











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
El tomate Solanum lycopersicum Mill. es la hortaliza más importante en Colombia y en el mundo dada la superficie sembrada, el volumen y el valor de la producción, así como por su consumo y el sustento económico que representa para pequeños, medianos y grandes agricultores. Sus frutos se consumen frescos y se utilizan como materia prima para la industria. En el 2018 en Colombia se cultivaron 11.227 hectáreas con un rendimiento promedio de 46,67 ton/ha y una producción total de 527.313 toneladas. (FAOSTAT, 2018). Sus frutos tienen un gran valor nutricional por el alto contenido de beta carotenos (licopeno), vitamina A y C, minerales (calcio, hierro, fósforo y potasio) y aminoácidos (tiamina y lisina). El licopeno es un poderoso antioxidante que se asocia con la baja incidencia de cáncer en humanos (Burbano & Vallejo, 2017).
El perforador del fruto Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae) es una de las plagas que más afecta la producción de tomate, causando pérdidas estimadas entre el 13 y el 77 % de los cultivos (Vallejo, 1999; Eiras & Blackmer, 2003; Almirón, et al., 2018). Inicia su ataque en los bordes del cultivo y, posteriormente, lo coloniza totalmente. Las hembras hacen la oviposición al inicio de la floración o de la formación de los primeros frutos, lo que produce un daño considerable porque, al nacer, la larva se introduce rápidamente en el fruto, lo consume por dentro y sale para transformarse en pupa dejando un orificio que permite la entrada de otros insectos o patógenos que deterioran completamente el fruto (Casas, et al., 2012). Figuras 1y2.

Figura 1 Perforador del fruto del tomate Neoleucinodes elegantalis (Guenée 1854) (Lepidoptera: Crambidae). A) Hembra. B) Macho (Casas, et aL, 2012)

Figura 2 Fruto de tomate dañado por el perforador Neoleucinodes elegantalis (Guenée 1854) (Lepidóptera: Crambidae) (Casas, et al., 2012)
En Colombia todas las variedades e híbridos del tomate son sensibles al perforador del fruto. El único método de control que están utilizando los agricultores para salvaguardar la producción comercial es la aplicación de insecticidas químicos altamente tóxicos; sin embargo, debido a que la larva perfora rápidamente el fruto después de eclosionar y permanece en su interior durante su desarrollo larval, dicho control es difícil y de cuestionable eficiencia (Salinas, et al., 1993).
La resistencia varietal, es decir la habilidad de una planta para defenderse del ataque de un insecto plaga, es la mejor forma de control, pero poco se ha estudiado y utilizado esta resistencia en el cultivo del tomate; solamente en Brasil y en Colombia se han hecho algunas investigaciones tendientes a dilucidar el mecanismo de la resistencia. Esta es una alternativa altamente deseable y ecológicamente sostenible, que disminuye el uso de insecticidas, los riesgos de contaminación ambiental y de salud, y garantiza una producción limpia y económicamente rentable (Vallejo & Estrada, 2013).
El tomate Solanum lycopersicum Mill. tiene una gran variedad de parientes silvestres que poseen resistencia a 16 insectos plagas (Rick, 1973; Vallejo, 1999) y, por lo tanto, constituyen una fuente adecuada de germoplasma para la obtención de cultivares resistentes a tales insectos. Entre estos parientes silvestres se encuentran las especies Solanum habrochaites, ubicada en el acervo genético secundario, y Solanum peruvianum, ubicada en el acervo genético terciario, las cuales se consideran posibles fuentes de resistencia al perforador del fruto del tomate. Ambas especies se distribuyen en las vertientes occidental y central de los Andes, entre el sur del Ecuador, pasando por el Perú, y la zona norte de Chile (Rick, 1973; Vallejo, 1999).
