











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
Introducción
La acelerada transformación y pérdida de ecosistemas y la consecuente erosión de la biodiversidad imponen retos cada vez mayores a quienes trabajan por su adecuado manejo y conservación. El número de especies consideradas en peligro de extinción a nivel global es cerca de un millón (Díaz, et al., 2019) y se ha demostrado que un gran número de poblaciones de vertebrados está en declive (Green, et al., 2019; Ceballos, et al., 2017). Colombia también ha experimentado estos procesos, y aunque más de la mitad del territorio nacional está aún cubierto con bosques, los motores de transformación han afectado cerca del 50 % de los ecosistemas (Etter, et al., 2017) y más de 1.500 especies de fauna y flora están amenazadas (Andrade-Correa, 2011).
Para dar respuesta a estos procesos de transformación y pérdida de biodiversidad, muchos países han implementado numerosas iniciativas de conservación, con mayor o menor éxito. Gran parte de estos esfuerzos se han enfocado en la conservación de especies particulares (single-species), en parte influenciados por el Endangered Species Act (ESA) de los Estados Unidos, una política que obliga al estado a tomar acciones de conservación de las especies (y sus hábitats) que se encuentren en dicha lista de riesgo.
Sin embargo, algunos investigadores han cuestionado la aproximación basada en especies, argumentando que no es posible salvarlas a todas con los recursos limitados que existen. La única alternativa, sugieren, es pasar a una aproximación de paisaje, en donde al manejar o conservar a una escala más amplia, se lograría salvar a todos los componentes de un sistema a través de una estrategia unificada (Franklin, 1993). Esta aproximación tiene cada vez más adeptos, aunque aún presenta ciertas dificultades conceptuales y operativas (Goldstein, 1999a; Simberloff, 1998). Una de las características principales del manejo a nivel de ecosistemas es el enfoque centrado en procesos ecológicos y no en especies individuales, pero no está claro cuáles procesos son más importantes, o cómo priorizarlos o medirlos (Tracy & Brussard, 1994). Tampoco es claro cómo la protección de ciertos procesos asegura los requerimientos de los organismos que se quieren mantener (Goldstein, 1999a). En muchos casos, es posible mantener procesos a expensas de perder especies, algo que no es lo ideal desde una perspectiva de conservación.
Por estas razones, algunos autores consideran el manejo de ecosistemas como un "atajo", que subestima el rol de las especies y los requerimientos de sus poblaciones en la construcción de los planes de manejo (Goldstein, 1999b). Además, aducen que los objetivos definidos en la aplicación de dicha aproximación tienden a ser vagos ("proteger la diversidad", "mantener los procesos ecológicos"), sin hipótesis claras y evaluables, y sin una medición objetiva del avance en su consecución. Tales críticas no implican un desconocimiento del valor e importancia del estudio del nivel ecosistémico, sin embargo, para que el manejo de ecosistemas sea útil en la práctica, los criterios que utiliza deben reflejar las necesidades de los organismos que busca proteger, más que propiedades emergentes, funciones no definidas claramente o procesos de grupos de organismos (Goldstein, 1999a; Goldstein, 1999b).
La conservación basada en especies, por su parte, tiene sus propios retos, siendo el principal la imposibilidad de estudiar y manejar todas las especies de un sistema (Simberloff, 1998). Por esto se hace necesario seleccionar elementos que representen las condiciones del sistema y que permitan la implementación de acciones de manejo y conservación. Algunas propuestas iniciales basadas en especies han intentado responder este reto (especies indicadoras, especies sombrilla, especies bandera). Al estudiar y reconocer las limitaciones de cada una de estas propuestas, el enfoque de especies ha ido evolucionando para conciliar mejor la teoría ecológica con la práctica. Uno de los primeros conceptos fue el de especie indicadora, basado en la premisa de que su presencia o fluctuación refleja la de otras especies de la comunidad (Landres, et al., 1988; Simberloff, 1998). Cuando se aspira a determinar la tendencia de otras especies, sería necesario evaluar la presencia concurrente de la especie indicadora y estas otras especies, o verificar que las acciones en pro de la conservación de las primeras inciden en las poblaciones de las segundas (Simberloff, 1998). Estas evaluaciones generalmente no se realizan, lo que resulta en que las especies indicadoras (en conservación) se basen, generalmente, en supuestos.
Las especies sombrilla, un concepto más amplio, son especies con requerimientos de hábitat muy grandes, por lo que su conservación automáticamente salvaría a otras especies (Wilcox, 1984; Roberge & Angelstam, 2004). De nuevo, este concepto requiere datos que lo soporten, pues salvar a la especie sombrilla no necesariamente salvará a todas las otras especies del ecosistema, que pueden tener requerimientos de hábitat muy específicos.
