











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
Introducción
La polinización es una de las interacciones biológicas fundamentales para la supervivencia de las poblaciones de mamíferos y aves, entre las especies animales, así como de muchos tipos de insectos (Ramos-Jiliberto et al., 2020). Esta interacción ha recibido en los últimos años atención por las evidencias que documentan la aparente declinación de las poblaciones de insectos (Goulson, 2019; Wagner et al., 2021), incluidos los polinizadores, debido a factores como los patógenos, el uso excesivo de pesticidas y el cambio climático (Dicks et al., 2021; Parreño et al., 2021). Aunque hay múltiples problemas que afectan la biodiversidad, el declive en las poblaciones de polinizadores podría tener efectos directos en varios ámbitos relacionados con el ser humano, como la economía, la seguridad alimentaria y la disponibilidad de vegetales (Klein et al, 2018). La comunidad científica procura indagar qué especies animales polinizan las especies vegetales, determinar su contribución al éxito reproductivo de las plantas y entender el mecanismo que usan para hacer efectivo el proceso (Amaya-Márquez, 2016), pero, aun así, en ecosistemas altamente diversos como los tropicales esta información es limitada por la gran riqueza de especies que, en muchos casos, está deficientemente documentada (Nates-Parra et al, 2016).
Los países muy diversos como Colombia están expuestos a ese nivel de desconocimiento de las redes de interacción asociadas a la polinización, situación que se acentúa en regiones como la Orinoquia, donde la investigación es escasa (Arbeláez-Cortés, 2013) por factores como el aislamiento geográfico, la ausencia de instituciones con programas sistemáticos de investigación y los problemas de orden público (Agudelo Martínez & Pérez-Buitrago, 2015). Como región biogeográfica, la Orinoquia colombiana abarca el 22 % del territorio nacional y está constituida por diferentes tipos de ecosistemas: bosques de galería, matas de monte y morichales inmersos en una matriz de sabanas de altillanura e inundables con un régimen climático contrastante de temporadas de lluvias y sequía intensas (Rippstein et al, 2001). Por su alta riqueza ecosistémica y biológica la región ha sido considerada un ecosistema estratégico para la humanidad (Correa et al., 2005). Esos atributos han propiciado el interés por explorar sus recursos, lo que ha causado transformaciones en el paisaje a través de la historia, primero, con actividades de ganadería extensiva desde finales del siglo XIX, y luego, en las últimas décadas del siglo XX, con la explotación de hidrocarburos y proyectos de agricultura extensiva de arroz y palma africana (Viloria-de la Hoz, 2009). Por otro lado, las iniciativas de explotación de los recursos de la región no han estado en consonancia con las iniciativas de investigación para entender su biodiversidad y los procesos del funcionamiento de sus ecosistemas.
En la región de la Orinoquia los estudios sobre la comunidad de polinizadores son escasos, lo que es más dramático en taxones como el de las abejas euglosinas (Apinae: Euglossini), que son importantes polinizadores de varias familias de plantas neotropicales como las orquídeas (Bonilla-Gómez & Nates, 1992; Ramírez et al., 2002; Otero & Sandino, 2003). En general, hay muchas brechas en la comprensión de la biología de las euglosinas, las cuales empezaron a ser documentadas apenas en los años 60, cuando se generaron técnicas de captura con compuestos aromáticos y terpenoides como cebos (López, 1963) para este elusivo taxón. Se ha sugerido que los machos de esta tribu de abejas buscan las especies de orquídeas para recolectar recursos aromáticos que funcionan como precursores de compuestos que atraen a las hembras durante la reproducción (Ramírez, et al, 2002; Parra-H, et al., 2016).
El objetivo de este estudio fue documentar la riqueza y la abundancia de especies de abejas de la tribu Euglossini en tres ambientes del norte de la Orinoquia colombiana y describir los cambios en la composición de la comunidad asociados con las temporadas climáticas de sequía y lluvias de la región.
Métodos
Area de estudio
Los especímenes se recolectaron en tres ambientes localizados en la zona rural del municipio de Arauca: sabana inundable (SI) (6°59'26,53"N; 70°36'39,49"W), bosque de galería (BG) (6°59'18,82"N; 70°36'9,87"W) y un terraplén (UN) que sirve como un sendero ecológico con vegetación secundaria que existe hace más de 20 años en la Sede Orinoquia de la Universidad Nacional de Colombia (7° 0'37,98"N; 70°44'37,80"W). El paisaje predominante en el área de estudio es de sabana inundable con presencia de bosques de galería y fragmentos de bosque aislados (Rangel-CH & Minorta-Cely, 2014). La precipitación promedio anual en la zona es de 1.500 mm, con un régimen unimodal compuesto por un periodo de lluvias de ocho meses (abril - noviembre) y uno seco de cuatro meses (diciembre - marzo) (Rippstein et al., 2001).
