










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Antropología
Print version ISSN 0486-6525
Rev. colomb. antropol. vol.48 no.1 Bogotá Jan./June 2012
COANCESTRÍA DE APELLIDOS Y LINAJES DEL CROMOSOMA Y EN EL NOROESTE DE COLOMBIA: UNA HERRAMIENTA ÚTIL PARA ESTABLECER MIGRACIÓN ENTRE POBLACIONES
SURNAMES AND Y CHROMOSOME COANCESTRY IN NORTHWEST COLOMBIA: A USEFUL TOOL TO ESTABLISH MIGRATION BETWEEN POPULATIONS
WINSTON ROJASA,
OMER CAMPOA, JENNY GARCÍAB, IVÁN SOTOA, CONSTANZA DUQUEA,
GABRIEL BEDOYAA, ANDRÉS RUIZ-LINARESA,C
ALABORATORIO DE GENÉTICA MOLECULAR, INSTITUTO DE BIOLOGÍA, UNIVERSIDAD DE ANTIOQUIA, MEDELLÍN, COLOMBIA. (winstonrojas@yahoo.com)
BLABORATORIO DE PSIQUIATRÍA, UNIVERSIDAD DE ANTIOQUIA, MEDELLIN, COLOMBIA.
CDEPARTAMENT OF GENETICS, EVOLUTION AND ENVIRONMENT,UNIVERSITY COLLEGE LONDON, LONDRES, REINO UNIDO.
Recibido: 10 de septiembre de 2011 Aprobado: 12 de diciembre de 2011
Resumen
Por medio del análisis de marcadores genéticos del cromosoma Y, se estudió la composición de los linajes y su distribución en los apellidos más comunes en una muestra de 471 hombres provenientes del municipio de Marinilla (Antioquia) y su zona de influencia y del municipio de Aranzazu (Caldas), y en un grupo de muestras de la población general de Medellín. Además de encontrar una tasa variable de coancestría entre apellidos y linajes del cromosoma Y, también se detectó una gran similitud en el patrón de distribución de haplogrupos/haplotipos/apellidos entre Marinilla y su zona de influencia y Aranzazu, hallazgo que refuerza la idea de la migración histórica entre estas dos regiones. Esta similitud indicaría además que en las dos poblaciones pueden circular variantes genéticas comunes vinculadas a enfermedades humanas.
Palabras clave: apellidos, cromosoma Y, paternidad.
Abstract
Using Y chromosome genetic markers we studied the lineage composition and distribution in the most common surnames in 471 males from the municipality of Marinilla and its zone of influence, the municipality of Aranzazu (department of Caldas) and a group of samples from the general population of Medellin (Antioquia). Despite a variable rate of coancestry between surnames and Y-chromosome markers, we found a high similarity in the pattern of distribution of haplogroups/haplotypes/ surnames between Marinilla and its zone of influence and Aranzazu, reinforcing the historical migration between these two regions. This similarity would indicate that in both populations may be circulating common genetic variants linked to human diseases.
Key words: surnames, Y chromosome, paternity.
INTRODUCCIÓN
Entre los movimientos de expansión territorial que hubo en Antioquia desde el periodo colonial, un acontecimiento importante fue el establecimiento de poblaciones de pequeños agricultores en el altiplano de la región oriental cercana a Medellín (capital de Antioquia), que llevó desde el año 1750 a la fundación de los municipios de Rionegro y Marinilla (Parsons 1968; Zapata-Cuéncar 1941). Posteriormente, los habitantes de Marinilla influyeron en la fundación de otros municipios como El Santuario, El Peñol, Carmen de Viboral, Granada, Cocorná y Guatapé (Parsons 1968), y se constituyó así el área de Marinilla y su zona de influencia (MZI) (Bedoya, García et ál. 2006; Soto 2003). En 1787 otro movimiento de expansión, denominado colonización antioqueña, se inició desde Antioquia, incluida la MZI, hacia la región centro-oeste de Colombia. Esta migración estableció nuevos asentamientos en la región hoy conocida como Eje Cafetero, que abarca los departamentos de Caldas, Risaralda, Quindío, Tolima y Valle del Cauca (Álvarez 1996). Los habitantes de Antioquia, incluida la MZI, participaron en la fundación de varios centros urbanos en Caldas, por ejemplo de su capital Manizales y de los municipios de Aguadas, Salamina, Salento, Chinchiná y Aranzazu. Estudios previos que comparan los censos de 1824 en Aranzazu y de 1851 en Marinilla informaron que ambas poblaciones compartían diez de sus apellidos principales, lo que sugiere que las familias de Marinilla contribuyeron a la fundación de Aranzazu (Arango-Mejía 1993). Cabe destacar que los actuales habitantes de estas poblaciones comparten aproximadamente el 46% de sus apellidos (Bedoya, Montoya et ál. 2006). Estudios anteriores sobre la MZI indican que su diversidad genética y de apellidos es compatible con un escaso número de hombres fundadores (Bedoya, García et ál. 2006; Bedoya, Montoya et ál. 2006; Soto 2003).