La especie S. habrochaites posee dos variedades, glabratum y typicum. Ambas poseen frutos muy pequeños de color verde con rayas o verde oscuro en estado maduro, pero la forma typicum posee tallos, hojas y frutos más pilosos y flores de mayor tamaño. La forma glabratum se cruza sin dificultad y en ambas direcciones con el tomate cultivado, en tanto que la typicum presenta incompatibilidad unilateral con el tomate cultivado, obteniéndose semilla solo cuando se emplea el tomate cultivado como progenitor femenino. Kennedy (2003) reportó que el follaje de estas dos formas era letal para varias especies de insectos plaga debido a la presencia de tóxicos como las metil cetonas (2 tridecanona y 2 undecanona), presentes en los tricomas glandulares de tipo VI.
Ortiz (2012) estudió la heredabilidad del contenido de 2 tridecanona en cruzamientos de S. lycopersicum con S. habrochaites como factores de resistencia a insectos y encontró una segregación normal en la generación F2, con valores extremos de baja frecuencia y abundancia de individuos con valores intermedios, lo que indica que se pueden seleccionar plantas con contenidos elevados de 2 tridecanona sin descuidar los caracteres de interés comercial en la búsqueda de nuevas líneas mejoradas. Los altos contenidos de 2 tridecanona en la especie S. habrochaites (cuatro veces superior a la variedad UNAPAL Maravilla) y en las plantas seleccionadas en la F2, se relacionaron con el mecanismo de resistencia de la antibiosis. La especie S. peruvianum, por su parte, es una planta alógama, autoincompatible y no se cruza fácilmente con el tomate cultivado; sus frutos son muy pequeños, de color verde o verde blanco en estado maduro (Rick, 1980).
La variedad UNAPAL Maravilla es un cultivar de tomate tipo "chonto" producido por el Grupo de investigación en mejoramiento genético y producción de hortalizas de la Universidad Nacional de Colombia. Su porte es intermedio (1,60 metros de altura), es de floración precoz y se cosecha a los 65 días. Se destaca por la alta producción por planta (8 a 9 kilos) y el peso promedio del fruto es de 150 gramos. El fruto es ovalado, bilocular o trilocular y de consistencia firme, lo que permite la fácil manipulación durante el proceso de cosecha y poscosecha. (Vallejo & Estrada, 2004). El objetivo de este trabajo fue producir líneas de tomate con resistencia genética al perforador del fruto N. elegantalis (Guenée) a partir de especies silvestres del género Solanum spp.
Materiales y métodos
Las investigaciones se realizaron en el Centro Experimental de la Universidad Nacional de Colombia, Sede Palmira, ubicado en el municipio de Candelaria, Valle del Cauca, corregimiento El Carmelo, a 3o 24' N y 76o 26' O y 980 m s.n.m., categorizado como una zona cálida moderada y semiárida, con 24 oC de temperatura promedio, 69 % de humedad relativa y 1.009 mm de precipitación promedia anual (Pardey, 2008).
Evaluación de la introducción de especies silvestres del género Solanum spp. resistentes al perforador del fruto
Se evaluó la resistencia a N. elegantalis de 12 especies silvestres del género Solanum spp. Como testigo sensible se usó la variedad UNAPAL Maravilla de la especie S. lycopersicum obtenida por la Universidad Nacional de Colombia (Tabla 1). Se utilizó el diseño experimental de bloques completos al azar con tres repeticiones; la parcela experimental estaba compuesta por siete plantas. Con el fin de obtener una población adecuada del insecto en el campo, se sembraron surcos intercalados con la variedad sensible UNAPAL Maravilla. Para calificar la resistencia al perforador se utilizó la escala propuesta por Vallejo, et al. (2008)(Tabla 2). Se cuantificó el porcentaje de frutos afectados por el perforador, el número de orificios de salida y el número de larvas por fruto. De acuerdo con el porcentaje de frutos afectados, se le otorgó una calificación a cada introducción.
