











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
El bosque seco tropical es uno de los hábitats más amenazados en el mundo por actividades antropogénicas como los incendios forestales y, con menor frecuencia, por eventos naturales como las erupciones volcánicas (Bocanegra-González, et al., 2018). El fuego afecta los procesos biológicos y biogeoquímicos de los ecosistemas (Verma & Jayakumar, 2015, 2018) modificando las propiedades fisicoquímicas del suelo (Verma & Jayakumar, 2012), la diversidad de las comunidades vegetales y de microorganismos como los hongos totales, saprofitos y hongos micorrízicos arbusculares (HMA), contándose estos últimos entre los más susceptibles (Longo, et al., 2014; Cofré, et al., 2018; Day, et al., 2019), especialmente en las estructuras miceliales y en la colonización micorrízica in situ; por el contrario, por ser estructuras de resistencia, las esporas no se ven drásticamente afectadas (Xiang, et al., 2015; Sun, et al., 2015; Holden, et al., 2016), aunque algunas veces a cierta profundidad, el calor producido por el fuego rompe su latencia (Claridge, et al, 2009).
Aunque los eventos naturales como las erupciones volcánicas no son frecuentes, al estudiar los HMA en suelos afectados por este fenómeno se ha encontrado que su riqueza y diversidad disminuyen, pero aumentan de nuevo al pasar los años (Gemma & Koske, 1990) mediante el establecimiento inicial de taxones generalistas y tolerantes con capacidades de adaptación y colonización que pueden tener implicaciones en la resiliencia de la vegetación después de una perturbación volcánica (Wu, et al., 2007; Aguilera, et al., 2018; Borie, et al., 2019; Atunnisa & Ezawa, 2019).
Los HMA pertenecen al filo Glomeromycota y hacen parte de los microorganismos ecológicamente más importantes del planeta (van der Heyde, et al., 2017), debido a que establecen simbiosis con el 72 % de las plantas vasculares, aumentando en estas la absorción de agua y de nutrientes como el fósforo y nitrógeno a cambio de los carbohidratos y lípidos asimilados por la planta (Keymer & Gutiahr, 2018; Lanfranco, et al., 2018). Los HMA tienen un papel clave en los ecosistemas terrestres, pues regulan los ciclos de nutrientes (Smith, et al., 2011), influyen en la estructura y multifuncionalidad del suelo (van der Heyde, et al., 2009), establecen plántulas, descomponen la hojarasca, y forman y agregan el suelo (Rillig & Mummey, 2006; Lindahl, et al., 2007). Ademas, confieren a las plantas tolerancia a la sequía, los metales pesados, las enfermedades, y los patógenos, entre otros factores (Auge, 2001; Girma, 2015; Zhang, et al., 2015; Garcia, et al., 2016; Brundrett & Tedersoo, 2018).
A pesar de que el fuego y las erupciones volcánicas reducen la diversidad de morfoespecies de HMA comparada con la de las áreas naturales preservadas (de Assis, et al., 2018), si las condiciones climáticas, edáficas y de vegetación son favorables (Orgiazzi, et al., 2016; Whitman, et al., 2019), las comunidades pueden restablecerse en el curso de unos decenios, lo que contribuye a una rápida recuperación del ecosistema después de una perturbación de ese tipo (Dove & Hart, 2017; Saini & Sharma, 2019).
En Colombia los estudios sobre los HMA se han centrado principalmente en cultivos de frutas, hortalizas y legumbres (Gómez & Sánchez de Prager, 2012; Guzmán, et al., 2013; Peña-Venegas, 2015; Rodríguez & Sanders, 2015; Montenegro-Goméz, et al., 2017; Ramírez-Gómez, et al., 2019; Sandoval-Pineda, et al., 2020) como promotores del crecimiento y el rendimiento vegetal (Castillo, et al., 2012; Hernández-Ortega, et al., 2012) o por su efecto en la tolerancia de las plantas frente a factores fisicoquímicos (Aguilera, et al., 2014; Seguel, et al., 2016).