La transmisión patrilineal del primer apellido, en muchas poblaciones, simula la transmisión de un alelo neutral a través de la región no recombinante del cromosoma Y (King y Jobling 2009a). Se espera que los hombres que comparten el primer apellido compartan también el linaje del cromosoma Y, y por ello el análisis de los apellidos puede ser una herramienta útil para examinar la composición poblacional, el flujo genético y la endogamia (Bowden et ál. 2008; Jobling 2001; King y Jobling 2009a, 2009b; Piazza et ál. 1987; Zei et ál. 2003). Sin embargo, es posible que una fracción de los apellidos compartidos no se correlacione con la ascendencia genética del cromosoma Y, por ejemplo, un apellido puede tener múltiples orígenes genéticos o un linaje genético puede contener varios apellidos. Además, la adopción y la infidelidad pueden alterar las correlaciones esperadas entre ambos marcadores (Bowden et ál. 2008; Jobling 2001).
En este estudio se analizó el patrón de distribución de los linajes del cromosoma Y en el primer apellido de personas con ancestros originarios de la MZI y Aranzazu, y se comparó este patrón con el de una muestra general de Antioquia. A pesar de encontrar un grado variable de paternidad, medido como las correlaciones entre linajes genéticos y apellidos, cuando las muestras fueron estratificadas por apellido se encontró una fuerte correlación en la composición y la distribución de haplogrupos/haplotipos del cromosoma Y en los apellidos de ambas poblaciones, hallazgo que apoya la idea de la migración histórica entre la MZI y Aranzazu.
MATERIALES Y MÉTODOS
Muestras
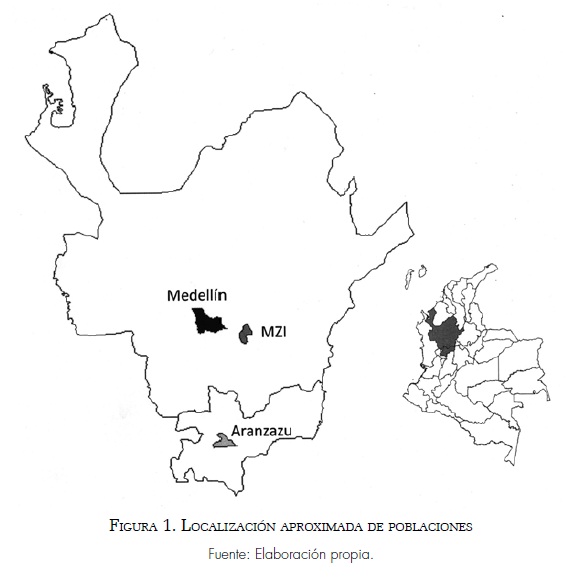
En total se incluyeron muestras de ADN de 497 personas voluntarias mayores de 18 años, agrupadas de acuerdo con su origen en Marinilla y su zona de influencia (MZI), Antioquia (personas de otros municipios de Antioquia excepto la MZI) y Aranzazu (departamento de Caldas). Varias muestras incluidas se analizaron en estudios previos (Bedoya, García et ál. 2006; Bedoya, Montoya et ál. 2006; Carvajal-Carmona et ál. 2000; Rojas et ál. 2010). La muestra de la MZI incluye individuos provenientes de Marinilla, El Peñol, Santuario, Cocorná, Carmen de Viboral, Granada y Guatapé. Las muestras de la MZI y de Aranzazu fueron colectadas en un anterior estudio epidemiológico sobre el síndrome bipolar. En la figura 1 se observa la ubicación aproximada del origen de las muestras. Marinilla y Medellín están distanciados por 28 km; Marinilla y Aranzazu por 102 km y Medellín y Aranzazu por 108 km. Una entrevista genealógica estándar se utilizó para confirmar que los individuos de la muestra no estuvieran relacionados consanguíneamente (por lo menos hasta tres generaciones) y que por lo menos cuatro de ocho ancestros fueran de origen local. Todos los individuos incluidos en este estudio firmaron un consentimiento informado y la aprobación ética fue proporcionada por el Comité de Ética de la Universidad de Antioquia.
Análisis de apellidos
En los tres grupos se analizó el primer apellido de 497 hombres y en cada grupo se estimó la diversidad de apellidos como el número de apellidos en la muestra total. Además se obtuvo el estimador alfa de Fisher, descrito en Barrai et ál. (2001), el cual representa el número efectivo de apellidos y es definido como:
α=1/Iii [donde Iii=Σnik (nik-1)/Ni(Ni-1)]
donde nik es la frecuencia absoluta del apellido k en la muestra i, con un tamaño N. Un valor pequeño de a indicaría un alto nivel de consanguinidad y baja inmigración, mientras que un valor alto sugeriría un bajo nivel de consanguinidad y una alta inmigración (Barrai et ál. 2001). La distancia euclidiana entre grupos se estimó como:
E=√(1-Σk √pkipkj)
donde pki y pkj son las frecuencias relativas del apellido k en las muestras i y j, respectivamente (Cavalli-Sforza y Edwards 1967). La distancia euclidiana es la unidad cuando dos grupos no comparten apellidos.