Tabla 1 Introducción de especies silvestres del género Solanum spp. evaluadas para determinar la resistencia al perforador del fruto del tomate Neoleucinodes elegantalis
| Código de introducción | Especie |
|---|---|
| PI134418 | S. habrochaites var glabratum |
| PI134417 | S. habrochaites var glabratum |
| PI 126449 | S. habrochaites var glabratum |
| PI 127826 | S. habrochaites var typicum |
| PI 127827 | S. habrochaites var typicum |
| LA 1223 | S. habrochaites var typicum |
| LA 1362 | S. habrochaites var typicum |
| LA 1264 | S. habrochaites var typicum |
| LA 2092 | S. habrochaites var typicum |
| PI251305 | S. habrochaites var typicum |
| LA 0444 | S. peruvianum |
| LA 0444-1 | S. perivianum |
| UNAPAL Maravilla | S. Lycopersicum (testigo sensible) |
Fuente: Vallejo, et al., 2008
Se evaluó la respuesta del perforador del fruto a los mecanismos de resistencia de dos de las especies silvestres de S. habrochaites var. glabratum (PI 134417 y PI 134418) y una de S. habrochaites var. typicum (LA 1264) previamente caracterizadas como resistentes. Las plantas se sembraron dentro de una casa de mallas de forma circular en baldes plásticos también dispuestos en forma circular. Se utilizó el diseño completamente al azar con ocho repeticiones, así como la prueba de antixenosis o no preferencia. Se hicieron cinco liberaciones del insecto plaga y se midió el porcentaje de frutos con oviposiciones por planta, el número de huevos por fruto y el número de huevos por planta.
Tabla 2 Clasificación de las plantas de tomate según el porcentaje de frutos afectados por el perforador del fruto Neoleucinodes elegantalis
| Frutos afectados (%) | Clasificación |
|---|---|
| 0 - 5 | Muy resistente |
| 6 - 5 | Resistente |
| 6 -20 | Ligeramente sensible |
| 21 - 50 | Sensible |
| 51 -100 | Muy sensible |
Fuente: Vallejo, et al., 2008
Introgresión de la resistencia presente en la introducción PI 134418 de la especie silvestre Solanum habrochaites var. glabratum en la variedad UNAPAL Maravilla
Se utilizó la introducción PI 134418 de la especie silvestre S. habrochaites var. glabratum, muy resistente al perforador, como progenitor donante y la variedad UNAPAL Maravilla de S. lycopersicum, muy sensible al insecto, como progenitor recurrente. Se obtuvieron las generaciones F1, F2, RC1, RC2, RC3 y RC4. A partir de la RC1 se seleccionaron las plantas según su resistencia y peso promedio de fruto, con el fin de avanzar a las siguientes generaciones de retrocruzamientos.
Se evaluaron conjuntamente los progenitores P1 (introducción PI 134418 de S. habrochaites var. glabratum) y P2 (variedad UNAPAL Maravilla de S. lycopersicum) y las generaciones F1, F2, RC1, RC2, RC3 y RC4. Se utilizó el diseño experimental de bloques completos al azar con cuatro repeticiones. Se cuantificó el porcentaje de frutos afectados, el peso promedio, el color y el formato del fruto. Con base en el porcentaje de frutos afectados se otorgó una calificación a cada generación.
Resultados y discusión
Evaluación de la resistencia al perforador del fruto de especies silvestres del género Solanum spp.
De las 12 introducciones de especies silvestres evaluadas, solamente tres de S. habrochaites var. glabratum (PI 134417, PI 134418 y PI 126449), dos de S. habrochaites var. typicum (LA 1264 y LA 2092) y una de S. peruvianum (LA 444-1) lograron crecer, desarrollarse y fructificar normalmente. Las otras seis presentaron anormalidades en el crecimiento y el desarrollo y, por lo tanto, no alcanzaron la fructificación, probablemente debido a las diferentes condiciones ambientales del sitio de evaluación en comparación con la región de origen de estas especies y su alto grado de autoincompatibilidad (Rick, 1971).