Los estudios sobre la asociación de las micorrizas en ecosistemas naturales (Peña-Venegas, et al., 2019) y el inventario de HMA en el país son escasos, principalmente en ecosistemas de bosque seco tropical con perturbaciones por el fuego y los depósitos fluviovolcánicos, los cuales pueden generar gran variabilidad en la respuesta microbiana debido al elevado número de factores que se ven alterados y que repercuten en los microorganismos. En este contexto surgió la pregunta sobre qué incidencia han tenido el fuego y los depósitos fluviovolcánicos en las comunidades de HMA del bosque seco tropical del departamento del Tolima. En este sentido, se plantea que en las áreas perturbadas por fenómenos antrópicos y naturales, la composición de los hongos micorrízicos está dominada por microorganismos tolerantes y que la abundancia y diversidad es menor que las de las áreas naturales. Por este motivo, el presente estudio se propuso evaluar el efecto del fuego y los depósitos fluviovolcánicos en la composición, la abundancia y la diversidad de los HMA en suelos de bosque seco tropical del departamento del Tolima, Colombia.
Materiales y métodos
Área de estudio y maestreo
El estudio se desarrolló en cuatro áreas del departamento del Tolima: dos se encuentran en el municipio de Ibagué, en la Reserva Ecológica Santafé de Los Guaduales (4°24'17,84" N & 75°11'46,02" O), a una distancia de 284 m entre ellas; otra, con condiciones de bosque nativo (RN), y la cuarta afectada por el fuego de un incendio forestal (RQ) ocurrido dos años antes del muestreo. La zona se encuentra a 1.090 m s.n.m. y presenta una temperatura promedio de 21,1 °C. En el área RN domina la vegetación leñosa y en la RQ las herbáceas de la familia Poaceae del género Brachiaria.
Las otras dos áreas están ubicadas en el municipio de Armero (4°55'52,5" N & 74°52'36,3" O), a una distancia de 4.881,2 m entre ellas, una con bosque nativo (AN) y otra afectada por depósitos fluviovolcánicos provenientes de la erupción del volcán Nevado del Ruiz en 1985 (ADV) (Figura 1). La zona se encuentra a 352 m s.n.m. y presenta una temperatura promedio de 34,1 °C. La cobertura vegetal de AN está dominada principalmente por las familias Fabaceae, Poaceae, Asteraceae, Malvaceae y Euphorbiaceae (Esquivel, et al., 2016), en tanto que la ADV es un área empleada en actividades agrícolas.

En puntos aleatorios de cada área se tomaron cuatro muestras compuestas de suelo desde el horizonte A, a una profundidad de 0 a 20 cm, con un barreno holandés. Las muestras se almacenaron a 4 °C hasta el momento de su análisis.
Multiplicación de HMA
Se hicieron cultivos trampa utilizando la metodología de Morton, et al. (1995), con el fin de detectar aquellas morfoespecies de HMA que no esporulaban en campo en el momento del muestreo (Posada, et al., 2018). Los cultivos se establecieron a partir de la siembra de plántulas de sorgo (Sorghum bicolor) en macetas que tenían como sustrato una mezcla de suelo y arena en proporción 2:1 (v/v), colocadas en autoclaves durante una hora por 3 días a 121 °C y 15 lb/pulg2. Posteriormente, el suelo extraído en campo se utilizó como inóculo. El sorgo es una planta de la familia Poaceae de rápida germinación y altamente micotrófica, por lo que se emplea como planta huésped para la multiplicación de estos hongos (Sánchez de Prager, et al., 2010).
Los cultivos se mantuvieron en el invernadero del Grupo de Genética y Biotecnología Vegetal y Microbiana de la Universidad del Tolima (GEBIUT) durante tres meses, se regaron a capacidad de campo cada 48 horas y se fertilizaron cada 30 días empleando la solución nutritiva de Hoagland & Amon (1950). Después de 120 días, los cultivos se sometieron a estrés hídrico y mecánico para estimular la producción de esporas.
Extracción e identificación morfológica de esporas de HMA
Mediante el método de tamización en húmedo y centrifugación en gradiente de sacarosa (Gerdemann & Nicolson, 1963), se extrajeron las esporas de HMA a partir de 10 g de las muestras directas de suelo (SD) y de los cultivos trampa (CT). Las esporas se cuantificaron y separaron en placas de Petri de 80 mm con cuadriculas de 1 cm bajo un microscopio estereoscópico (Brundrett, et al., 1994). Posteriormente, se agruparon y montaron en portaobjetos de vidrio con polivinil lactoglicerol (PVLG) y PVLG + reactivo de Melzer (1:1 v/v) (Morton, 1988). Estas se observaron a 40 x y 100 x en un microscopio compuesto Primo Star y se fotografiaron usando una cámara acoplada Axiocam ERc 5s de cinco megapíxeles en el programa Zen.