Análisis del cromosoma Y
Las muestras de ADN fueron extraídas por procedimientos de laboratorio de rutina, que fueron descritos previamente (Bedoya, García et ál. 2006; Carvajal-Carmona et ál. 2000). Para definir los linajes paternos mayores del cromosoma Y (haplogrupos), se emplearon ocho marcadores genéticos binarios (de evolución lenta) en ensayos de reacción en cadena de la polimerasa combinados con polimorfismos en la longitud de fragmentos de restricción, utilizando la actual nomenclatura del cromosoma Y (Karafet et ál. 2008). Estos marcadores se han definido jerárquicamente y permiten la identificación de los principales linajes en poblaciones continentales europeas, africanas y nativas americanas. El estado derivado en el marcador YAP sumado al estado derivado M2 definen el haplogrupo E1b1a, y sumado al estado ancestral M2 definen el haplogrupo DE*(xE1b1a); el estado ancestral en M9 define el haplogrupo F*, y sumado al estado ancestral en 92R7 define K*(xP); o sumado al estado derivado en 12f2a define el haplogrupo J; el estado derivado 92R7 y el ancestral en M167 definen el haplogrupo P*(xQ); el estado derivado M167 define R1b1b2d; el derivado en M242 define el haplogrupo Q*(xQ1a3a), y sumado al estado derivado en M3 define Q1a3a.
Los linajes indicados con un asterisco (paragrupos) son potencialmente parafiléticos. Ambos haplogrupos Q son los principales linajes presentes en grupos nativos americanos. El linaje E1b1a incluye los haplogrupos más frecuentes en África y se considera como la contribución africana paterna; el linaje J tiene una frecuencia alta en las poblaciones del este y centro del Mediterráneo (Di Giacomo et ál. 2004). Los restantes haplogrupos tienen una alta frecuencia en poblaciones europeas (aunque algunos de ellos se han observado en el este de Asia, Oceanía, Indonesia y Australia). La mayoría de los linajes de Europa occidental pueden ser asignados al haplogrupo P*(xQ); el haplogrupo R1b1b2d tiene sus más altas frecuencias en poblaciones vascas y catalanas (Flores et ál. 2004).
Además de los linajes mayores se tipificaron seis marcadores genéticos microsatélites de la región no recombinante del cromosoma Y, que consisten en polimorfismos de repeticiones cortas (DYS19, DYS388, DYS390, DYS391, DYS392 y DYS393); estos marcadores fueron útiles para evaluar la diversidad y composición de los haplogrupos del cromosoma Y, como se describió previamente (Bedoya, García et ál. 2006; Thomas, Bradman y Flinn 1999). En cada muestra se estimó la composición y frecuencia de haplotipos microsatélites (tabla suplementaria 1). La diversidad genética de los haplogrupos y haplotipos del cromosoma Y en cada grupo se obtuvo mediante el uso del programa Arlequin versión 3.1 (http://cmpg.unibe.ch/software/arlequin3) de Excoffier, Laval y Schneider (2005).
El mismo apellido en dos haplotipos que difieren en solo un paso mutacional podría reflejar un origen ancestral común, con divergencia debido a una mutación reciente (Bedoya, García et ál. 2006); en consecuencia, se definió la tasa de paternidad (TP) dentro de cada apellido como la frecuencia del haplotipo modal más aquellos haplotipos que difieren a un paso mutacional, todos ellos asignados al mismo haplogrupo. La tasa de no paternidad se estimó como TNP = TP − 1. La distribución de los apellidos en los haplotipos de cada haplogrupo se obtuvo a través de redes, usando el programa Network v. 2.0 (http://fluxus-engineering.com) de Bandelt, Forster y Rohl (1999).
RESULTADOS
Apellidos
En un total de 497 hombres se encontró un total de 115 apellidos diferentes, 16 de los cuales (con una frecuencia de 38,6%) se observaron simultáneamente en los tres grupos. Cada grupo tiene apellidos no presentes en ningún otro (tabla 1 y tabla suplementaria 2). En el grupo de Antioquia se encontró la mayor diversidad de apellidos. Los grupos de la MZI y Aranzazu compartieron 35 apellidos (78,9 y 70% respectivamente), mientras que la MZI y Antioquia compartieron 26 (42,7% y 60,7, respectivamente) y Aranzazu y Antioquia compartieron 21 (49,5 y 50,8% respectivamente) (tabla 1). Los apellidos más frecuentes en la muestra total y en cada sitio, además del estimativo a, se presentan en la tabla 1 y en la tabla suplementaria 2. Los 15 apellidos más frecuentes sumaron 63,8%, 54,8% y 47,6% en la MZI, Aranzazu y Antioquia, respectivamente. De estos apellidos de alta frecuencia la MZI y Aranzazu comparten ocho, la MZI y Antioquia comparten seis y Aranzazu y Antioquia comparten siete. Entre la MZI y Aranzazu se encontró la menor distancia euclidiana (0,57), mientras que la MZI y Antioquia, y Aranzazu y Antioquia, tuvieron distancias similares (0,73 y 0,72, respectivamente).