De las introducciones silvestres que alcanzaron a fructificar, ninguna presentó frutos afectados o dañados por el perforador, en tanto que el 60,3 % de los frutos del testigo sensible UNAPAL Maravilla se vio afectado por el perforador. Ninguna de las introducciones silvestres presentó frutos con orificios de salida de larvas ni larvas dentro del fruto, en tanto que el testigo sensible presentó tres orificios de salida y dos larvas dentro del fruto, lo que indicaba que cada fruto fue atacado, en promedio, por cinco larvas del perforador (Tabla 3).
Tabla 3 Resistencia al perforador del fruto Neoleucinodes elegantalis en introducciones de especies silvestres del género Solanum spp.
| Introducción | Frutos Afectados (%) | Orificios de salida | Frutos con orificios de salida (%) | Larvas por fruto | Clasificación |
|---|---|---|---|---|---|
| PI 134417 | 0,0 | 0,0 | 0,0 | 0,0 | Muy resistente |
| PI 134418 | 0,0 | 0,0 | 0,0 | 0,0 | |
| PI 126449 | 0,0 | 0,0 | 0,0 | 0,0 | |
| LA 1264 | 0,0 | 0,0 | 0,0 | 0,0 | |
| LA 2092 | 0,0 | 0,0 | 0,0 | 0,0 | |
| LA 444-1 | 0,0 | 0,0 | 0,0 | 0,0 | |
| Testigo | 60,30 | 3,00 | 57,20 | 2,00 | Muy sensible |
Fuente: Vallejo, et al., 2008
Con base en esta información, las introducciones PI 134417, PI 134418 y PI 126449 de la especie silvestre S. habrochaites var. glabratum, LA 1264 y LA 2092 de S. habrochaites var. typicum y LA 444-1 de S. peruvianum, se clasificaron como muy resistentes al perforador del fruto, en tanto que la variedad UNAPAL Maravilla se clasificó como muy sensible al insecto (Tabla 3).
En 1993, Salinas, et al . reportaron una introducción de la especie S. habrochaites var. glabratum como altamente resistente al perforador del fruto, por cuanto no presentaba ningún fruto afectado. Parra, et al. (1997) evaluaron por su parte especies silvestres de Solanum spp. como posibles fuentes de resistencia al insecto Scrobipalpula absoluta y concluyeron que las introducciones silvestres de S. habrochaites var. typicum fueron resistentes, lo que sugiere que dicha resistencia estaba relacionada con el tamaño pequeño del fruto y su alta pubescencia, que dificultaba la ovoposición del insecto adulto y la locomoción de la larva en el fruto.
Salazar (2013) evaluó cinco introducciones de tomate tipo Cherry, con el fin de determinar su grado de resistencia al perforador del fruto usando como testigo resistente la especie silvestre S. habrochaites var. glabratum y como testigo sensible el híbrido comercial FT 006. Los resultados indicaron que la especie silvestre no presentó frutos afectados por el insecto plaga, confirmando, una vez más, la resistencia, en tanto que las introducciones de tomate Cherry presentaron menor incidencia en comparación con el testigo sensible, el híbrido FT 006.
Mecanismos de resistencia al perforador del fruto en especies silvestres del género Solanum spp.
En cuanto al número de frutos con oviposición por planta, se observaron diferencias significativas entre las tres introducciones silvestres y el testigo sensible UNAPAL Maravilla. Las introducciones PI 134417, PI 134418 y LA 1264 presentaron la menor oviposición de frutos por planta (7,3, 6,8 y 0,5, respectivamente) en comparación con el testigo, con 70,0 frutos con oviposición por planta (Tabla 4).