Con base en las características morfológicas de las esporas; color, diámetro, número de paredes, tamaño, forma de la conexión hifal y tipo de ornamentaciones, se hizo la identificación taxonómica teniendo como referencia el manual de Pérez & Schenk (1990) y la información disponible en los sitios web International Culture Collection of Arbuscular & Vesicular-Arbuscular Mycorrhizal Fungi (http://fungi.invam.wvu.edu/the-fungi/species-descriptions.html), así como, la del profesor Janusz Blaszkowski (http://www.zor.zut.edu.pl/Glomeromycota/index.html). Además, se tuvieron en cuenta publicaciones científicas que dan soporte para la clasificación de género y morfoespecies (Blaszkowski & Chwat, 2013; Sieverding, et al., 2014; Pereira, et al., 2015) y el apoyo de personal del laboratorio de microbiología agrícola de la Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA).
Análisis del suelo
Se analizó el pH en suspensión de suelo y agua en proporción de 1:2,5 (p/v), la conductividad eléctrica (CE) según la norma colombiana NTC 5596:2008, la materia orgánica (MO) por el método de Walkley & Black, el fósforo disponible (P) por el método de Bray II y el calcio (Ca2+), el magnesio (Mg2+), el potasio (K+) y el sodio (Na+) intercambiable por acetato de amonio (CH3COONH41M pH 7,0). Por último, la capacidad de intercambio catiónico (CICE) y la saturación de Ca2+, Mg2+, K+, y Na+ se determinaron por cálculo. Las muestras fueron analizadas en el Laboratorio de Química Analítica del Centro de Investigación Tibaitatá de AGROSAVIA en Mosquera, Cundinamarca. Las variables químicas fueron evaluadas por área de muestreo.
Análisis ecológico
Se calculó la composición de los HMA correspondiente a las morfoespecies que componen cada una de las áreas y la abundancia de esporas a partir del número total de esporas en 10 g-1 de suelo, la abundancia relativa de cada morfoespecie y la diversidad a partir de los números efectivos de Hill. Las series de Hill incluyen la diversidad de orden, q=0, que equivale a la riqueza de morfoespecies; q=1, que se basa en el exponente de Shannon-Wiener (H') y corresponde al número de morfoespecies raras, y el orden q=2 a partir del inverso de Simpson (D'), que determina el número de morfoespecies muy abundantes o dominantes (Hill, 1973). Asimismo, a partir de las muestras de suelo extraídas directamente en campo (SD), se identificaron las morfoespecies "indicadoras" correspondientes a aquellas con alta especificidad y fidelidad a un área determinada (Dufrêne & Legendre, 1997).
Análisis estadístico
Para evaluar los cambios en la composición de la comunidad de HMA en los sitios de muestreo, se empleó un análisis de escalamiento no métrico multidimensional (ENMM) y un análisis de similitud (ANOSIM) utilizando la distancia de Bray Curtis. Las diferencias en la abundancia de esporas de HMA se analizaron mediante análisis de varianza (ANAVA) con pruebas de normalidad (Shapiro-Wilks), homogeneidad de la varianza (prueba de Levene) y comparación de Tukey (p≤0,05). Sin embargo, no se cumplieron los supuestos de normalidad, por lo cual se recurrió a una Kruskal Wallis con prueba post hoc por pares.
Para determinar diferencias en la diversidad de los HMA, se hizo una extrapolación y una interpolación utilizando los números de Hill (Chao, et al., 2014). Este análisis permitió establecer en qué suelos se registró una mayor diversidad de HMA. Con el fin de determinar las morfoespecies indicadoras en cada una de las áreas de SD, se utilizó el análisis indicador de especies propuesto por Dufrêne & Legendre (1997). Se consideraron los taxones con un valor indicador (VI) ≥ 80 % y p≤0,05, como lo sugieren Tejeda-Cruz, et al. (2008). Por último, se realizó un análisis canónico de correspondencias (ACC) a partir de la transformación de las abundancias (raíz cuadrada) para inferir la relación entre las comunidades de HMA y las características químicas del suelo. Todos los análisis estadísticos se hicieron en el programa estadístico "R" versión 3.4.1 (R Core Team, 2020).
