorta-Cely, et al., 2019), se encontró una mayor representación de las especies de bosques ribereños (50, 57 y 67 %, respectivamente) que de las especies de sábanas (38, 39 y 46 %, respectivamente).
Además de aportar al conocimiento de la flora del Casanare y de los municipios de San Luis de Palenque y Trinidad, la presente investigación pone en evidencia la necesidad de aumentar el muestreo en los bosques y sabanas de la región en donde ya se ha advertido sobre los vacíos de información florística (Córdoba, et al., 2011; Mora-Fernández, et al., 2015).
Por otra parte, las leguminosas son una de las familias más ricas en especies en los bosques ribereños del río Pauto. Esto se explica porque las plantas de esta familia obtienen nutrientes de la atmósfera y de suelos extremadamente pobres gracias a asociaciones simbióticas con microorganismos del suelo en sus raíces, lo que les permite ser exitosas en casi todos los hábitats del planeta (Steidinger, et al., 2019); a esto se suman una excepcional variación morfológica y procesos evolutivos de diversificación que, en conjunto, explican su vasta diversidad (Yahara, et al., 2013). Otros estudios realizados en la Orinoquía encontraron resultados similares (Veneklaas, et al., 2005; Caro, 2006; Miranda, 2006; Correa-Gómez & Stevenson, 2010; Castro-Lima, 2010; Pinzón-Pérez, et al., 2011; Pérez-Buitrago, et al., 2020); dichos trabajos destacan otras familias de gran riqueza en la cuenca baja del río Pauto como Rubiaceae, Melastomataceae y Moraceae que, además, presentan una gran diversidad en el piedemonte y el bosque subandino de la Orinoquía (Trujillo-C. &Henao-Cárdenas, 2017), así como leguminosas, reconocidas entre las más importantes en el Neotrópico (Gentry, 1982, 1988).
En el caso de las sabanas, la gran riqueza de familias como Leguminosas, Poaceae, Cyperaceae y Malvaceae concuerda con los estudios realizados en sabanas y ambientes húmedos e inundables de Meta, Arauca, Casanare, Vichada (Pinzón-Pérez, et al., 2011; Fernández, et al., 2015; Vera-Ospina, 2017; Romero-Duque, et al., 2018; Mijares, et al., 2019). Además, la riqueza a nivel de géneros y la composición de especies en bancos, bajos y esteros es similar a la encontrada para dichos ambientes en otras sabanas inundables en la Orinoquía colombiana y en la venezolana (Sarmiento, et al., 1971; Torres, et al., 2003; Romero-Duque, et al., 2018; Mijares, et al., 2019).
La proporción de especies según su forma de crecimiento coincide con los resultados de los trabajos analizados, pues evidentemente hay más especies herbáceas en las sabanas que en los bosques (Sarmiento, et al., 1971; Parra-O., 2006; Pinzón-Pérez, et al., 2011), donde son más abundantes las de árboles y arbustos (Sarmiento, et al., 1971; Veneklaas, et al., 2005; Caro, 2006; Parra-O., 2006; Carvajal, et al., 2007; Correa-Gómez & Stevenson, 2010).
Diferencias florísticas entre las unidades de vegetación
Tal como lo afirma Sarmiento (1990), la distribución de agua en el suelo, la microtopografía y el grado de intervención antropogénica influyen en la composición de cada unidad de vegetación; la separación entre sabanas inundables (bajos o esteros) y sabanas secas (bancos) puede explicarse por el régimen marcado de sequía que estas últimas soportan. Lo anterior se reflejó en la presencia de especies como Andropogon bicornis y Axonopus purpusii en bancos y bajos, respectivamente, y como Caperonia castenifolia y Leersia hexandra (una forrajera nativa) en bajos y esteros, concordando con trabajos previos en la Orinoquía colombiana y en la venezolana, lo que evidencia la importancia de dichas especies en sus respectivas unidades de vegetación (Tejos, 2002; Pinzón-Pérez, et al., 2011). La degradación antropogénica causa cambios evidentes en la composición de la vegetación transformada (arrozales y potreros inundables) frente a la más conservada, pero también en los bancos de la sabana inundable típica comparada con la sabana inundable sometida a la influencia eólica, más cercana al río Meta. Dado que no se inundan, los bancos de la sabana inundable típica se convierten en caminos y carreteras muy transitados en la estación lluviosa (Goosen, 1964), lo que implica cierto nivel de degradación (Mora-Fernández & Peñuela-Recio, 2013a).