Cromosoma Y
Las frecuencias de nueve haplogrupos del cromosoma Y son muy similares en las tres muestras (tabla suplementaria 1). En promedio, las frecuencias de los linajes Q nativos americanos y del linaje E1b1a africano fueron muy bajas; dentro de los linajes europeos (aproximadamente el 96% del total), el haplogrupo P*(xR1b1b2d) fue el más frecuente, seguido de los haplogrupos J y K*(xP).
Dentro de los 9 haplogrupos del cromosoma Y se encontraron 120 haplotipos microsatélites (tabla suplementaria 1), de los cuales 74 se encontraron en la MZI, 53 en Aranzazu y 38 en Antioquia. Setenta
haplotipos se encontraron una sola vez y 40 fueron compartidos por todas las muestras (54,7%): 27 haplotipos fueron compartidos entre la MZI y Aranzazu (75,2 y 74,7%, respectivamente), 17 entre la MZI y Antioquia (60,2 y 64%, respectivamente) y 14 entre Aranzazu y Antioquia (50 y 59,1%, respectivamente). El haplotipo/haplogrupo más frecuente (haplotipo 1-P*(xQ), cuya composición de alelos microsatélites es 14-12-24-11-13-13) en la muestra total alcanzó una frecuencia de 25,8%; se encontró en la MZI (30,1%), Aranzazu (22,1%) y Antioquia (19,7%). El segundo haplotipo/haplogrupo más frecuente (haplotipo 2, 11,3%) estuvo presente en los haplogrupos P*(xQ) y R1b1b2d. La diversidad en la composición de haplogrupos, así como la diversidad de haplotipos, es mayor en el grupo de Antioquia.
Coancestría entre apellidos y cromosoma Y
Algunos individuos con apellidos diferentes portaron el mismo haplotipo (tabla 2, figura 2); además, muchos haplotipos no relacionados fueron vinculados a un apellido (tabla 3, tabla suplementaria 1). Cuando se asumió un modelo mutacional por pasos (Kimura y Ohta 1978), la correlación entre haplotipos/haplogrupos/ apellidos (expresada como la tasa de paternidad) fue menor en Antioquia, seguida por la MZI (tabla 3). Sin embargo, esta correlación fue variable entre los apellidos más frecuentes. Por ejemplo, en 45 individuos que portaban el apellido Giraldo, 34 portaban dos haplotipos estrechamente relacionados (haplotipos 1-P*(xQ) y 2-P*(xQ); tasa de paternidad ~76%). Dentro de los apellidos Duque, Ramírez, Castaño, Gómez y Botero la tasa de paternidad fue más alta (> 80%), mientras que fue muy baja en Quintero (33,3%), Cardona (50%), Alzate (56,3%) y García (55,8%). El haplotipo 1-P* (xQ) está presente en 11 de los 15 apellidos más frecuentes y se encontró de forma simultánea en todas las muestras que portaban el apellido Gómez; sin embargo, en la MZI y Aranzazu este haplotipo fue compartido como haplotipo de paternidad dentro de los apellidos Giraldo, Ramírez, Alzate, Quintero, González y Castaño, así como haplotipo compartido de no paternidad en los apellidos Zuluaga, Salazar y Botero.
Al analizar los 15 apellidos más frecuentes, los siguientes haplotipos/ haplogrupos nucleares son evidentes: haplotipo 1-P (xQ) con los apellidos Giraldo, Gómez, Ramírez, Alzate, González y Castaño; haplotipo 3 DE*(xE1b1a) con García; haplotipo 4-F* (xK) con Zuluaga; haplotipo 8-K*(XP) con Salazar; haplotipo 2-R1b1b2d con Duque; haplotipo 7-K*(XP) con López; haplotipo 15-P*(xQ ) con Jiménez; haplotipo 10-J con Botero y el haplotipo 11-J con Cardona (tabla 3). Cuando se consideran los apellidos menos frecuentes (datos no mostrados), se observan las siguientes correlaciones, todas ellas compartidas entre la MZI y Aranzazu: haplotipo 2-P*(xQ)/Aristizábal, 6-P*(xQ)/Tamayo, 11-P*(xQ)/Jaramillo, 9-K*(xP)/Carvajal; 31-J/Montes y 13-J/Serna. En la MZI y Antioquia se observaron simultáneamente las siguientes correlaciones: haplotipo 1-P*(xQ)/Echeverry, 40-P*(xQ)/Franco; 2-P*(xQ)/Montoya y 19-P*(xQ)/Suárez; mientras en Aranzazu y Antioquia se observaron las correlaciones: haplotipo 16-DE*(xE1b1a)/Londoño, 1-P*(xQ)/Marín y 1-P*(xQ)/Valencia.