Resultados
Composición de la comunidad de HMA
El análisis ENMM reveló que la composición de los HMA difirió significativamente entre las áreas de muestreo (Figura 2), lo que se confirmó en el ANOSIM en las muestras de SD (R=0,9828, p= 0,001) y de CT (R=0,937, p=0,001). Sin embargo, a pesar de que cada área evaluada exhibió su propia composición de HMA, se logró detectar que las áreas perturbadas desarrollaron una comunidad característica, en tanto que las naturales presentaron morfoespecies similares entre ellas.
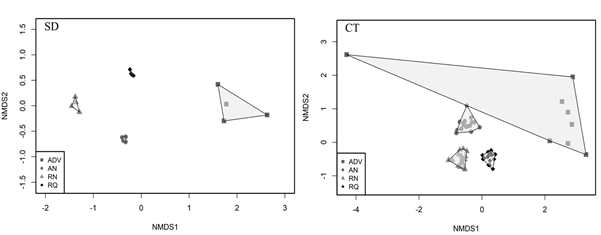
Figura 2 Diagrama de ordenación ENMM de la composición de HMA en las áreas de bosque seco tropical del departamento del Tolima
Se registraron 64 morfoespecies pertenecientes a cuatro órdenes, nueve familias y 17 géneros en todas las áreas de muestreo desde SD y CT, con Glomus y Acaulospora como los géneros con más representantes (Tabla 1). En las áreas de la Reserva se recolectaron 50 morfoespecies, de las cuales 27 se encontraron en RQ y SD, 33 en RQ y CT, 15 en RN y SD y 19 en RN y CT. Por otra parte, Armero presentó 34 morfoespecies: 14 en AN y SD, 23 en AN y CT, 9 en ADV y SD y 19 en ADV y CT.
| Abundancia relativa % | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Orden | Familia | Morfoespecies de HMA | RN | RQ | AN | ADV | ||||
| SD | CT | SD | CT | SD | CT | SD | CT | |||
| Glomerales | Glomeraceae | Funneliformis mosseae | 2,7 | 3,2 | 4,8 | 12,3 | 45,4 | |||
| Funneliformis geosporum | 2,1 | 2,7 | 1,9 | |||||||
| Glomus microcarpum | 10,4 | 11,4 | 4,3 | 6,7 | 71,3 | 29,7 | 3,4 | 0,4 | ||
| Glomus atrouva | 4,2 | 6,3 | 5,7 | 0,8 | 2,1 | 0,3 | ||||
| Glomus badium | 2,1 | 6,4 | 8,6 | 7,2 | 2 | 8,3 | 1,1 | |||
| Glomus coremioides | 2,6 | 0,7 | ||||||||
| Glomus albidum | 9,4 | 4,6 | 3,4 | 0,2 | ||||||
| Glomus aureum | 7,5 | 10,6 | 6,7 | 2 | 5,5 | 1,3 | 0,2 | |||
| Glomus caledonium | 4,2 | 5,8 | 0,2 | |||||||
| Glomus clavisporum | 2 | 1,2 | ||||||||
| Glomus heterosporum | 0,6 | |||||||||
| Glomus gibbosum | 1,9 | 2,4 | ||||||||
| Glomus glomerulatum | 0,9 | |||||||||
| Glomus tenerum | 0,4 | |||||||||
| Glomus trufemi | 0,6 | |||||||||
| Glomus coronatum | 0,5 | |||||||||
| Glomus hyderabadensis | 0,5 | |||||||||
| Glomus sp | 13,5 | 4,6 | ||||||||
| Glomus sp1 | 4,2 | 1,5 | ||||||||
| Rhizoglomus fasciculatum | 6,2 | 5,3 | 7,1 | 1,9 | 9,1 | 1,3 | ||||
| Rhizoglomus agregatum | 12,1 | 5,5 | 3 | 5,6 | 4,8 | |||||
| Rhizoglomus sp | 1 | |||||||||
| Rhizoglomus sp1 | 1,9 | |||||||||
| Rhizoglomus intraradices | 1,3 | |||||||||
| Rhizoglomus manihotis | 5,2 | 3,5 | 0,9 | 2 | ||||||
| Sclerocystis pachycaulis | 0,9 | |||||||||
| Sclerocystis sinuosum | 3,8 | 6,6 | 4,7 | 2,7 | 2,7 | |||||
| Sclerocystis rubiforme | 5,8 | 8,6 | ||||||||
| Simiglomus hoi | 1,1 | |||||||||