Por otra parte, las especies bandera (flagship species) son un concepto con objetivos orientados más a la gestión que a lo ecológico. Son aquellas especies que, por su carisma, reconocimiento y aprecio por parte de los humanos, se usan como símbolo de programas de conservación con el objetivo de involucrar al público y llevar un mensaje (Shrader-Frechette & McCoy, 1993). Si bien el método es popular y puede ser útil en la consecución de recursos, se basa únicamente en el potencial de la especie para cautivar al público. En este sentido, este concepto, al igual que los anteriores, carece del soporte ecológico que demuestre que su conservación se traduce en beneficios para otras especies (Caro, et al., 2004). Otra limitación potencial de la aplicación de este concepto es que no todas las regiones o territorios cuentan con especies llamativas y carismáticas que puedan ser usadas como especie bandera en programas de conservación.
Una aproximación novedosa en su momento, fue el uso del concepto de especie clave, o piedra angular (keystone species) en conservación (Mills, et al., 1993; Simberloff, 1998), el cual alude a aquellas especies que tienen una función esencial para mantener la estructura o composición de un sistema, más allá de lo esperado por su abundancia (Paine, 1969). Al conservar estas especies, se estarían conservando sus funciones y dichos mecanismos de regulación. De cierta forma, este concepto intenta conjugar la aproximación de especies con la de procesos ecosistémicos. No obstante, la identificación y comprobación de especies clave y sus funciones no es sencilla, y no sabemos cuántos ecosistemas contienen especies clave que puedan ser objeto de conservación.
Una aproximación que busca conciliar el enfoque centrado en las especies con el de ecosistemas es la conservación basada en especies paisaje. En este modelo se selecciona un conjunto de especies cuyas características biológicas, requerimientos espaciales e importancia para las comunidades humanas permiten determinar y abordar actividades que tienen impacto en la biodiversidad en el paisaje (Sanderson, et al., 2002). Este tipo de conservación se diferencia de otras aproximaciones basadas en especies en el hecho de que se enfoca no solo en la selección de las especies sino en el proceso de planificación, detección de las amenazas y las estrategias necesarias para su mitigación. Las especies paisaje se definen como aquellas que usan áreas extensas, ecológicamente diversas, y que usualmente tienen un impacto significativo en la estructura y función de los ecosistemas, pero son susceptibles a las alteraciones humanas de los paisajes (Sanderson, et al., 2002). Los cinco criterios para elegir el conjunto de especies paisaje son: requerimientos de área, heterogeneidad de hábitats, función ecológica, vulnerabilidad e importancia socioeconómica (Coppolillo, et al., 2004). Con base en las especies seleccionadas, se construyen modelos conceptuales que identifican las presiones a que están sometidas sus poblaciones, los factores que contribuyen a estas presiones y las estrategias para mitigarlos en el paisaje. Así, este modelo de trabajo considera los factores biológicos y sociales que inciden sobre el estado de las especies y ecosistemas, y viceversa. La conservación de especies paisaje reconoce, entonces, que los organismos pueden existir en espacios heterogéneos que también satisfacen requerimientos económicos y culturales de las personas.
Este tipo de conservación se basa, no obstante, en una serie de supuestos, siendo el principal que al proteger y cumplir con las necesidades de hábitat y supervivencia de las poblaciones de especies paisaje y reducir sus presiones a través de las acciones de conservación, se logrará la conservación de otras especies y del paisaje en general (Sanderson, et al., 2002). En teoría, la conservación de las especies paisaje ayudaría también a mantener la estructura de los ecosistemas y, por lo tanto, la función derivada de esa estructura. Así, es necesario estimar la validez de estos supuestos y desarrollar mecanismos de evaluación del impacto de las iniciativas de conservación implementadas bajo dicha aproximación. En este estudio proponemos un modelo de planificación basado en especies paisaje y alternativas para evaluar la efectividad en su conservación y la de otros elementos de la biodiversidad (especies de vertebrados en este caso), utilizando una aproximación cuantitativa.
Materiales y métodos
Para describir la planificación en el marco del modelo y evaluar su impacto en la conservación de la biodiversidad utilizamos como ejemplo el "Proyecto Vida Silvestre", una iniciativa de conservación de la biodiversidad a escala de paisaje desarrollada en dos regiones del país, la Orinoquia y el Magdalena Medio. En la planificación e implementación de las acciones en cada región se desarrollaron los siguientes pasos: 1) selección del conjunto complementario de especies paisaje, 2) definición del paisaje de conservación, 3) planificación de estrategias de conservación, 4) implementación de acciones de conservación y 5) diseño e implementación de un programa de monitoreo.
Las especies paisaje en cada región se seleccionaron a partir de una lista de especies candidatas (Corzo, et al., 2010), con apoyo del programa Landscape Species Selection -LSS (Strindberg & Didier, 2006). Se compiló información secundaria en torno a cada especie para evaluar los cinco criterios mencionados y calcular un puntaje agregado. La especie con mayor puntaje se seleccionó como la primera, en tanto que las siguientes se seleccionaron una a una en iteraciones consecutivas. En cada iteración se estableció la especie más complementaria a las ya seleccionadas, es decir la que cubría nuevos (distintos) hábitats y amenazas, y así sucesivamente hasta completar todos los hábitats y amenazas detectadas en el paisaje (Coppolillo, et al., 2004; Strindberg & Didier, 2006).