Por otro lado, la composición florística de los bosques fue específica para cada sitio y tuvo, en general, una gran variación, lo que se ha observado también en otros bosques ribereños de Casanare (Romero-Duque, et al., 2018). Aunque estos ecosistemas comparten las familias más importantes, su composición y número de especies difiere en los estudios (Veneklaas, et al., 2005; Romero-Duque, et al., 2018). Las mayores diferencias en la composición de los bosques se explican por la cercanía geográfica, más no por el paisaje o el tipo de bosque. Cabrera-Amaya & Rivera-Díaz (2016) agruparon los bosques de la misma área de estudio en dos bloques por su similitud florística: paisajes de sabana inundable típica (llanura aluvial) y aquellos con influencia eólica (llanura eólica); sin embargo, omitieron el paisaje de vegas de ríos andinenses al cual pertenecen realmente los bosques de vega. Por ello, un factor que explica mejor los dos grupos encontrados es el efecto de corredor de los ríos Pauto y Meta, lo que facilita la dispersión de especies provenientes de ambas cuencas (bosques ribereños del piedemonte, de la altillanura, o la Serranía de la Macarena). Este papel de los ríos como corredores biológicos es típico en este tipo de redes hídricas y ha sido descrito ampliamente por varios autores en regiones templadas y tropicales (Johansson, et al., 1996; Naiman & Decamps, 1997; Rosales, et al., 1999).
Patrones de distribución de las especies
El hecho de que la mayoría de las especies registradas se comparta con el departamento del Meta y no con Casanare o Arauca (cuya geomorfología y clima son similares con el área estudiada), podría explicarse porque se han realizado más recolecciones de flora en el Meta, en tanto que el muestreo es bajo en Casanare y Arauca, tal como se observa en el catálogo de espermatofitas de la Orinoquía (Minorta-Cely & Rangel-Ch., 2014). También es interesante que las especies de bosques y de sabanas muestren un patrón similar de distribución en otras provincias y dominios biogeográficos de Latinoamérica, lo que reflejaría las similitudes ecológicas entre ambientes marcadamente estacionales tropicales y subtropicales, como las sabanas y los bosques secos o caducifolios de los dominios Pacífico, Mesoamericano y Chaqueño, y las selvas de tierras bajas de los dominios biogeográficos de la subregión Brasilera (dominios Mesoamericano, Pacífico, y Brasilero norte y sur). También, dicha similitud se debería a la conectividad o continuidad que perdieron las floras del norte y sur de Suramérica durante el último avance de los glaciares (Last Glacial Maximum, LGM), hace aproximadamente 18 mil años (Hooghiemstra, et al., 2002). Estos cambios paleoclimáticos produjeron una retracción pronunciada de la vegetación quedando floras fragmentarias, con importantes disyunciones (por ejemplo, Macroptilium monophyllum (Benth.) Marechal & Baudet), tal como se observa actualmente en los bosques caducifolios de ambas regiones (Banda, et al., 2016; Hooghiemstra, et al., 2002; Prado, 2000; Prado & Gibbs, 1993).
Las selvas húmedas de la Amazonía llegaban hasta el norte de Suramérica, pero su continuidad con el Chocó biogeográfico, Centroamérica y el Caribe se interrumpió casi totalmente por el levantamiento de los Andes septentrionales y las glaciaciones (Hooghiemstra & Van der Hammen, 2001). Además, cuando el clima se hizo más frío y seco durante el último máximo glacial, se produjo la contracción de las selvas a favor de la expansión de ecosistemas secos como bosques caducifolios y sabanas (Rosales, et al., 1999; Behling & Hooghiemstra, 2000; Pennington, et al., 2000). En este contexto se conectaron enclaves secos y sabanas en Suramérica permitiendo el contacto de dichas floras (Bucher, 1982; Pennington, et al., 2000). No obstante, los bosques húmedos remanentes siguieron conectados con comunidades más secas a través de los corredores ribereños (Rosales, et al., 1999; Pennington, et al., 2000). Así, el flujo de especies entre diferentes ecosistemas se mantuvo a través de la red hídrica que forman todos los cursos de agua asociados con cuencas más grandes, como la del Orinoco o la del Amazonas (Rosales, et al., 1999; Pennington, et al., 2000). Este patrón ha sido descrito en el cerrado brasileño y explica la distribución presente y pasada de la flora, que es una mezcla de selvas húmedas y ecosistemas secos en constante expansión o contracción según el clima reinante (Cardoso Da Silva & Bates, 2002).