| Septoglomus constrictum | 2,1 | 1,2 | 1 | |||||||
| Septoglomus desertícola | 3,8 | 0,6 | 2,2 | 0,4 | ||||||
| Entrophosporaceae | Entrophospora sp | 0,3 | ||||||||
| Entrophospora báltica | 10,3 | 8,4 | 7,1 | 1,3 | ||||||
| Claroideoglomus sp | 2,4 | 0,9 | 0,6 | |||||||
| Claroideoglomus sp1 | 3,5 | |||||||||
| Claroideoglomus luteum | 1,4 | |||||||||
| Claroideoglomus lamellosum | 0,7 | |||||||||
| Claroideoglomus etunicatum | 3,1 | |||||||||
| Diversisporales | Acaulosporaceae | Acaulospora sp | 1,3 | |||||||
| Acaulospora sp1 | 2,2 | 0,3 | ||||||||
| Acaulospora morrowiae | 1,4 | |||||||||
| Acaulospora spinosa | 6,4 | |||||||||
| Acaulospora mellea | 1,4 | 1,1 | 1,9 | 3,5 | 7,5 | |||||
| Acaulospora foveata | 0,6 | 2,8 | 3,4 | |||||||
| Acaulospora reducta | 22,9 | 5,5 | 3,4 | 6,4 | 4 | |||||
| Acaulospora colombiana | 2,2 | 1,4 | 0,6 | |||||||
| Diversisporaceae | Diversispora epigaea | 4,9 | 0,8 | 0,6 | ||||||
| Diversispora trimurales | 7,1 | |||||||||
| Diversispora aurantia | 1,7 | |||||||||
| Diversispora clara | 1 | |||||||||
| Diversispora sp | 3 | 36,6 | ||||||||
| Diversispora sp1 | 3,1 | |||||||||
| Redeckera sp | 3 | 0,4 | ||||||||
| Redeckera sp1 | 3 | |||||||||
| Redeckera sp2 | 1,7 | 1,3 | ||||||||
| Pacisporaceae | Pacispora sp | 0,2 | ||||||||
| Pacispora sp1 | 0,8 | |||||||||
| Archaeosporales | Ambisporaceae | Ambispora sp | 0,6 | 0,2 | 71,5 | 0,2 | ||||
| Ambispora sp1 | 1,4 | |||||||||
| Gigasporales | Gigasporaceae | Gigaspora sp | 0,8 | |||||||
| Scutellosporaceae | Scutellospora sp | 1 | 0,7 | |||||||
| Racocetraceae | Racocetra sp | 0,4 | ||||||||
| Cetraspora pellucida | 0,3 | |||||||||
Abundancia de la comunidad de HMA en el bosque seco tropical
La abundancia presentó diferencias significativas entre las áreas de muestreo (p<0,05), siendo mayor en RQ, con un total de 899 esporas en 10 g-1 de suelo en SD y 1.867 esporas en 10 g-1 de suelo en los CT. Las áreas naturales (RN y AN) mostraron abundancias similares en SD; sin embargo, en los CT fueron diferentes entre sí, con la mayor abundancia en RN. De todas las áreas evaluadas, ADV presentó la menor abundancia tanto en SD como en CT (Figura 3). En los CT aumentó 2,1 veces el número de esporas en RQ y RN, 1,83 veces en AN y 4,4 en ADV
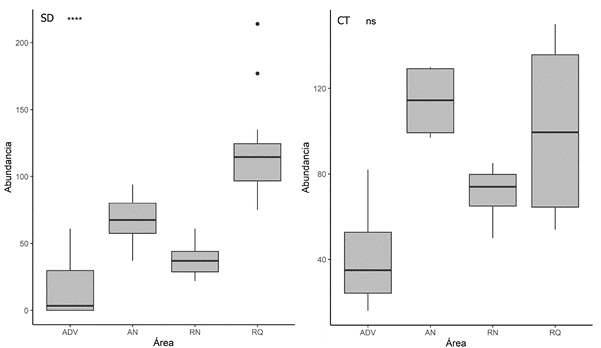
Figura 3 Abundancia de HMA en las áreas de estudio de bosque seco tropical del departamento del Tolima. ****= p<0,001; ns = no significativo
De todas las morfoespecies encontradas, las más abundantes fueron Ambispora sp. y Funneliformis mosseae en ADV en muestras de SD y CT, respectivamente, seguidas de Glomus microcarpum en AN-SD, AN-CT y RN-CT, Acaulospora reducta en RN-SD, Glomus aureum y Sclerocystis rubiforme en RQ en SD y CT, respectivamente (Tabla 1).