En el paisaje del Magdalena Medio, partiendo de una lista de 65 especies candidatas, se seleccionaron el paujil de pico azul (Crax alberti), el choibo (Ateles hybridus), el manatí del Caribe (Tricechus manatus), el bagre del Magdalena (Pseudoplatystoma magdaleniatum) y el árbol del carreto (Aspidosperma megalocarpon). En el paisaje de la Orinoquia las especies seleccionadas fueron la danta de tierras bajas (Tapirus terrestris), el caimán llanero (Crocodylus intermedius), la tortuga charapa (Podocnemis expansa), la palma de moriche (Mauritia flexuosa) y el árbol de congrio (Acosmium nitens). Una vez seleccionadas las especies, se delimitó el paisaje de conservación dentro de cada región considerando su distribución y la presencia de actividades humanas. En cada región se incluyeron las áreas con presión debida a la deforestación con base en los datos de IDEAM (Galindo, et al., 2014). El paisaje del Magdalena Medio se ubica en los municipios de Yondó, en Antioquia, y Puerto Parra y Cimitarra en Santander. Este paisaje tiene un área de 2.536 Km2, de los cuales el 57 % es territorio agrícola y 5 %, humedales. Presenta ecosistemas fragmentados por actividades humanas, pero contiene relictos de bosque húmedo tropical que albergan una gran diversidad biológica. El paisaje de la Orinoquia incluye partes de la cuenca media y baja del río Bita, en Vichada, en un área de 4.074 Km2, de los cuales el 80 % corresponde a sabanas y bosques naturales, y tan solo el 1 %, a actividades agrícolas.
Para planificar las estrategias de conservación de cada paisaje, construimos de forma participativa modelos conceptuales. Un modelo conceptual es una representación gráfica de la situación en un área particular, que muestra lo que el proyecto aspira a lograr, los factores que tienen un impacto negativo (o positivo) sobre las especies seleccionadas, y cómo las acciones de conservación proyectadas abordan las amenazas y logran resultados (The Conservation Measures Partnership, 2020). Con base en los modelos conceptuales se construyeron las estrategias de conservación en cada paisaje. El trabajo y la participación de las comunidades locales fue un eje transversal en todo el proceso.
Con el fin de evaluar el impacto de las acciones en la conservación de las especies paisaje y la biodiversidad, se diseñó e implementó un programa de monitoreo entendido como un componente integral de la adopción de decisiones basado en modelos de respuesta del sistema a las intervenciones, o monitoreo para la conservación activa (Nichols & Williams, 2006). El programa contiene indicadores de gestión, de resultados y de impacto. En el presente estudio nos enfocamos en el impacto de las actividades implementadas en la biodiversidad. Otros indicadores se describirán en detalle en publicaciones futuras.
Evaluamos dos hipótesis fundamentales en el marco de esta aproximación: 1) que las acciones de conservación derivadas de la planificación adecuada con base en las especies paisaje tienen un efecto positivo en sus poblaciones (abundancia estimada como ocupación) y en otros elementos de la biodiversidad, y 2) que la estrategia de conservación en el paisaje tiene efectos positivos en la estructura de los ecosistemas medida, en este caso, a través de la reducción en la deforestación.
Para evaluar la primera y principal hipótesis del modelo, utilizamos una aproximación cuantitativa basada en modelos de ocupación multitemporada a partir de datos provenientes de trampas cámara y transectos. Estos modelos son un método para describir la ocupación (occupancy) o la proporción de sitios ocupados por una especie durante un intervalo de tiempo limitado cuando la probabilidad de detectar dicha especie es menor a uno (Mackenzie, et al., 2003). Los modelos de ocupación permiten estimar parámetros como la colonización y la extinción en los sitios evaluados, para lo cual se requieren observaciones replicadas de detección o no detección en las unidades espaciales de muestreo. En cada paisaje se sobrepuso una cuadrícula de celdas de 1x1 Km2, siendo la celda la unidad espacial de análisis. De cada cuadrícula (paisaje) se seleccionaron y muestrearon 60 celdas, cada una con más del 10 % de su área boscosa. Se realizaron temporadas anuales de muestreo con cámaras trampa (Reconyx HC500) en el 2015 y el 2017 durante la época seca (enero-mayo), con un muestreo efectivo de por lo menos 45 noches por cámara y por temporada. Para los análisis, se dividieron los datos de las cámaras trampa en periodos de 15 días, de manera que cada uno representaba una ocasión de muestreo. Para complementar el muestreo, en cada celda se recorrieron tres transectos de 800 m en dos ocasiones (en total seis réplicas: dos temporales y tres espaciales), en los que se registraron encuentros visuales (primates) y señales y rastros de mamíferos terrestres. La información referente a los ungulados se analizó a partir de los dos métodos combinados (cuatro réplicas para cámaras y seis de transectos), en tanto que para Ateles hybridus solo se tuvieron en cuenta los seis transectos por celda. Las demás especies se trabajaron solo con los datos de las cámaras trampa.