Recolección de especímenes
Los muestreos se realizaron en las temporadas de sequía (febrero - marzo) y lluvias (abril -mayo) del 2018 durante diez días consecutivos en cada uno de los ambientes y por temporada climática (total 20 días por ambiente). Para la recolección de las abejas se implementaron dos técnicas complementarias. Primero, se dispuso un transecto lineal de 350 m con ocho estaciones cada 50 m; en cada estación se instalaron cuatro trampas de botella modificando el método propuesto por Vélez & Pulido-Barrios (2005). Se usaron botellas de 500 ml, con aberturas de 2,5 cm de diámetro cebadas con 0,5 ml de eucaliptol (Laboratorio San Jorge), metil salicilato (Laboratorio San Jorge), y extracto de vainilla comercial (Levapan), compuestos que han sido ampliamente empleados como atrayentes en estudios previos (Ricklefs et al, 1969; Aguiar & Gaglianone, 2008, 2011; Sydney & Gonçalves, 2015). Además, se empleó extracto de flores secas de Vernonanthura brasiliana (L.) H. Rob. (Asteraceae), una especie aromática común en las sabanas de la región. Segundo, se usó la técnica de "cebos libres" que consiste en colgar un algodón impregnado con 1 ml de atrayente a 1,5 m de altura para capturar directamente las abejas con red entomológica (Nemésio & Morato, 2006). Se hacían cuatro recorridos por día, a las 7:00, 9:00, 11:00 y 14:00 horas, recolectando y registrando los especímenes capturados en las trampas y en los cebos libres. Los especímenes recolectados fueron trasladados y depositados en la Colección Entomológica de la Universidad Nacional de Colombia, sede Orinoquia (CEO), para su limpieza, montaje e identificación hasta el nivel taxonómico más explícito posible usando la clave de abejas euglosinas de Colombia (Bonilla-Gómez & Nates, 1992) y el apoyo de un experto en el taxón. Se siguió la nomenclatura propuesta por Moure et al. (2012) para este grupo taxonómico. La información asociada a los especímenes puede consultarse en la base de datos del SiB Colombia y de la Global Biodiversity Information Facility (GBIF) en el sitio de CEO (Agudelo-Martínez & Pérez-Buitrago, 2020).
Análisis de datos
Se describió la comunidad de Euglossini detallando la cantidad de individuos por especie; se usaron los estimadores no paramétricos Chao 1 y Jackenife 1 para calcular el número de especies probables en la zona de estudio y evaluar la eficiencia del muestreo (Colwell & Coddington, 1994; Chazdon et al., 1998). Para evaluar la diversidad, se calculó el índice de diversidad alfa de Shannon-Wiener usando el logaritmo natural (H') y a partir de este valor se estimó el número efectivo de especies en cada localidad (Jost, 2006). Se calculó el índice de Jaccard para establecer la similitud entre los ambientes muestreados. Con el fin de comparar la riqueza de especies y abundancias en cada uno de ellos se hizo una prueba de Kruskal-Wallis utilizando cada uno de los ambientes como grupos. Para establecer diferencias en la tasa de recolección de especímenes entre las horas de muestreo también se empleó una prueba de Kruskal -Wallis considerando cada hora (7:00, 9:00, 11:00 y 14:00) como un grupo. Para comparar la cantidad de abejas recolectadas por técnica, se hizo una prueba de Yuen contrastando la cantidad de especímenes capturados con cebos libres, y las trampas de botella. Además, se empleó esta misma prueba pareada para comparar la abundancia y la riqueza de las especies en la temporada seca y en la de lluvias. Para visualizar los cambios en la riqueza y la equitatividad de las euglosinas, se elaboraron curvas de rango-abundancia de las temporadas de sequía y lluvias. Los análisis se realizaron con el programa R (R Core Team, 2019) y el cálculo de los estimadores no paramétricos, con el programa EstimateS 9.1.0 (Colwell, 2013).