Especies amenazadas
En cuanto a las especies evaluadas en la Orinoquía colombiana (Córdoba, et al., 2011), el número de aquellas cuya categoría de amenaza ha sido evaluada en Casanare es muy bajo (8 %), y en el río Pauto se refleja la misma tendencia, con solo tres especies incluidas en las categorías de riesgo en los libros rojos de la flora de Colombia (Calderón, et al, 2002, 2005; Cárdenas & Salinas, 2007) y solo diez citadas en los apéndices del CITES. Por otra parte, apenas 15 de las especies registradas en el presente trabajo están incluidas en la Estrategia Nacional para la Conservación de Plantas en la Orinoquía, aunque dicha lista prioriza únicamente 73 especies (Instituto Humboldt, 2013), lo que evidencia la necesidad de categorizar y proteger las especies de la cuenca baja del río Pauto, especialmente en las sabanas. Por esta razón los esfuerzos de conservación de la Orinoquía colombiana deben aumentar, pues actualmente se desconoce la categoría de amenaza de extinción de la mayoría de sus especies, lo que es crítico para adoptar medidas efectivas de protección y manejo de su hábitat y sus poblaciones.
Otras consideraciones
El presente estudio resalta la importancia de hacer estudios florísticos que consideren la heterogeneidad espacial, edáfica, e hidrológica, factores determinantes en la composición florística de bosques y sabanas de la Orinoquía (Veneklass, et al., 2005; Baruch, 2005). Se resalta, igualmente, la necesidad de que los muestreos de vegetación representen la variación climática de la Orinoquía, ya que la marcada estacionalidad de las lluvias afecta directamente la diversidad y, por ello, los trabajos que solo consideran una temporada pueden subestimarla (Mora-Fernández & Peñuela-Recio, 2013a; Mijares, et al., 2019). Cabe destacar que esta investigación, además de abarcar ambientes naturales, también contribuyó al conocimiento de la diversidad florística de zonas con transformación antropogénica, cuya salud ecosistémica es deficiente, con una reducción de la diversidad y una composición diferente a la de los ambientes de sabana naturales en la zona de estudio (Mora-Fernández & Peñuela-Recio, 2013a; Giraldo-Kalil, 2013). Es evidente, entonces, que el desarrollo de este tipo de estudios es una necesidad creciente dado el rápido deterioro que enfrentan los ecosistemas de la Orinoquía (Mora-Fernández, et al., 2015).
Conclusiones
Del total de 701 especies recolectadas, 52 no habían sido registradas para la Orinoquía; dado que la mayoría de ellas habita en las sabanas, ello refleja la necesidad de incrementar el muestreo en este tipo de ambientes. Los patrones de riqueza y composición florística de la cuenca baja del río Pauto coinciden con los reportados en estudios previos en la Orinoquía colombiana (Minorta-Cely & Rangel-Ch., 2014, Cárdenas, et al., 2016) y en la venezolana (Aymard, 2017), especialmente en cuanto a las familias, pues las de mayor riqueza en bosques ribereños y sabanas fueron las mismas. Se encontró un patrón de riqueza de las formas de crecimiento coincidente con lo reportado en la literatura; no obstante, la riqueza de hierbas en los bosques de vega del río Pauto fue mayor que la esperada en comparación con la de las especies leñosas. La composición florística en sabanas evidencia la importancia de la distribución del agua en el suelo, la topografía, y los disturbios antropogénicos en la configuración de las especies en cada unidad de vegetación, en tanto que en los bosques ribereños hubo diferencias de composición entre las áreas de influencia del río Pauto y del río Meta, probablemente debido a diferencias en los suelos y al efecto de corredor que ejercen los cauces en los que estos se desarrollan. Las especies tienen una distribución amplia en el norte de Suramérica y también en otros bosques húmedos, secos y sabanas del continente, lo que coincide con la hipótesis de continuidad de las floras antes y después del levantamiento de los Andes y el papel de los bosques ribereños como corredores de diversidad. Por último, se evidencia la necesidad de desplegar mayores esfuerzos en la categorización de amenaza de las especies de plantas de la Orinoquía, pues en el área de estudio tan solo el 2% ha sido evaluado en este sentido.