Para cada especie se generaron modelos de ocupación multitemporada (Fiske, et al., 2011; Mackenzie, et al., 2003; MacKenzie, et al., 2006), los cuales proporcionan estimaciones de una ocupación dinámica que tiene en cuenta la ocupación inicial (ψ1) y las tasas de colonización (y) y extinción (ε) promedio por sitio, lo que permite estimar los cambios en la ocupación entre dos periodos de tiempo (2015-2017), corrigiendo por la probabilidad de detección (p) por temporada (Fiske, et al., 2011; Mackenzie, et al., 2003; MacKenzie, et al., 2006). Estos modelos permiten establecer las covariables que afectan las probabilidades de colonización (que una celda no ocupada pase a ser ocupada) o extinción (que una celda ocupada deje de estarlo). De esta forma se evaluaron de manera explícita las relaciones de causalidad entre la intervención (acuerdos) y los procesos ecológicos de ocupación de un área.
Para determinar el efecto de los acuerdos de conservación en la ocupación de cada una de las especies, se generaron modelos para evaluar el efecto del área bajo acuerdo en las tasas de colonización y extinción en cada celda. El área bajo acuerdo correspondía al área incluida en los acuerdos de conservación firmados con los propietarios de predios. En los predios con acuerdos se establecieron compromisos para reducir o eliminar la cacería de especies silvestres y proteger los ecosistemas naturales, y se fomentaron prácticas sostenibles de producción como incentivo. Además, en los modelos se incluyó el área de bosque presente en el 2017 como un factor ecológico que podría influir en la ocupación de las especies. Se construyeron y evaluaron 16 modelos para cada una de las especies, incluidas las combinaciones posibles entre las dos variables (área de acuerdo, área de bosque) para ambos parámetros (colonización y extinción), que representaban hipótesis viables y relevantes, y los modelos nulos en los que no existía un efecto de las covariables en los parámetros. Los modelos se ajustaron utilizando el paquete Unmarked en el programa R (Fiske, et al., 2011). Para determinar el modelo que representaba de mejor manera los datos, se utilizaron el criterio de información de Akaike corregido (ωICc) y los pesos AIC (coAIC) para cada especie (puntaje AIC más bajo y ω más alto) (Burnham & Anderson, 2002). Para comparar el efecto de cada variable en las tasas de colonización y de extinción de cada especie, se sumaron los pesos de AIC para todos los modelos que contenían la variable de interés como una medida de su peso relativo (Arnold, 2010).
Para evaluar la segunda hipótesis, se estimó el impacto de las acciones en la deforestación ocurrida en el período de estudio. No se midió directamente la estructura de los ecosistemas, pero se usó como medida indirecta la deforestación, asumiendo que reducir la deforestación implica un mantenimiento de la estructura. Debido a que algunas intervenciones se realizaron en áreas específicas (predios con acuerdos), pero otras tuvieron alcance en todo el paisaje (fortalecimiento comunitario, educación ambiental, etc.), se evaluó el efecto en las dos escalas espaciales: de predio y de paisaje. Para determinar el efecto a escala de paisaje, se compararon las tasas de pérdida de bosque dentro de cada paisaje (el área que engloba las áreas de intervención) y un área de amortiguamiento de 5 km alrededor de cada uno (Magdalena Medio y Orinoquia), con los mismos ecosistemas y condiciones similares, reduciendo así la heterogeneidad en variables asociadas a la deforestación (Armenteras, et al., 2009). Para esta comparación se usó la información sobre pérdida de bosques de Hansen, et al. (2013), con una resolución de 30 m2, entre el 2015 y el 2018. Este método permite evaluar el impacto de las intervenciones en el paisaje en la reducción de las tasas de deforestación al compararlas con lo ocurrido en un área con condiciones similares y en ausencia de las intervenciones. Usamos estadísticas no paramétricas y pruebas de Wilcoxon pareadas debido al bajo número de réplicas (4 años) y la ausencia de normalidad en los datos de tasas de deforestación. En esta primera aproximación, hubo un valor de deforestación pareado por año (el paisaje y su zona de amortiguamiento) y las réplicas (pseudoréplicas) fueron los años, y el análisis se hizo de manera independiente para cada paisaje. Para evaluar el efecto a escala de los predios con acuerdos de conservación, se usó una aproximación similar, esta vez con una zona de amortiguamiento para cada predio, con el fin de comparar de manera pareada las tasas de deforestación. En este segundo análisis, las réplicas fueron los predios con sus respectivas zonas de amortiguamiento; se hizo una prueba estadística para cada año y cada paisaje comparando de forma pareada los valores de deforestación en predios y las zonas de amortiguamiento (2015-2018). Las pruebas de Wilcoxon se realizaron con el paquete exactRankTests de R (Hothorn & Hornik, 2019).