Todos los cromosomas Y nativos americanos (8 individuos asignados a haplogrupos Q) y africanos (10 individuos asignados al haplogrupo E1b1a) fueron portadores de alguno de los apellidos más frecuentes (Giraldo, Gómez, García, Castaño, Quintero, Salazar y Alzate). Teniendo en cuenta que la no paternidad se ha acumulado durante 250 años desde la fundación de la población (10 generaciones, para un tiempo de 25 años por cada generación), el promedio de la tasa de no paternidad fue de ~1,07x10-3/ generación, consistente con previos reportes (Bedoya, García et ál. 2006; Macintyre y Sooman 1991; Sykes e Irven 2000).
DISCUSIÓN
El análisis de los apellidos patrilineales es una potencial herramienta complementaria del análisis de marcadores genéticos para investigar el flujo y la estructura de las poblaciones humanas recientes y contemporáneas (Barrai et ál. 1996; Barrai et ál. 2000, 2001; Cabral et ál. 2005; Castro de Guerra, Arvelo y Pinto-Cisternas 1999; Dipierri et ál. 2005; González-Martín et ál. 2006; Rodríguez-Larralde et ál. 1993; Rodríguez-Larralde, Barrai y Alfonzo 1993). Sin embargo, el supuesto de que un apellido común entre individuos también implica una ascendencia genética compartida ha sido probado solamente en algunas poblaciones (Bedoya, García et ál. 2006; Bowden et ál. 2008; Zei et ál. 2003; King y Jobling 2009b). En este estudio se analizaron marcadores de ADN binarios de evolución lenta, simultáneamente con marcadores microsatélites polimórficos de evolución rápida, en una muestra de individuos provenientes de Antioquia, la MZI y Aranzazu, para estimar la coancestría de estos marcadores con los apellidos paternos.
Marinilla se estableció como municipio en 1750 y fue fundado por don Juan Duque de Estrada y Francisco Manzueto Giraldo (Zapata-Cuéncar 1941). El núcleo colonizador de Marinilla hacia Caldas fue iniciado por apellidos como Duque, Giraldo, Aristizábal Botero, García, Gómez, González, Ramírez y Hoyos (Arango-Mejía 1993), algunos de los cuales fueron los apellidos más frecuentes en la muestra de la MZI y Aranzazu; esta constatación muestra la persistencia de los apellidos fundadores en la población de Aranzazu. La alta frecuencia de apellidos como Giraldo en ambas poblaciones sugiere que no se han visto afectados por el crecimiento demográfico, a diferencia de apellidos como Ramírez y Duque, cuya menor frecuencia en Aranzazu apunta a una deriva en un corto tiempo, en contraste con su alta frecuencia en la población de origen.
Los resultados del estudio muestran que muchos apellidos presentes en estas poblaciones son de origen polifilético. Debido a la naturaleza y la abundancia de muchos apellidos patronímicos (por ejemplo García, Gómez, González, López y Ramírez), no se espera hallar una correlación entre los haplotipos/haplogrupos y los apellidos, ya que muchos apellidos se encontraron en varios linajes genéticos sin relación de parentesco entre ellos. Sin embargo, la similitud en el patrón de distribución de los apellidos en la MZI (Antioquia) y Aranzazu (Caldas) proporciona una fuerte evidencia del flujo genético histórico entre ellas: los apellidos más frecuentes en la MZI y Aranzazu también portan el haplotipo modal más frecuente; y algunos de estos haplotipos/apellidos no se encuentran en la muestra de Antioquia. Además, la MZI y Aranzazu son también los grupos con menor diversidad tanto en los apellidos como en el cromosoma Y. Aunque no es posible determinar si todos los territorios entre estos dos departamentos comparten estas características, el análisis de los apellidos podría complementar los estudios genéticos sobre la estructura y el origen de los emigrantes en poblaciones de Antioquia y Caldas.
Otro aspecto relevante es que se encontraron varios haplotipos no relacionados dentro de un apellido, lo cual representa la tasa de no paternidad acumulada en el pasado; esta tasa es variable dentro de los apellidos examinados. Esto sugiere que en la dinámica social del periodo colonial existió una selección social de algunos apellidos: portar ciertos apellidos representaba acceder a importantes beneficios sociales en la población mestiza en formación. Sin embargo, otra explicación posible consiste en que el linaje genético ancestral correspondiente al apellido haya acumulado suficiente variación genética y por ello el apellido fundador pertenezca a varios linajes. Además, el hecho de que todos los individuos portadores de haplogrupos nativos americanos y africanos sean portadores de apellidos europeos sugiere un alto índice de no paternidad en estos componentes ancestrales, que confirma que estas personas adoptaban el apellido de sus propietarios.