Resultados
Se recolectaron 367 especímenes pertenecientes a tres géneros y 17 especies. El género mejor representado fue Euglossa, con 12 especies (70,6 %), seguido por Eulaema, con cuatro (23,5 %) y Exaerete con una (5,9 %). La especie más abundante fue Euglossa tridentata Moure 1970, con 129 especímenes (35,1 %), seguida por Eg. modestiorDressler 1982, con 109 especímenes (29,7 %) y Eulaema nigrita Lepeletier 1841, con 67 especímenes (18,3 %). Las especies Eg. chlorina Dressler 1982 y Eg. liopoda Dressler 1982 estuvieron representadas por 15 especímenes cada una, en tanto que Eu. meriana (Olivier 1789) por seis, y Eu. deceptrix Moure 1986 y Eulaema sp1., por cinco cada una. Las especies restantes estuvieron representadas por tres especímenes o menos (Tabla 1). El cálculo de estimadores de riqueza no paramétricos Chao1 y Jackenife 1 indicó que podrían encontrarse entre 19 y 21 especies, respectivamente; así, las 17 especies recolectadas tendrían una representatividad del muestreo que osciló entre el 81 % y el 89,5 % del ensamblaje de abejas de las orquídeas en la zona de estudio. El índice de Shannon fue de 1,6 para el sitio UN, 1,68 para la Sabana, y 1,65 para el bosque, y el número efectivo de especies para las localidades fue de 4,9, 5,3 y 5,2, respectivamente.
Tabla 1 Abundancia y riqueza de especies de abejas euglosinas en cada una de las temporadas climáticas y ambientes muestreados en el municipio de Arauca. Los asteriscos indican especies únicas para cada ambiente. UN: Sendero Universidad Nacional; SI: Sabana inundable; BG: Bosque de galería
La complementariedad según el índice de Jaccard presentó valores medios (UN:SI: 0,60; UN:BG: 0,40; SI:BG: 0,53), lo que indica que cada ambiente albergaba especies únicas (Tabla 1). No se detectaron diferencias en la riqueza de especies entre los tres ambientes (Kruskal-Wallis: F = 5,21; g.l.= 2; p = 0,073), pero sí en las abundancias totales (Kruskal-Wallis: F = 6,24; g.l. = 2; p = 0,044). La prueba de Mann-Whitney con corrección de significación entre grupos indicó que hubo diferencias entre los ambientes de bosque de galería y sabana inundable (p=0,02) (Figura 1, 2a).
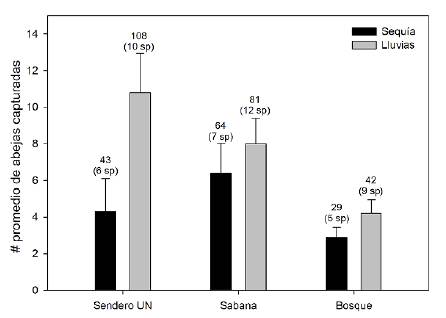
Figura 1 Promedios de abejas euglosinas capturadas en 10 sesiones de muestreo por estación climática en tres ambientes en el municipio de Arauca. Los números sobre las barras indican la abundancia total y la cantidad de especies registradas. Las líneas indican el error estándar
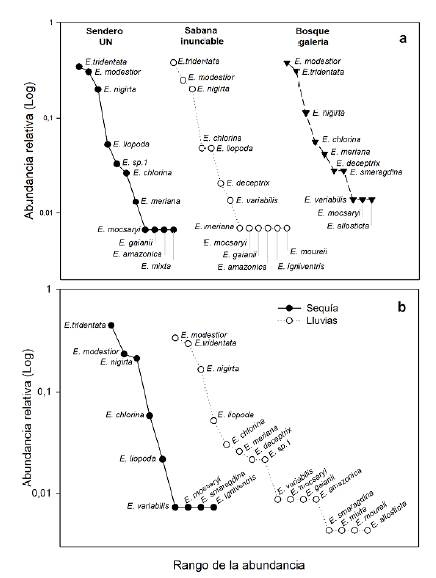
Figura 2 Curvas de rango abundancia de especies de abejas euglosinas para los tres ambientes monitoreados en este estudio el municipio de Arauca (a), y para las temporadas seca y de lluvias en (b)
Las curvas de rango-abundancia en las temporadas climáticas evidenciaron una mayor riqueza de especies en la temporada de lluvias (16 especies) con respecto a la temporada seca (9 especies) (Figura 2b). Las especies más abundantes de abejas euglosinas en la sabana inundable fueron Eg. tridentata, Eg. Modestior y El. nigrita, en tanto que Eg. liopoda y Eg. Chlorina registraron abundancias intermedias. El número de especies exclusivas de la temporada de lluvias fue de ocho (El. meriana, Eg. deceptrix, El. sp1, Eg. gaianii, Eg. amazónica, Eg. mixta, Eg. mourei, Eg. allosticta), y en el periodo de sequía solo fue exclusiva Eg. igniventris en el ambiente de sabana inundable (Tabla 1).