Resultados
Las estrategias implementadas, derivadas de la planificación participativa en cada paisaje, incluyeron acuerdos de conservación con propietarios (no cacería, protección de hábitat, restauración), generación de corredores biológicos, implementación de proyectos productivos sostenibles como incentivos y fortalecimiento comunitario. En este trabajo pudimos evaluar de manera cuantitativa el efecto de los acuerdos de conservación a escala de predio en algunas especies de vertebrados. Con base en la información obtenida de las trampas cámara y los transectos, fue posible analizar las siguientes especies en el paisaje del Magdalena Medio: paujil de pico azul (C. alberti), choibo (A. hybridus), paca (Cuniculuspaca), guatín (Dasyproctapunctata) y pecarí de collar (Pecari tajacu). Las dos primeras son especies paisaje del proyecto, las otras tres, ejemplos de otras especies que se verían favorecidas por las acciones dirigidas a las primeras. En el paisaje de Orinoquia obtuvimos información para la danta de tierras bajas (T. terrestres), especie paisaje, y otras cinco especies de vertebrados terrestres (Tabla 1).
Tabla 1 Resultado de los modelos ajustados para cada especie en el paisaje del Magdalena Medio
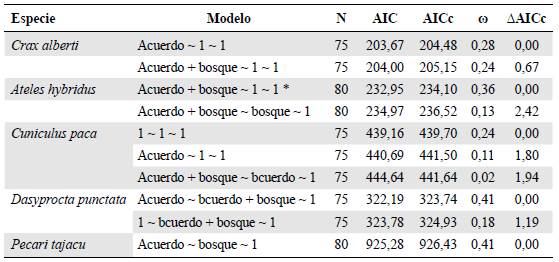
*El modelo no converge.
Para cada modelo se describen las variables usadas para explicar la probabilidad de colonización, la probabilidad de extinción, y la probabilidad de detección, respectivamente. Se muestran los modelos con un AAIC menor a dos. (1: variable constante. Bosque: área de bosque en el 2017). La probabilidad de detección es constante (1) en todos los modelos, pues no se encontró un efecto de ninguna covariable.
El área bajo acuerdos de conservación en el Magdalena Medio fue una variable explicativa de la probabilidad de colonización para C. alberti y D. punctata, apareciendo en los modelos con mejor ajuste y teniendo un efecto positivo y significativo (Tabla 1,figura 1).
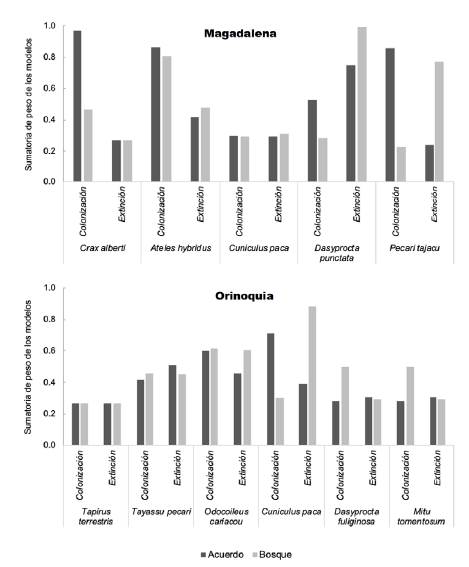
Figura 1 Peso cumulativo de las dos covariables (área bajo acuerdo de conservación en la celda y área de bosque en la celda) en los parámetros de colonización y extinción en los modelos evaluados
Para A. hybridus esta variable no fue significativa, sin embargo, apareció en los dos mejores modelos y presentó el mayor peso acumulado (Tabla 1,figura 1). Para C. paca no hubo un efecto de las covariables en la probabilidad de colonización, siendo el mejor modelo el nulo (Tabla 1). Por otro lado, para el pecarí de collar (P tajacu), el área de bosque tuvo un efecto significativo en la probabilidad de extinción, pues a mayor área de bosque menor la probabilidad de extinción (Tabla 1,figura 1).
En el caso del paisaje de Orinoquia, para C. paca se evidenció un efecto positivo y significativo del área de acuerdo en la colonización (Tabla 2,figura 1); para el venado (Odocoileus cariacou) se observó un efecto significativo del área de bosque en la colonización y en la extinción, respectivamente (Tabla 2,figura 1). Para las demás especies de este paisaje no hubo un efecto de las covariables en las probabilidades de colonización o extinción, siendo el nulo el mejor modelo en todos los casos (Tabla 2).
Tabla 2 Resultado de los modelos ajustados para cada especie en el paisaje de la Orinoquia
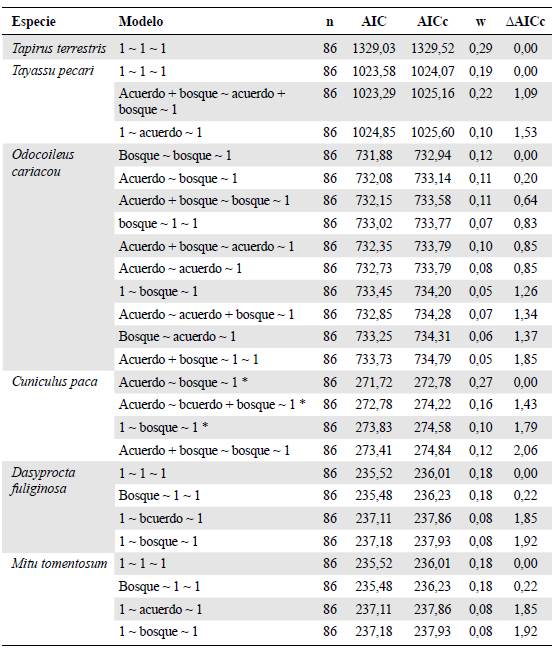
Para cada modelo se describen las variables usadas para explicar la probabilidad de colonización, la probabilidad de extinción, y la probabilidad de detección, respectivamente. Se muestran los modelos con un AAIC menor a dos. (1: variable constante. Bosque: área de bosque en el 2017). La probabilidad de detección es constante (1) en todos los modelos, pues no se encontró efecto de ninguna covariable.