Diversidad de HMA en el bosque seco tropical
A partir de la extrapolación e interpolación del número efectivo de especies (q=0, q=1 y q=2) en SD y CT, se determinó que RQ concentró la mayor riqueza y diversidad de morfoespecies; en contraste, las áreas AN y ADV registraron el menor número de morfoespecies. Los intervalos de confianza obtenidos en el análisis permitieron evidenciar diferencias significativas de la diversidad de HMA en las áreas evaluadas (Figura 4).
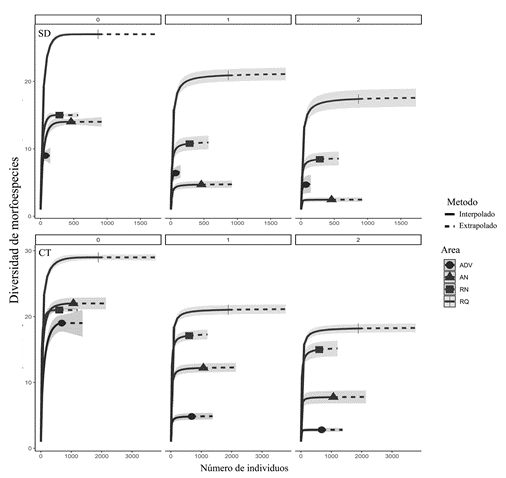
Morfoespecies de HMA indicadoras en bosque seco tropical
En la Reserva, RN presentó como morfoespecies indicadoras a Glomus sp. y Septoglomus desertícola; en RQ fueron Acaulospora sp, Claroideoglomus luteum, Claroideoglomus sp., Funneliformis geosporum, Glomus caledonium, Sclerocystis pachycaulis, S. rubiforme, Glomus atrouva, G. albidum y G. aureum. En la zona de Armero, las morfoespecies indicadoras de AN fueron G. microcarpum y Scutellospora sp., en tanto que en ADV fue Ambispora sp. (Tabla S1,https://www.raccefyn.co/index.php/raccefyn/article/view/1482/3154).
Análisis del suelo
Los resultados indican que tres de las cuatro áreas de estudio (RQ, RN y AN) presentaron un pH ácido y solo ADV exhibió un pH alcalino y un alto contenido de P, K+, Mg2+, CICE y saturación de Ca2+. RQ se caracterizó por presentar bajos contenidos de nutrientes con excepción del Na+. Las áreas naturales (RN y AN) mostraron parámetros químicos similares, en los que se destaca un elevado contenido de Mg2+ y CE. Por último, la materia orgánica fue baja en las áreas de la Reserva y alta en Armero, principalmente en ADV (Tabla S2,https://www.raccefyn.co/index.php/raccefyn/article/view/1482/3155).
Análisis canónico de correspondencias (ACC)
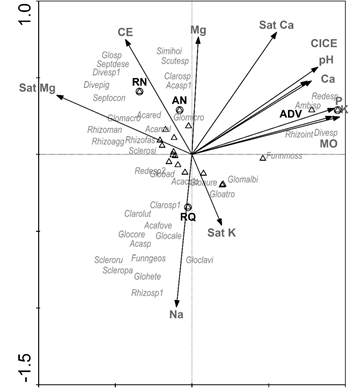
Ninguna de las variables químicas evaluadas influyó de manera significativa en la distribución de la comunidad de HMA. Sin embargo, en el diagrama de ordenación fue posible evidenciar una asociación positiva entre las variables de saturación de K y Na+ con las morfoespecies de las familias Glomeraceae y Acaulosporaceae. Además, fue posible identificar que las áreas naturales (RN y AN) exhibían una composición similar de la comunidad al compartir el 48 % de las morfoespecies y los altos niveles de Mg2+ y CE.
En ADV se registraron taxones como Ambispora sp., Funneliformis mosseae, Redeckera sp., Rhizoglomus intraradices y Diversispora sp., los cuales podrían tener la capacidad de desarrollarse en pH alcalino con altos contenidos de nutrientes, principalmente P y K+ (Figura 5).
Discusión
En general, los resultados del estudio sugieren que la composición, abundancia y diversidad de las comunidades de HMA difirieron significativamente, especialmente en suelos afectados por el fuego, seguidos de las áreas naturales y menor medida en el área de depósitos fluviovolcánicos.