Tabla 4 Frutos por planta con oviposiciones, número de huevos por fruto y número de huevos por planta de Neoleucinodes elegantalis en diferentes introducciones de especies de Solanum spp. en condiciones de casa de malla
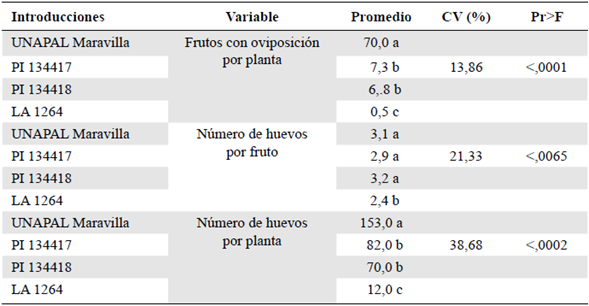
CV= Coeficiente de variación; Pr= Probabilidad; F = Prueba F de Fisher; Los promedios con la misma letra no fueron significativamente diferentes según la prueba de diferencia mínima significativa (DMS) (p<0,01). Fuente: Casas, et al., 2012.
Estos resultados confirman la gran resistencia de las introducciones silvestres y la marcada sensibilidad del testigo UNAPAL Maravilla. La oviposición tan reducida en las introducciones silvestres, comparada con la encontrada en el testigo sensible, sugiere un efecto antixenótico en el perforador del fruto.
En cuanto al número de huevos por fruto, se presentaron diferencias significativas entre las introducciones. La introducción LA 1264 presentó un valor significativamente menor comparado con las introducciones PI 134417 y PI 134418 y con el testigo UNAPAL Maravilla, y no se presentaron diferencias significativas entre las introducciones PI 134417, PI 134418 y el testigo UNAPAL Maravilla (Tabla 4). Es importante mencionar que el número de huevos por fruto está condicionado por la actividad reproductiva natural del insecto, sobre la cual no es posible tener control.
En el número de huevos por planta se presentaron diferencias significativas entre el testigo sensible UNAPAL Maravilla y las tres introducciones silvestres del género Solanum spp. En el testigo sensible se reportó un promedio significativamente mayor que el promedio de las introducciones PI 134417, PI 134418 y LA 1264. Asimismo, la introducción LA 1264 presentó un valor significativamente menor que el de las introducciones PI 134417 y PI 134418.
Hay evidencia de la presencia de diferentes mecanismos de resistencia que le permiten a las especies silvestres del género Solanum spp. defenderse de diversos insectos plagas, entre ellos, las barreras físicas (tricomas) y las químicas (sustancias tóxicas en los tricomas y liberación de volátiles repelentes) (Vallejo, 1999). Luckwill (1943) identificó en el género Solanum spp. siete tipos de tricomas, los no glandulares (tipos II, III y V) y los glandulares (tipos I, IV, VI y VII). Kennedy (2003) y Simmonds & Gurr (2005) mencionaron que los tricomas glandulares y no glandulares en las plantas del género Solanum spp. son componentes fundamentales de su defensa frente a insectos plagas.
Por su parte, De Sousa, et al. (2002) estudiaron los metabolitos secundarios en las especies silvestres de Solanum como responsables de los mecanismos de resistencia al perforador del fruto del tomate. Encontraron que los tricomas de tipo VI presentes en la especie S. habrochaites var. glabratum están asociados con el metabolito 2 tridecanona, el cual es extremadamente tóxico para los huevos y las larvas neonatas.
En las introducciones de especies silvestres del género Solanum spp. los huevos se encontraron adheridos a los tricomas de tipo I, lo que sugiere un efecto antixenótico sobre el insecto adulto de N. elegantalis, dado que se presentó una alteración en su comportamiento reproductivo comparado con la variedad UNAPAL Maravilla, en la cual la ovoposición se presentó sobre el fruto. Las larvas que eclosionaron de los huevos depositados en los tricomas de tipo I morían rápidamente y muy cerca de los coriones al descender y llegar a la superficie del fruto donde se encuentran los tricomas de tipo IV y VI. Asimismo, se detectó que algunas larvas que intentaban escapar de la superficie del fruto ascendían nuevamente por los tricomas de tipo I, donde morían después de un corto tiempo. Estas reacciones de las larvas neonatas sugieren un posible efecto antibiótico.