Algunos estudios anteriores realizados en Antioquia mostraron una mezcla interétnica genética principalmente de linajes paternos provenientes de Europa (Bedoya, García et ál. 2006; Carvajal-Carmona et ál. 2000; Rojas et ál. 2010). En este estudio, la mayoría de los cromosomas Y se asignaron al haplogrupo europeo P*(xQ), que alcanza sus mayores frecuencias en Europa occidental. Sin embargo, los resultados mostraron una importante frecuencia de los haplogrupos R1b1b2d y J, con una alta frecuencia en poblaciones vascas y sefardíes, respectivamente. Esto no es un patrón propio de poblaciones de Antioquia y Caldas, tal como se reportó recientemente (Rojas et ál. 2010). Aunque los registros históricos sugieren una importante migración de Sevilla y Andalucía (España) a Antioquia (Boyd-Bowman 1976), se ha sugerido que durante la época de la Conquista llegó un grupo considerable de vascos (Twinam 1980) y judíos conversos (Mesa 1989); por lo tanto, nuestros resultados formulan de nuevo la pregunta acerca de su llegada a la región.
Nuestros hallazgos pueden tener implicaciones médicas, ya que indican que la MZI y Aranzazu comparten una composición y una diversidad genéticas disminuidas. Marinilla y Aranzazu podrían también compartir patrones epidemiológicos de enfermedades complejas y mendelianas (Jenny García, comunicación personal), y así aumentaría la posibilidad de que en ambas poblaciones puedan circular variantes genéticas comunes relacionadas con enfermedades humanas. Estos resultados destacan la importancia de identificar enfermedades en estas poblaciones y emprender estudios de mapeo genético de potenciales genes causales.
CONCLUSIONES
En este estudio se comprueba la complejidad de la coancestría entre apellidos y linajes genéticos del cromosoma Y en las poblaciones humanas de Colombia. Además, pese al origen polifilético y la tasa variable de no paternidad de apellidos comunes en poblaciones colombianas, se demuestra la utilidad del análisis simultáneo de apellidos y linajes del cromosoma Y para inferir procesos poblacionales.
AGRADECIMIENTOS
Los autores agradecen a todos los voluntarios que participaron en este estudio, así como a las entidades que contribuyeron en la colección de muestras. La financiación para este estudio fue aportada por el Comité para el Desarrollo de la Investigación (CODI) de la Universidad de Antioquia, Sostenibilidad de Grupos 2009-2011.
REFERENCIAS
1. ÁLVAREZ, VÍCTOR M. 1996. "Poblamiento y población en el valle de Aburrá y Medellín, 1541-1951". En Historia de Medellin, editado por Jorge Orlando Melo, 57-84. Medellín: Suramericana de Seguros. [ Links ]
2. ARANGO-MEJÍA, GABRIEL. 1993. Genealogías de Antioquia y Caldas, 3.ª ed. Medellín: Litoarte. [ Links ]
3. BANDELT, HANS J., PETER FORSTER Y ARNE ROHL. 1999. "Median-joining Networks for Inferring Intraspecific Phylogenies". Molecular Biology and Evolution 16 (1): 37-48. [ Links ]
4. BARRAI, ITALO, ÁLVARO RODRÍGUEZ-LARRALDE, ELISABETTA MAMOLINI, FRANZ MANNI Y CHIARA SCAPOLI. 2000. "Elements of the Surname Structure of Austria". Annals of Human Biology 27 (6): 607-622. [ Links ]
5. BARRAI, ITALO, ÁLVARO RODRÍGUEZ-LARRALDE, ELISABETTA MAMOLINI, FRANZ MANNI Y CHIARA SCAPOLI. 2001. "Isonymy Structure of USA Population". American Journal of Physical Anthropology 114 (2): 109-123. [ Links ]
6. BARRAI, ITALO, CHIARA SCAPOLI, MARÍA BERETTA, CARLO NESTI, ELISABETTA MAMOLINI Y ÁLVARO RODRÍGUEZ-LARRALDE. 1996. "Isonymy and the Genetic Structure of Switzerland. i. The Distributions of Surnames". Annals of Human Biology 23 (6): 431-455. [ Links ]
7. BEDOYA, GABRIEL, JENNY GARCÍA, PATRICIA MONTOYA, WINSTON ROJAS, MARÍA AMÉZQUITA, IVÁN SOTO, MARÍA LÓPEZ, JORGE OSPINA-DUQUE Y ANDRÉS RUIZ-LINARES. 2006. "Análisis de isonimia entre dos poblaciones del noroeste de Colombia". Biomédica 26 (4): 538-545. [ Links ]