La prueba de Yuen pareada indicó que la riqueza y la abundancia de especies fueron mayores en la época de lluvias que en la sequía (F = -3,81, g.l. = 23, p = 0,001 y F = -2,65, g.l. = 23; p = 0,016, respectivamente) (Figura 1). Se registraron diferencias en las horas de captura de las_abejas (Kruskal-Wallis: F = 32,9, p<0,05), siendo las 9:00 la hora de mayor recolección (x̅ = 4,92 ± 4,21; rango = 0 - 20), seguida por las 11:00 (x̅ = 2,57 ± 3,1; rango = 0 - 13_). Las horas con menos capturas fueron las 7:00 (x̅ = 1,27 ± 1,7; rango = 0 - 7) y las 14:00 (x̅ = 1,16 ± 1,9; rango = 0 - 7) (Figura 3). Las trampas de botella tuvieron un mejor desempeño en la captura de euglosinas (X = 6,02 ± 3,78, rango = 0 - 17) que la recolección de especímenes con red entomológica (x̅ = 3,89 ± 4,07; rango = 0 - 17) (prueba de Yuen: W = -3,79; g.l. = 22, p = 0,001). El eucaliptol fue el atrayente más eficiente, con 359 capturas (97,8 %), seguido del metil-salicilato con siete especímenes (1,9 %), en tanto que el extracto de V. brasiliana solo atrajo un espécimen (0,27 %) y el extracto de vainilla ninguno.
Discusión
En las últimas décadas se ha incrementado en Colombia el número de especies de abejas euglosinas reportadas, pasando de 108 especies en 1992 (Bonilla-Gómez & Nates-Parra, 1992) a 113 (Ramírez et al., 2002), 134 (Parra-H & Nates-Parra, 2012) y, más recientemente, a 174 especies (Chilito-Rodríguez et al., 2021). Esta cifra posiciona a Colombia como uno de los países más diversos en este taxón, junto con Brasil (200 spp.: Olivera-J et al., 2015), Ecuador (115 spp.: Padrón, et al., 2018), Perú (77 spp.), Venezuela (70 spp.) y Panamá (69 spp.) (Chilito-Rodríguez et al., 2021).
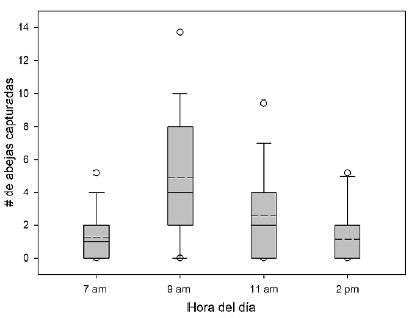
Figura 3 Cantidad de abejas euglosinas recolectadas por horas en el municipio de Arauca, Colombia. Las líneas discontinuas representan el promedio
En este estudio, el primero en el departamento de Arauca, se recolectaron 367 machos de tres géneros y 17 especies de abejas de la tribu Euglossini, con un buen grado de completitud en el muestreo según los estimadores Chao1 y Jacknife 1, correspondiente al 9,7 % de la riqueza nacional. La literatura sobre las localidades colombianas es limitada en gran parte del territorio nacional, pero en un estudio en el piedemonte llanero se reportaron 17 especies de 26 conocidas en la zona (Parra-H & Nates, 2007). En los departamentos amazónicos el rango de especies registradas es amplio: 43 especies en Caquetá, 36 en Amazonas, siete en Putumayo y Vichada, cinco en Guainía, tres en Vaupés, y cero en Guaviare (Chilito-Rodríguez et al., 2021), cifras que denotan la ausencia de estudios en muchas localidades e impiden tener una idea más precisa de la riqueza real que el país alberga. En otros departamentos como el Chocó se han reportado 31 especies (Otero y Sandino, 2003) y en Antioquia entre 21 (Arcos, 2020) y 50 (Ramírez et al., 2015). En localidades de otros países con más tradición de estudios de abejas Euglossini como Costa Rica se han registrado entre 35 (Ramírez, 2019) y 40 especies (Santos Murgas et al., 2018) , en Panamá entre 15 y 28 (Ricklefs et al., 1969; Osorio-Arenas & Santos Murgas, 2017; Reyes-Ledezma et al., 2020; Vega-Hidalgo et al., 2020) y en Perú, 31 (Allen et al., 2019) . Brasil es posiblemente el país con más arraigo en la investigación de Euglossini; en múltiples localidades de Rio de Janeiro se han registrado entre 11 y 17 especies (Aguiar & Gaglianone, 2011), en Maranhao entre nueve (Rebélo & Cabral, 1997) y 14 (Silva, et al., 2009), y en la Mata Atlántica, nueve (Sofia, et al., 2004), 23 (Rocha & Garófalo, 2013), y 12 (Rocha et al., 2020), en tanto que en Mato Grosso, en el ecotono cerrado-selva, se han registrado 16 (Olivera-Junior et al., 2015), en la sabana 14 (Viana et al, 2021), en la catinga, cinco (Carneiro et al., 2018) y en bosques higrófilos, cinco especies (Castro et al., 2013). Así, la riqueza de 17 especies registrada en Arauca es notable y comparable a ambientes estructuralmente más complejos (por ejemplo, los bosques), a pesar de ser un ecosistema predominantemente compuesto de sabanas con fragmentos discretos de bosques donde las euglosinas pueden encontrar los recursos que las orquídeas y otras especies vegetales les proveen, ya que probablemente en la sabana la oferta es limitada.