Por otra parte, se observaron algunas pocas posturas sobre la superficie de los frutos de las introducciones de especies silvestres PI 134417, PI 134418 y LA 1264, cuyas larvas eclosionadas morían a pocos milímetros de los coriones, lo que indica una alteración en la biología del insecto relacionada con un posible efecto antibiótico sobre su primer estadio de desarrollo.
Los tricomas glandulares de tipo I, IV y VI presentes en las tres especies silvestres introducidas atraparon a las larvas neonatas del perforador, limitando sus movimientos y ocasionándoles la muerte por la acción de los metabolitos secundarios presentes en ellos.
En este sentido, Simmonds, et al. (2004) mencionaron que al liberarse los contenidos de los tricomas glandulares 2 tridecanona y 2 undecanona y entrar en contacto con la plaga pueden causarle irritación, atraparla e, inclusive, matarla.
Las reacciones negativas observadas en el comportamiento reproductivo y la biología del insecto evidenciaron un posible efecto antibiótico y antixenótico (conferido principalmente por la presencia de tricomas glandulares de tipo IV y VI en los frutos de las introducciones silvestres PI 134417, PI 134418 y LA 1264) sobre el perforador del fruto N. elegantalis.
Introgresión de la resistencia presente en la introducción PI 134418 de la especie silvestre Solanum habrochaites var. glabratum en la variedad UNAPAL Maravilla de la especie Solanum lycopersicum Mill.
Se produjo semilla híbrida solamente cuando se cruzó UNAPAL Maravilla como progenitor femenino con tres introducciones silvestres de S. habrochaites var. glabratum (PI 134417, PI 134418 y PI 126449) y con tres de S. habrochaites var. typicum (PI 127826, PI 126445 y LA 1264) usadas como progenitores masculinos (Tabla 5), lo que indicaba la presencia de compatibilidad unidireccional. Los cruzamientos con las introducciones silvestres de S. peruvianum no produjeron semilla híbrida debido a la marcada barrera de incompatibilidad, ya que esta especie pertenece al acervo genético terciario (Rick, 1980).
Tabla 5 Eficiencia de cruzamiento, germinación de la semilla y número de semillas normales y anormales en los cruzamientos interespecíficos entre la variedad UNAPAL Maravilla, Solanum lycopersicum y las introducciones silvestres del género Solanum spp.
| Cruzamiento | Número de cruzamientos | Eficiencia de cruzamiento (%) | Número de semillas normales/anormales | Germinación de semilla (%) |
|---|---|---|---|---|
| UNAPAL x PI 134417 | 274 | 11,7 | 320-219 | 21,5 |
| UNAPAL x PI 134418 | 396 | 6,3 | 279-119 | 32,7 |
| UNAPAL x PI 127826 | 217 | 7,4 | 320- 55 | 6,9 |
| UNAPAL x PI 126445 | 150 | 6,0 | 111- 33 | 11,5 |
| UNAPAL x PI 126449 | 218 | 11,9 | 432-187 | 57,5 |
| UNAPAL x LA 1264 | 156 | 5,8 | 82- 24 | 39,3 |
Fuente: Restrepo, et al., 2007
La eficiencia de los cruzamientos fue muy baja, fluctuó entre 5,8 y 11,9 %, posiblemente porque las dos variedades de S. habrochaites pertenecen al acervo genético secundario, y se presenta cierto grado de incompatibilidad (Moyle & Graham, 2005).
En todas las hibridaciones interespecíficas se produjo una mayor cantidad de semillas normales (entre 82 y 320) que anormales (entre 24 y 219), pero la germinación fue relativamente baja, fluctuando entre 6,9 y 57,5 %; sin embargo, esto permitió realizar sucesivos retrocruzamientos para obtener líneas de tomate con resistencia al perforador del fruto.