8. BEDOYA, GABRIEL, PATRICIA MONTOYA, JENNY GARCÍA, IVÁN SOTO, STEPHANE BOURGEOIS, LUIS CARVAJAL, DAMIAN LABUDA, VÍCTOR ÁLVAREZ, JORGE OSPINA, PHILIP HEDRICK Y ANDRÉS RUIZ-LINARES. 2006. "Admixture Dynamics in Hispanics: A Shift in the Nuclear Genetic Ancestry of a South American Population Isolate". Proceedings of the National Academy of Sciences USA 103 (19): 7234-7239. [ Links ]
9. BOWDEN, GEORGINA, PATRICIA BALARESQUE, TURI KING, ZIFF HANSEN, ANDREW LEE, GILES PERG L-WILSON, EMMA HURLEY, STEPHEN ROBERTS, PATRICK WAITE, JUDITH JESCH, ABIGAIL JONES, MARK THOMAS, STEPHEN HARDING Y MARK JOBLING. 2008. "Excavating Past Population Structures by Surnamebased Sampling: The Genetic Legacy of the Vikings in Northwest England". Molecular Biology and Evolution 25 (2): 301-309. [ Links ]
10. BOYD-BOWMAN, PETER. 1976. "Patterns of Spanish Emigration to the Indies until 1600". The Hispanic American Historical Review 56 (4): 580-604. [ Links ]
11. CABRAL, RITA, CLAUDIA BRANCO, SONIA COSTA, GIANUMBERTO CARAVELLO, MIRO TASSO, BERNARDO PEIXOTO Y LUISA MOTA-VIEIRA. 2005. "Geography of Surnames in the Azores: Specificity and Spatial Distribution Analysis". American Journal of Human Biology 17 (5): 634-645. [ Links ]
12. CARVAJAL-CARMONA, LUIS, IVÁN SOTO, NICOLÁS PINEDA, DANIEL ORTIZ-BARRIENTOS, CONSTANZA DUQUE, JORGE OSPINA-DUQUE, MARK MCCARTHY, PATRICIA MONTOYA, VÍCTOR ÁLVAREZ, GABRIEL BEDOYA Y ANDRÉS RUIZ-LINARES. 2000. "Strong Amerind/white Sex Bias and a Possible Sephardic Contribution among the Founders of a Population in Northwest Colombia". American Journal of Human Genetic 67 (5): 1287-1295. [ Links ]
13. CASTRO DE GUERRA, DINORAH, HENRY ARVELO Y JUAN PINTO-CISTERNAS. 1999. "Population Structure of Two Black Venezuelan Populations Studied through Their Mating Structure and Other Related Variables". Annals of Human Biology 26 (2): 141-150. [ Links ]
14. CAVALLI-SFORZA, LUIGI Y ANTHONY EDWARDS. 1967. "Phylogenetic Analysis. Models and Estimation Procedures". American Journal of Human Genetic 19 (3, parte 1): 233-257. [ Links ]
15. DI GIACOMO, FABIO, FRANCESCA LUCA, LUIS POPA, NEJAT AKAR, NICHOLAS ANAGNOU, JURAJ BANYKO, RADIM BRDICKA, GUIDO BARBUJANI, FRANCO PAPOLA, GIOVANNI CIAVARELLA, FRANCESCO CUCCI, LEANDRO DI STASI, LUCIAN GAVRILA, MARINA KERIMOVA, DIMITAR KOVATCHEV, ANDREY KOZLOV, APHRODITI LOUTRADIS, VINCENZO MANDARINO, CORRADO MAMMI, EMMANUEL MICHALODIMITRAKIS, GIORGIO PAOLI, KALLIOPI PAPPA, GIULIANA PEDICINI, LUCIANO TERRENATO, SERGIO TOFANELLI, PATRIZA MALASPINA Y ANDREA NOVELLETTO. 2004. "Y Chromosomal Haplogroup J as a Signature of the Postneolithic Colonization of Europe". Human Genetics 115 (5): 357-371. [ Links ]
16. DIPIERRI, JOSÉ, EMMA ALFARO, CHIARA SCAPOLI, ELISABETTA MAMOLINI, ÁLVARO RODRÍGUEZ-LARRALDE E ITALO BARRAI. 2005. "Surnames in Argentina: A Population Study through Isonymy". American Journal of Physical Anthropology 128 (1): 199-209. [ Links ]
17. EXCOFFIER, LAURENT, GUILLAUME LAVAL Y STEFAN SCHNEIDER. 2005. "Arlequin (version 3.0): An Integrated Software Package for Population Genetics Data Analysis". Evolutionary Bioinformatics Online 1: 47-50. [ Links ]
18. FLORES, CARLOS, NICOLE MACA-MEYER, ANA GONZÁLEZ, PETER OEFNER, PEIDONG SHEN, JOSÉ PÉREZ, ANTONIO ROJAS, JOSÉ LARRUGA Y PETER UNDERHILL. 2004. "Reduced Genetic Structure of the Iberian Peninsula Revealed by Y-chromosome Analysis: Implications for Population Demography". European Journal of Human Genetics 12 (10): 855-863. [ Links ]
19. GONZÁLEZ-MARTÍN ANTONIO, CLARA GARCÍA-MORO, MIGUEL HERNÁNDEZ Y PEDRO MORAL. 2006. "Inbreeding and Surnames: a Projection into Easter Island's Past". American Journal of Physical Anthropology 129 (3): 435-445. [ Links ]