El índice de Jaccard indicó que, aunque los tres ambientes comparten especies abundantes como Eg. tridentata, Eg. modestior y El. nigrita (Figura 2a), el bosque es el ambiente más disímil en riqueza de especies. Por otra parte, la sabana y el sitio perturbado fueron más similares entre sí, lo que podría deberse al hecho de compartir características abióticas similares al ser ambientes abiertos y soleados. En cuanto a la riqueza de especies, esta fue similar en los tres ambientes durante los cuatro meses de estudio (Tabla 1). Este resultado fue contrario a lo esperado, pues, en general, ambientes con poca estructura vegetal (como en este caso la sabana inundable) suelen tener menor número de especies. En este sentido, Rasmussen (2009) no registró diferencias entre los diversos hábitats y lo atribuyó a la corta duración del estudio (< 3 meses). Además, Faria & Gonçalves (2013), en la costa oriental de Brasil, y Castro et al. (2013) plantearon que la precipitación incide en la mayor oferta de recursos florales, lo que contribuye a determinar la riqueza de especies de abejas y propicia ensamblajes más diversos en zonas de pastos mezcladas con bosques. Sin embargo, es amplio el número de investigaciones que postulan que las euglosinas son más diversas en ambientes en buen estado de conservación (Parra-H & Nates, 2007; Aguiar & Gaglianone, 2011; Rocha & Garófalo, 2013; Carneiro et al., 2018; Razo-León et al., 2018; Chilito-Rodríguez et al., 2021; Viana et al., 2021) debido a la mayor diversidad botánica (Knoll & Penatti, 2012; Villaça et al., 2009) y la oferta de recursos como polen, néctar, y materiales para la construcción de nidos (Viana et al., 2021). Esta tendencia de mayor diversidad de la tribu Euglossini en ambientes bien preservados ha llevado a que este taxón sea considerado como bioindicador de la calidad del hábitat (Rocha-Filho & Garófalo, 2013; Parra-H et al., 2016; Allen, 2019).
Otros estudios han detectado un patrón opuesto a lo esperado en términos de riqueza. Por ejemplo, Otero & Sandino (2003) reportaron una baja riqueza en bosques primarios comparada con bosques secundarios y granjas de cultivo en el Chocó y sugirieron que las abejas posiblemente prefieren forrajear en sitios abiertos, aspecto que no documentamos en este estudio. Aunque la Orinoquia está expuesta a altos niveles de deforestación, el paisaje orinocense es naturalmente fragmentado en lo que respecta a las formaciones boscosas que alberga, un factor que posiblemente ha propiciado el desarrollo de mecanismos de comportamiento en las abejas euglosinas para lidiar con esa configuración del paisaje. En particular, la alta vagilidad de las abejas (entre 23 y 40 Km: Ricklefs et al., 1969; Janzen, 1971, Wikelski et al., 2010) les permitiría moverse eficientemente entre fragmentos en busca de los recursos para su sostenimiento, y explicaría la similitud en el número de especies registradas en los tres ambientes. De ser así, es posible que la escala espacial del estudio no haya sido lo suficientemente amplia, es decir, los tres sitios representan un solo ensamblaje de euglosinas dado que la distancia máxima lineal entre las localidades es de 15,8 Km.