Al comparar el efecto de las variables (área de acuerdo y área de bosque) en los dos paisajes, observamos que el área de acuerdo tuvo un efecto importante en las especies de Magdalena Medio, ya que los pesos acumulados de los modelos con esta variable fueron altos y mayores que para el área de bosque (Figura 1), y este efecto fue positivo, pues a mayor área protegida por un acuerdo, mayor la probabilidad de colonización (Figura 2). En el paisaje de Orinoquia los pesos acumulados fueron más bajos, denotando el poco poder explicativo de las covariables en la colonización o extinción en este paisaje, excepto para C. paca y O. cariacou (Figura 1). Aquí el área total de bosque en el 2017 tuvo un efecto negativo sobre la probabilidad de extinción, pues a mayor área de bosque, menor la probabilidad de extinción en una celda dada (Figura 3).
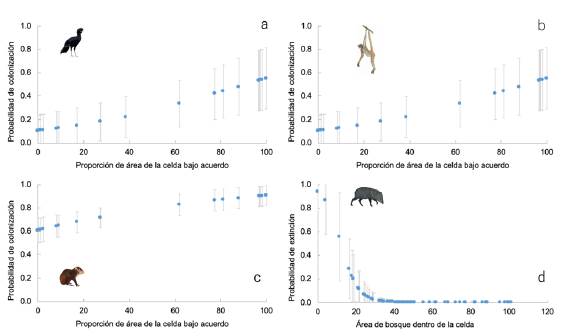
Figura 2 Relación entre las probabilidades de colonización o extinción en un cuadrante de lxl km2 entre el 2015 y el 2017 y las distintas variables explicativas: proporción del área bajo acuerdos de conservación y área de bosque en el cuadrante. Especies del paisaje del Magdalena Medio: a) Crax alberti, b) Ateles hybridus, c) Dasyprocta punctata y d) Pecari tajacu
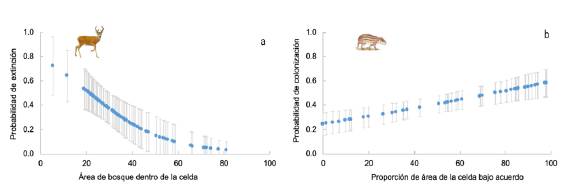
Figura 3 Relación entre las probabilidades de colonización o extinción en un cuadrante de 1x1 km2 entre el 2015 y el 2017 y las distintas variables explicativas, proporción del área bajo acuerdos de conservación y área de bosque en el cuadrante. a) O. cariacou, b) C. paca
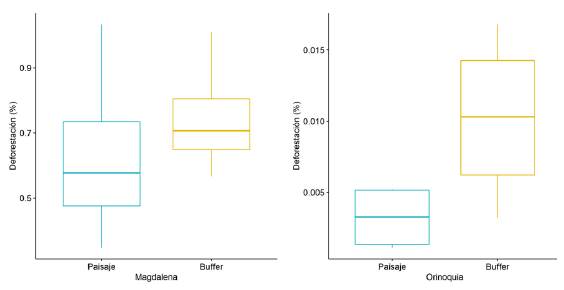
En cuanto a la segunda hipótesis evaluada, a escala de paisaje la deforestación promedio en los cuatro años fue mayor en el área de amortiguamiento que dentro del paisaje, tanto en el Magdalena Medio como en la Orinoquía (Figura 4), sin embargo, esta diferencia no fue significativa según la prueba estadística pareada no paramétrica (V=1, p=0,25; V=0, p=0,125, n=4). Es importante señalar que el número de réplicas (años) no tuvo un poder estadístico suficiente para detectar un efecto. A escala de predio (acuerdos de conservación) se encontró un efecto significativo en la reducción de la deforestación únicamente para el 2017 (V=66, p=0,033, n=46) en el paisaje de Magdalena. En el paisaje de Orinoquia los valores de deforestación fueron demasiado bajos y la mayoría de predios no presentó pérdida de bosque, por lo que no fue posible realizar la prueba estadística a escala de predio.