Los resultados observados en RQ pueden atribuirse a la cobertura vegetal presente en esta área, dominada por gramíneas del género Brachiaria (Leal, et al., 2013), las cuales encuentran mejores oportunidades de crecimiento y establecimiento en suelos afectados por el fuego debido a la eliminación de la cubierta del dosel y un aumento en la entrada de luz al sotobosque (Coroi, et al., 2004). Se ha reportado que algunas especies de este género son huéspedes potenciales de HMA debido a su fácil establecimiento simbiótico y asociación con una alta producción de esporas (Sánchez de Prager, et al., 2010; Álvarez-Lopeztello, et al., 2019). En algunos trabajos se han reportado resultados similares, observándose un incremento en la abundancia de HMA después de experimentar incendios forestales, resultados que fueron atribuidos no al efecto del fuego y sí al aumento en la diversidad de herbáceas que se presentaron después del suceso (Dhillion, et al., 1988; Zhang, et al., 2004; Mirzaei, 2016; Dove & Hart, 2017; Carson, et al., 2019). Además, algunas esporas de HMA son termotolerantes y su germinación es estimulada por el estrés frente al fuego (Dhillion, et al., 1988; Purin & Rillig, 2007; Mirzaei, 2016; Ambrosino, et al., 2018), sin embargo, es una reacción que depende de la intensidad del mismo (Holden, et al., 2016; Verma & Jayakumar, 2018).
En el área de depósitos fluviovolcánicos, los menores valores observados pueden explicarse debido a que la erupción del volcán Nevado del Ruiz cubrió gran parte del suelo natural, modificando sus propiedades y eliminando la cobertura vegetal (Torres Benítez, 2013). A pesar de que en esta área se han iniciado nuevos procesos de formación de suelos, con una sucesión vegetal representada por 72 familias en las que se destacan Asteraceae, Malvaceae, Fabaceae, Euphorbiaceae y Poaceae (Esquivel, et al., 2016), la abundancia y la diversidad de HMA después de 33 años de ocurrido el evento sigue siendo escasa. Estos resultados pueden deberse a las características generadas por la exposición a las cenizas volcánicas (Satti, et al., 2007), o al efecto de la agricultura extensiva ejercida en la zona, la cual tiene un fuerte efecto negativo en la riqueza y diversidad de estos hongos debido al alto aporte de nutrientes producto de la fertilización que disminuye la dependencia simbiótica de las plantas y los HMA (Sieverding, 1991; Thirkell, et al., 2017; Hage-Ahmed, et al., 2019).
En las áreas naturales (RN y AN) se encontró que la composición y riqueza tiende a ser similar, pese a las diferencias significativas y a la localización en distintas zonas, lo que se explicaría por la similitud en las condiciones edafoclimáticas y de composición vegetal del bosque seco tropical conservado (Pizano & García, 2014). Estas áreas se caracterizaron por presentar las familias Scutellosporaceae, Gigasporaceae y Racocetraceae, las cuales se han reportado principalmente en zonas conservadas y se han catalogado como sensibles y menos capaces de adaptarse a las perturbaciones por el gran tamaño de sus esporas (>200 μm) (Morton & Benny, 1990), lo cual las hace más susceptibles al daño físico (Tchabi, et aL, 2008; Allen, et al., 2009; Turrini & Giovannetti, 2011; Chagnon, et al., 2013; van der Heyde, et al., 2017). De allí la importancia del cuidado de las áreas naturales, ya que son el reservorio de morfoespecies de HMA esenciales para el mantenimiento de los bosques nativos.
Una alta diversidad vegetal como la encontrada en las áreas naturales permite tener más morfoespecies raras de HMA debido a que existe un mayor número de huéspedes (Hontoria, et al., 2019; Davison, et al., 2020; Guzman, et al., 2021). Las morfoespecies raras o poco frecuentes son producto de la respuesta coevolutiva entre los HMA y las plantas para un mejor beneficio simbiótico; estas desarrollan capacidades funcionales según los requerimientos del huésped de su preferencia (van der Heijden, et al., 2003; Oliveira, et al., 2006; Helgason, et al., 2007). Por otro lado, la dominancia de algunas morfoespecies como las observadas en las áreas de depósitos fluviovolcánicos puede explicarse por la ausencia de distintos huéspedes y al establecimiento de hongos generalistas, los cuales tienen la capacidad de tolerar cambios o alteraciones en el ecosistema (Chagnon, et al., 2013) y de colonizar plantas en procesos de sucesión, generando competitividad e inhibiendo la incorporación de HMA de lenta germinación, sensibles y específicos (Helgason, et al., 2007).