La menor abundancia de euglosinas se registró en el bosque, en tanto que en el sitito con intervención intermedia (UN) y en la sabana inundable fue mayor (Tabla 1, Figura 1). En concordancia con este resultado, se ha planteado que el comportamiento de forrajeo incluye desplazamientos a través de sitios abiertos entre fragmentos (Razo-León et al., 2018) y que la heterogeneidad ambiental favorece la integridad de las comunidades de abejas euglosinas (Carneiro et al., 2021). Alternativamente, la menor abundancia registrada en el bosque podría ser consecuencia de las menores temperaturas del sotobosque, lo cual induce una menor volatilidad de los cebos, reduciendo las tasas de captura en este ambiente. Sin embargo, otros estudios documentan mayores abundancias en bosques por ser ambientes sombreados con temperaturas más frescas (Silva et al., 2009; Carneiro et al., 2018; Allen, 2019) y plantean que la fragmentación es percibida como un obstáculo por algunas especies de euglosinas (Aguiar & Gaglianone, 2011).
En concordancia con otros estudios, en este se registraron valores mayores de riqueza y abundancia durante la época de lluvias (Rebêlo & Cabral, 1997; Sofia et al., 2004; Silva et al., 2009; Castro et al., 2013; Carneiro et al., 2018). Este patrón ha sido documentado en variados taxones de insectos (Wolda, 1988; Franzen et al., 2017; Habel et al., 2018) y está asociado con la mayor oferta de alimento y otros productos como consecuencia de la disponibilidad de recurso hídrico para las plantas (Ramírez et al., 2015), las bajas temperaturas en la época de lluvias comparadas con la época seca (Rebêlo & Cabral, 1997) y por tener un efecto en los periodos de anidación y emergencia de las abejas (Ackerman, 1983). Silva et al. (2009) sugirieron que estudiar la fenología de las orquídeas puede ayudar a entender la interacción euglosina-planta, lo que es relevante en países como Colombia, donde se encuentra la mayor diversidad de especies de Orchidaceae (Alba-Patiño et al., 2021).
En relación con las horas del día, se ha documentado una mayor frecuencia de capturas de abejas euglosinas en la mañana (Dressler, 1982; Olivera-Junior et al., 2015), lo que se atribuye a que en este periodo del día la producción de compuestos secundarios por las plantas es más intensa (Oliveira, 1999). En este estudio la máxima tasa de captura fue a las 9:00, lo que ratifica esta tendencia. El reactivo más eficiente como cebo de atracción fue el eucaliptol (97,8 %), un compuesto ampliamente empleado en estudios de euglosinas (Peruquetti et al., 1999; Silva & Rebêlo, 1999; Brito & Rêgo, 2001; Sofia et al., 2004; Silva et al., 2009). Sin embargo, las disparidades en las tasas de captura con diversos atrayentes en diferentes estudios sugieren que algunas especies parecen responder a ciertos atrayentes, pero a otros no (Nemesio & Silveira, 2004; Castro et al., 2013). Por ello la comunidad científica sigue explorando compuestos químicos que permitan documentar con más precisión la diversidad de este taxón de abejas. Se sugiere que en futuros estudios en la Orinoquia se empleen compuestos adicionales.
Conclusiones
En este estudio se documentan por primera vez algunos aspectos de la ecología básica de las abejas euglosinas en relación con el ciclo de precipitación anual en el departamento de Arauca. Es notable la riqueza de especies (17) y comparable con ecosistemas estructuralmente más complejos, considerando que el área de estudio es un paisaje donde predomina el ecosistema de sabana. Por el importante rol de las abejas de la tribu Euglossini como polinizadores, es imperativo minimizar actividades antrópicas en los ecosistemas de la Orinoquia como la deforestación, las quemas anuales y la fumigación, dados los efectos que estas prácticas tienen en la oferta de hábitats disponibles para este taxón, lo que afecta de forma directa su supervivencia. Sin embargo, a partir de la literatura disponible se hace evidente que no existen patrones bien establecidos sobre diversos aspectos ecológicos de este taxón, por ejemplo, los factores que determinan la riqueza de especies, su abundancia, el uso del hábitat y sus múltiples interacciones bióticas y abióticas. Esto implica la necesidad de desarrollar más investigaciones sobre las abejas Euglosinas para mejorar la comprensión acerca de su biología, determinar su rol en el funcionamiento de los ecosistemas que habitan y generar estrategias para garantizar la supervivencia de sus poblaciones.