Discusión
Se presentó aquí un método para abordar dos hipótesis de la aproximación de conservación de especies paisaje mediante la evaluación del impacto de acciones de conservación en dichas especies y otras, y en la deforestación (como medida del mantenimiento de la estructura). En el ejemplo utilizado se encontró que las intervenciones basadas en la planificación concertada, específicamente los acuerdos de conservación, tuvieron un impacto positivo en algunas de las especies paisaje. En el Magdalena Medio, las dos especies paisaje evaluadas (paujil y choibo) se vieron favorecidas por las intervenciones en los predios con acuerdos. El éxito de los acuerdos sobre la ocupación de estas especies se debió seguramente a una combinación de reducción en la cacería y la protección y restauración de su hábitat. Estas dos especies dependen de bosques en buen estado y son muy sensibles a la cacería. En el período de tiempo evaluado pasaron a ocupar preferentemente sitios con acuerdos dentro del paisaje, lo que indica que la estrategia funcionó. En la Orinoquia, la especie paisaje T. terrestris no se vio influenciada positivamente por los acuerdos de conservación. La proporción del área ocupada por la danta en la cuenca del rio Bita ha sido alta (>80 %) desde el inicio del proyecto, lo que sugiere que sus poblaciones se encuentran en buen estado. En este sentido, las acciones de conservación se han orientado en mantener dichas poblaciones y no a recuperarlas. Aunque no encontramos evidencia de que ocupen preferencialmente sitios con acuerdos de conservación, las bajas tasas de extinción en el paisaje (<0,1 %), indican que la especie persiste. Es posible que a escala del paisaje la apropiación que ha generado la especie como símbolo de conservación esté contribuyendo a su mantenimiento en todos los predios, no solo en aquellos con acuerdos. Para determinar el efecto del proyecto en la danta a escala de paisaje, sería necesario comparar el estado de la especie en paisajes similares donde no se esté realizando una iniciativa de conservación.
Nuestra aproximación permitió evidenciar que otras especies de vertebrados (aparte de las especies paisaje) también se vieron favorecidas por las acciones de conservación. En el Magdalena Medio los guatines (D. punctata) y en la Orinoquia las pacas (C. paca) aumentaron su probabilidad de colonización en áreas de acuerdos. Estas especies, tradicionalmente objeto de cacería en estas regiones del país, se vieron favorecidas por la protección conferida por los predios con acuerdos. Por otro lado, otras especies evaluadas (no especies paisaje), especialmente en el paisaje de Orinoquia, no se vieron favorecidas por los acuerdos de conservación. Las tasas de colonización o extinción de varias de ellas no se correlacionaron con las variables evaluadas, lo que indica que los acuerdos de conservación han favorecido claramente a algunas especies, pero no a todas.
El pecarí en el Magdalena y el venado en la Orinoquia mostraron verse afectados también por el área de bosque. Su probabilidad de extinción disminuye a medida que el área de bosque aumenta. Este es un factor ecológico que se ha reportado como importante para la ocupación de algunos vertebrados tropicales (Boron, et al., 2019). Si bien estas especies no fueron favorecidas por los acuerdos directamente, el hecho de que el área de bosque sea importante para ellas nos muestra que pueden verse favorecidas por las intervenciones, ya que es posible proteger y restaurar dichos bosques.
Con base en estos resultados, podemos decir que la hipótesis de la conservación de especies paisaje se cumplió en la iniciativa evaluada con algunas salvedades. La acción de conservación (acuerdos de conservación) sí favoreció a las especies paisaje y a algunas otras de vertebrados. Sin embargo, no hay evidencia de que los acuerdos a escala de predio, en particular, estén incidiendo de forma directa en el mantenimiento de las poblaciones de varias especies (no paisaje) evaluadas (Mitu tomentosum, Odocoileus cariacou, Dasyprocta fuliginosa). No obstante, es importante señalar que los acuerdos no son la única estrategia implementada en el proyecto y que existen otras acciones centradas en las comunidades (educación ambiental, proyectos económicos asociativos, etc.), que pueden tener efectos difusos en todo el paisaje de intervención. Sería necesario evaluar si estas últimas especies están siendo protegidas, no a escala de los predios con acuerdos, sino a escala de todo el paisaje. La ocupación de algunas de ellas se mantuvo o aumentó en el paisaje durante el período de trabajo. Este es un buen indicio de éxito, aunque no permite demostrar causalidad, como se hizo con la actividad específica de los acuerdos de conservación. Además, es posible que el tiempo de las intervenciones no haya sido suficiente para generar un impacto sobre ciertas especies. Se ha observado que existen retrasos temporales en la respuesta de las especies a las acciones de conservación, los cuales pueden enmascarar el avance en los objetivos (Watts, et al., 2020). El marco metodológico presentado aquí permitirá evaluar en el tiempo si están ocurriendo este tipo de retrasos.
La segunda hipótesis evaluada fue si las acciones de conservación tenían un efecto en la tasa de deforestación como medida indirecta del mantenimiento de la estructura. Los resultados a escala de paisaje mostraron que, si bien la deforestación fue mayor en el área de amortiguamiento a lo largo del periodo estudiado, los datos no permiten aún detectar una tendencia significativa. En el Magdalena Medio, en tres de los cuatro años la pérdida de bosque fue mayor por fuera del paisaje, y en el paisaje de Orinoquía fue mayor en todos los años. Es un caso de réplicas insuficientes aún y requerirá acumulación de evidencia. Los resultados a escala de predio contaban con más replicas e indicaron que solamente en el 2017 las intervenciones tuvieron un efecto significativo en el Magdalena Medio en cuanto a la disminución de la pérdida de bosque en comparación con áreas sin acuerdos en el mismo paisaje. Reducir la deforestación es una de las metas más complejas de lograr en áreas rurales del país. Los acuerdos de conservación tienen el potencial de contribuir con esta meta, como quedó demostrado en este estudio, pero requieren un decidido acompañamiento a los propietarios y una verificación continua. Esta contribución a la protección de los bosques, sumada a los resultados en las especies, confirman que los acuerdos de conservación en tierras privadas, como parte de las estrategias complementarias de conservación (Other effective area-based conservation measures, OECM), tienen el potencial de contribuir al mantenimiento de la biodiversidad y el cumplimiento de metas nacionales y globales (metas AICHI) de desarrollo sostenible (Dudley, et al., 2018; Hardy, et al., 2017).