El número de morfoespecies encontradas en este estudio fue mayor al registrado en otras investigaciones en bosque seco tropical (Chaudhary, et al., 2014; Balázs, et al., 2015; de Assis, et al., 2018; Carrillo-Saucedo, et al., 2018), con excepción de 80 morfoespecies registradas por Marinho, et al. (2019) en Brasil. Los principales géneros identificados en el presente trabajo fueron Glomus y Acaulospora, similar a lo reportado en otras zonas de bosque seco tropical del mundo (Holste, et al., 2016; de Assis, et al., 2018; Sousa, et al., 2018; Mahdhi, et al., 2019; Wang, et al., 2019; Rodrigues, et al., 2021) y de Colombia (García, et al., 2000; Herrera & Arana, 2010; Rodríguez-López, et al., 2015).
La familia Glomeraceae, con los géneros Glomus, Sclerocystis, Funneliformis y Septoglomus, presentó la mayoría de las morfoespecies indicadoras en todas las áreas de muestreo, con excepción de ADV, donde fue Ambispora sp. Esto puede deberse a que las especies de la familia Glomeraceae producen numerosas esporas de pequeño diámetro en un periodo corto de tiempo, tienen altas tasas de germinación, renovación de hifas, mayor eficiencia en el uso del carbono y tolerancia a condiciones extremas, lo que les confiere una gran capacidad adaptativa y amplia distribución en todo el mundo (Krüger, et al., 2015; Martínez-García, et al., 2015; Trejo, et al., 2016; Stürmer, et al., 2018; Carrillo, et al., 2018; Wang, et al., 2019). La familia Ambisporacea ha sido registrada como indicadora de áreas alteradas y se ha encontrado que las concentraciones de P en el suelo tienen una menor influencia en esta comparada con otras familias de HMA (van der Heyde, et al., 2017; Phillips, et al., 2019; Marihno, et al., 2019). Sin embargo, el efecto de las variables ambientales cambia de un taxón a otro y se necesita una mejor comprensión de la respuesta de algunos géneros y familias frente algún tipo de estrés.
Por otro lado, en diversos trabajos se sugiere que las propiedades químicas, principalmente el pH y los nutrientes como el P, son factores particularmente importantes que afectan fuertemente la distribución de las morfoespecies y la germinación de esporas de micorrizas (Xiang, et al., 2015; Xu, et al., 2017; Sousa, et al., 2018; Tipton, et al., 2018; Day, et al., 2019; Rezácová, et al., 2019; Dudinszky, et al., 2019; Jiang, et al., 2020), y a pesar de que en este estudio no se encontró una influencia estadísticamente significativa de las variables químicas y las comunidades de HMA, se observó en las morfoespecies de la familia Acaulosporaceae una mayor capacidad de establecerse en suelos ácidos (Phillips, et al., 2019; Wang, et al., 2019), como los de RQ, RN y AN. Asimismo, la familia Glomeraceae, capaz de tolerar diferentes condiciones químicas del suelo, se registró en las cuatro áreas evaluadas, así como morfoespecies como Ambispora sp. y F. mosseae, cuya abundancia no se ve alterada en suelos con altas concentraciones de P, como el caso de ADV, además de ser tolerantes a las perturbaciones (van der Heyde, et al., 2017; Zhang, 2016; Mahdhi, et al., 2019).
Los resultados de este estudio evidencian la gran diversidad de morfoespecies de hongos micorrízicos que hacen parte del bosque seco tropical en comparación con otros ecosistemas. Este estudio informa sobre la composición, abundancia y diversidad de HMA basada en el muestreo de áreas perturbadas y naturales del bosque seco en el departamento del Tolima.
Conclusión
La composición, abundancia y diversidad difirieron entre las áreas de muestreo de bosque seco tropical, siendo mayores en la zona afectada por el fuego debido a la presencia de plantas herbáceas altamente micotróficas, como las gramíneas del género Brachiaria, y a la gran adaptabilidad y dominancia de las familias Glomeraceae y Acaulosporacea.
Las áreas naturales se caracterizaron por presentar comunidades de HMA similares a pesar de estar distantes, probablemente por la semejanza en las condiciones edafoclimáticas y la vegetación particular del bosque seco tropical conservado, que permite el establecimiento de morfoespecies raras, sensibles y específicas con afinidad por suelos naturales y una mayor diversidad de huéspedes.