La eficiencia del retrocruzamiento uno (RC1) fue muy baja, fluctuando entre 6,5 y 9,1 %, por lo cual se necesitaron muchos retrocruzamientos para producir semilla híbrida en cantidades adecuadas. El problema se agravó cuando se avanzó al retrocruzamiento dos (RC2), en el cual se obtuvo semilla únicamente en el retrocruzamiento con la introducción silvestre PI 134418 (61 semillas), con una eficiencia del 1,5 % (Tabla 6), lo que se debió, posiblemente, a la presencia de incompatibilidad en la introducción silvestre y a la distancia genética con el tomate cultivado S. lycopersicum. Al estudiar la base genética de la incompatibilidad de híbridos entre S. lycopersicum y S. habrochaites,Moyle & Graham (2005) encontraron una reducción importante en la fertilidad del polen de las plantas híbridas en comparación con la de los progenitores, lo que probablemente sea la causa de la poca eficiencia del cruzamiento y la baja producción de semillas híbridas.
Tabla 6 Eficiencia de los retrocruzamientos uno y dos (RC1 y RC2) y número de semillas normales obtenidas
| Padre donante | RC1 | RC2 | ||
|---|---|---|---|---|
| Eficiencia (%) | Semillas normales | Eficiencia (%) | Semillas normales | |
| PI134417 | 8,0 | 363 | 0,0 | 0,0 |
| PI134418 | 9,1 | 302 | 1,5 | 61,0 |
| PI 126449 | 6,5 | 429 | 0,0 | 0,0 |
| LA 1264 | 8,1 | 102 | 0,0 | 0,0 |
Fuente: Restrepo, et al., 2007
Al evaluar los progenitores y las generaciones F1, RC1, RC2, RC3 y RC4, se observó que el daño promedio del perforador fue mayor en el padre recurrente UNAPAL Maravilla (57,52 %) que en el padre donante PI 134418 de la especie S. habrochaites var. glabratum (1,27 %), con la generación híbrida F1 (7,14 %) y con todas las generaciones de retrocruzamiento, el cual fluctuó entre 8,20 y 14,22 %, por lo que se puede sugerir que el método de retrocruzamiento con selección fue efectivo para mantener cierta cantidad de la resistencia otorgada por el progenitor PI 134418 de S. habrochaites var. glabratum (Tabla 7).
Tabla 7 Frutos afectados por el perforador del fruto (%), peso promedio del fruto (g), formato y color del fruto maduro en generaciones P1 (S. habrochaites var. glabratum, introducción PI 134418), P2 (UNAPAL Maravilla), y F1, RC1, RC2, RC3 y RC4
| Generación | Plantas seleccionadas | Promedio de frutos afectados (%) | Peso promedio de fruto (g) | Densidad tricomas | Color de fruto | Formato de fruto |
|---|---|---|---|---|---|---|
| P1 | - | 1,27 a | 2,37 a | 5,0 a | Verde | Redondo |
| P2 | - | 52,52 b | 136,04 b | 2,0 b | Rojo | Chonto |
| F1 | - | 7,14 c | 17,88 c | 4,0 a | Amarillo/rojo | Ovoide |
| RC1 | 3 | 13,23 d | 16,98 c | 2,6 b | Amarillo/rojo | Ovoide |
| RC2 | 2 | 14,22 d | 30,85 d | 2,6 b | Rojo | Ovoide |
| RC3 | 9 | 8,20 e | 79,71 e | 2,6 b | Rojo | Chonto |
| RC4 | 3 | 9,76 e | 74,67 e | 2,6 b | Rojo | Chonto |
Los promedios seguidos por la misma letra no son significativamente diferentes según la prueba de diferencia mínima significativa (DMS) (p<0,01). Fuente: Pérez, 2010
Asimismo, a medida que se avanzó en las generaciones de retrocruzamientos con selección se fue recuperando el peso promedio del fruto, el color rojo intenso y el formato tipo chonto del progenitor recurrente UNAPAL Maravilla. En la generación RC1 el peso promedio del fruto fue de 16,98 gramos, el color varió entre amarillo y rojo y el formato fue ovoide, y en la generación RC4, el peso promedio del fruto fue de 74,67 gramos, el color fue rojo y el formato fue tipo chonto, lo que nuevamente demuestra que el método de mejoramiento por retrocruzamiento con selección fue efectivo para recuperar el peso promedio del fruto, su color y el formato del progenitor recurrente (Tabla 7).