La aproximación presentada en este estudio permitió evaluar de forma cuantitativa los efectos de un programa de conservación. De manera general, la disciplina de la conservación ha sido lenta en la adopción de métodos de evaluación de impacto que permitan determinar el efecto de las medidas o programas de conservación implementados (Ferraro & Pattanayak, 2006; Baylis, et al, 2016). Sin embargo, la evidencia sobre cuáles prácticas son exitosas y cuáles no es indispensable para diseñar programas costo-efectivos y hacer uso eficiente de los recursos limitados para conservación. Algunos retos de implementar evaluaciones de impacto rigurosas en programas de conservación incluyen la búsqueda de objetivos múltiples en múltiples escalas, los factores de desviación (confounding factors), la limitada posibilidad de aleatorizar y replicar, y el poco tiempo de ejecución en la mayoría de las iniciativas (Ferraro & Pattanayak, 2006). Sin embargo, es necesario evitar recolectar solamente indicadores descriptivos y enfocarse en la pregunta fundamental de una evaluación de impacto: ¿qué habría sucedido si la intervención no se hubiera realizado? Esto requiere plantear hipótesis claras y evaluables, e implementar diseños que permitan identificar relaciones de causalidad, aislando el efecto de la intervención de otros posibles factores que generen sesgos (Ferraro & Pattanayak, 2006). Es necesario también contar con una ética de responsabilidad y reporte, pues muchas veces se encontrará que la intervención, cuando se aísla de los factores de sesgo, no es la causa real del cambio observado (Ferraro & Pattanayak, 2006).
Con la presentación de esta aproximación buscamos aplicar principios de la disciplina de evaluación de impacto para estudiar el efecto de las acciones de conservación de especies paisaje y probar dos hipótesis en ese modelo. En los dos casos (efecto sobre especies y deforestación) generamos hipótesis evaluables, usamos información sobre la línea de base, los grupos de control (celdas sin acuerdos, áreas de amortiguamiento sin intervención) y evaluación del efecto de las covariables. El análisis de ocupación multitemporada se basó en muestreos en celdas con acuerdos y sin acuerdos en todo el paisaje a lo largo de un periodo de dos años. El análisis de deforestación se basó en la comparación de dichas tasas en áreas equivalentes, reduciendo así efectos de covariables asociadas al riesgo de deforestación (Armenteras, et al., 2009). La medición de la reducción de la deforestación en áreas protegidas ha sido uno de los campos donde más ha avanzado la evaluación de impacto en la conservación (Joppa & Pfaff, 2010; Armenteras, et al., 2009;Negret, et al., 2020), pero generalmente ha estado enfocada en áreas protegidas, con polígonos relativamente bien definidos, mientras que ha tenido menos desarrollo en iniciativas de conservación por fuera de áreas protegidas, donde la escala espacial del impacto es más difusa. Sugerimos que estos métodos cuantitativos sean implementados también en estos casos, pues permiten entender los efectos en las tasas de pérdida de bosque causadas por el programa y evaluar la escala de dichos impactos.
Para el análisis de ocupación evaluamos solamente un tipo de intervención, los acuerdos de conservación, a una escala específica, el predio. Sin embargo, hay acciones en todo el paisaje y un trabajo social extenso. Es posible que el efecto en algunas especies no se esté dando asociado únicamente con los acuerdos, limitados a un predio, sino en todo el paisaje. Para evaluar esto será necesario hacer un ejercicio similar comparando la ocupación con áreas que no tengan influencia del proyecto.
Los métodos utilizados representan un ejemplo del uso de enfoques cuantitativos para evaluar los impactos de un programa de conservación. Fue posible observar que la aproximación de conservación de especies paisaje implementada a escala del paisaje tiene el potencial de impactar positivamente a las especies focales y a otros vertebrados. Sin embargo, no todas las especies se ven afectadas de la misma forma por ciertas intervenciones. Las variaciones en la respuesta a las intervenciones también se evidencian entre paisajes. Además, las diferencias entre las historias de vida de las especies y el tiempo de evaluación podrían hacer que algunas especies evidencien efectos positivos, en tanto que otras aún no. Por esto, es necesario usar diseños basados en principios de evaluación de impacto para medir y entender el éxito de las intervenciones en el marco de un proyecto. Solo un reporte riguroso y transparente permitirá avanzar en el ejercicio de la conservación.