Al analizar individualmente las plantas del retrocruzamiento RC4, se logró identificar una recombinante con un 4,65 % de frutos afectados por el insecto plaga, peso promedio de 130,67 gramos, color rojo intenso y formato de fruto tipo chonto, lo cual indicó que se logró la introgresión de la resistencia de la especie silvestre S. habrochaites var. glabratum a la variedad comercial UNAPAL Maravilla, y se pudieron recuperar las buenas características agronómicas de esta variedad.
Vallejo, et al. (2008) reportaron una asociación positiva y muy significativa entre el peso promedio del fruto y el número de frutos afectados por el perforador (0,85), lo que indica que a medida que se recuperaba el peso promedio del fruto se aumentaba el número de frutos afectados; sin embargo, Pérez (2010) demostró que la asociación positiva entre una mayor sensible a la plaga y un mayor peso del fruto se pudo romper con el método de retrocruzamiento acompañado de la selección, que se obtuvo en la RC4 y en líneas recombinantes resistentes con peso promedio, color y formato del fruto similares al del padre recurrente UNAPAL Maravilla. En el grupo de investigación se continúa avanzando en generaciones de retrocruzamiento con selección para recuperar aún más el peso promedio del fruto y mantener la resistencia al perforador del fruto y otras características agronómicas del padre recurrente deseables.
En cuanto a la densidad de tricomas en el fruto, se observó una diferencia altamente significativa entre los dos progenitores y que la introducción silvestre PI 134418 presentó una densidad de 5,0 tricomas por milímetro cuadrado, en tanto que la de la variedad UNAPAL Maravilla fue de 2,0 tricomas por milímetro cuadrado. Las cuatro generaciones de retrocruzamiento (RC1, RC2, RC3 y RC4) no presentaron diferencias significativas entre sí ni con el progenitor recurrente UNAPAL Maravilla (Tabla 5), lo que indica que a medida que se recuperaba el peso promedio del fruto, la densidad de los tricomas permanecía baja (2,6 tricomas por milímetro cuadrado) y estadísticamente igual a la del progenitor recurrente UNAPAL Maravilla (2,0 tricomas por milímetro cuadrado); ello sugiere que los tricomas no necesariamente son los responsables de la resistencia al perforador del fruto.
Vallejo, et al. (2008) reportaron resultados diferentes y mencionan que el peso promedio del fruto presentó una asociación negativa y significativa (-0,59) con la densidad de tricomas, lo que sugería que a medida que aumentaba el peso promedio del fruto, se disminuía la densidad de tricomas, es decir, que los tricomas probablemente no son los únicos responsables de la resistencia al perforador del fruto.
Conclusiones
La resistencia genética al perforador del fruto del tomate presente en las especies silvestres de Solanum spp. puede ser transferida por hibridación sexual a la variedad UNAPAL Maravilla de la especie S. lycopersicum con mayor o menor éxito. Los tricomas glandulares presentes en las introducciones silvestres del género Solanum, y las sustancias químicas asociadas con ellas, ejercieron un efecto antixenótico y antibiótico en el perforador del fruto. El retrocruzamiento y la selección fueron efectivos para producir líneas recombinantes en la generación RC4, con resistencia al perforador y con peso promedio, color y formato del fruto similares a los del padre recurrente.