20. JOBLING, MARK. 2001. "In the Name of the Father: Surnames and Genetics". Trends in Genetics 17 (6): 353-357. [ Links ]
21. KARAFET, TATIANA, FERNANDO MÉNDEZ, MÓNICA MEILERMAN, PETER UNDERHILL, STEPHEN ZEGURA Y MICHAEL HAMMER. 2008. "New Binary Polymorphisms Reshape and Increase Resolution of the Human Y Chromosomal Haplogroup Tree". Genome Research 18 (5): 830-838. [ Links ]
22. KIMURA, MOTOO Y TOMOKO OHTA. 1978. "Stepwise Mutation Model and Distribution of Allelic Frequencies in a Finite Population". Proceedings of the National Academic of Sciences USA 75 (6): 2868-2872. [ Links ]
23. KING, TURI Y MARK JOBLING. 2009a. "Founders, Drift, And Infidelity: The Relationship between Y Chromosome Diversity and Patrilineal Surnames". Molecular Biology and Evolution 26 (5): 1093-1102. [ Links ]
24. KING, TURI Y MARK JOBLING. 2009b. "What's in a Name? Y Chromosomes, Surnames and the Genetic Genealogy Revolution". Trends in Genetics 25 (8): 351-360. [ Links ]
25. MACINTYRE, SALLY Y ANNE SOOMAN. 1991. "Non-paternity and Prenatal Genetic Screening". Lancet 338 (8771): 869-871. [ Links ]
26. MESA, DANIEL. 1989. "Los judíos en el descubrimiento de America". Repertorio histórico de la Academia Antioqueña de Historia 38 (252): 1-20. [ Links ]
27. PARSONS, JAMES. 1968. Antioqueño Colonization in Western Colombia. Berkeley: University of California Press. [ Links ]
28. PIAZZA, ALBERTO, SABINA RENDINE, GIANNA ZEI, ANTONIO MORONI Y LUIGI CAVALLI-SFORZA. 1987. "Migration Rates of Human Populations from Surname Distributions". Nature 329 (6141): 714-716. [ Links ]
29. RODRÍGUEZ-LARRALDE, ÁLVARO, ITALO BARRAI Y J. C. ALFONZO MORENO. 1993. "Isonymy Structure of Four Venezuelan States". Annals of Human Biology 20 (2): 131-145. [ Links ]
30. RODRÍGUEZ-LARRALDE, ÁLVARO, G. FORMICA, CHIARA SCAPOLI, MARÍA BERETTA, ELISABETTA MAMOLINI E ITALO BARRAI. 1993. "Microevolution in Perugia: Isonymy 1890-1990". Annals of Human Biology 20 (3): 261-274. [ Links ]
31. ROJAS, WINSTON, MARÍA VICTORIA PARRA, OMER CAMPO, MARÍA ANTONIETA CARO, JUAN GUILLERMO LOPERA, WILLIAM ARIAS, CONSTANZA DUQUE, ANDRÉS NARANJO, JAIR GARCÍA, CANDELARIA VERGARA, JAIME LOPERA, ÉRICK HERNÁNDEZ, ANA VALENCIA, YURI CAICEDO, MAURICIO CUARTAS, JAVIER GUTIÉRREZ, SERGIO LÓPEZ, ANDRÉS RUIZ-LINARES Y GABRIEL BEDOYA. 2010. "Genetic Make Up and Structure of Colombian Populations by Means of Uniparental and Biparental DNA Markers". American Journal of Physical Anthropology 143 (1): 13-20. [ Links ]
32. SOTO, IVÁN. 2003. "Evaluación histórica y genética del origen de Marinilla y su zona de influencia". Tesis, Instituto de Biología, Universidad de Antioquia, Medellín. [ Links ]
33. SYKES, BRYAN Y CATHERINE IRVEN. 2000. "Surnames and the Y Chromosome". American Journal of Human Genetic 66 (4): 1417-1419. [ Links ]
34. THOMAS, MARK, NEIL BRADMAN Y HELEN M. FLINN. 1999. "High Throughput Analysis of 10 Microsatellite and 11 Diallelic Polymorphisms on the Human Y-Chromosome". Human Genetics 105 (6): 577-581. [ Links ]
35. TWINAM, ANN. 1980. "From Jew to Basque: Ethnic Myths and Antioqueño Entrepreneurship". Journal of Interamerican Studies and World Affairs 22 (1): 81-107. [ Links ]
36. ZAPATA-CUÉNCAR, HERIBERTO. 1941. Monografías de Antioquia. Medellín: Cervecería Unión. [ Links ]
37. ZEI, GIANNA, ANTONELLA LISA, ORNELLA FIORANI, CHIARA MAGRI, LUIS QUINTANA- MURCI, ORNELLA SEMINO Y AUGUSTA SANTACHIARA-BENERECETTI. 2003. "From Surnames to the History of Y Chromosomes: The Sardinian Population as a Paradigm". European Journal of Human Genetics 11 (10): 802-807. [ Links ]
ARCHIVOS SUPLEMENTARIOS
TABLA SUPLEMENTARIA 1. FRECUENCIA DE HAPLOTIPOS DE SEIS LOCI EN CADA MUESTRA
TABLA SUPLEMENTARIA 2. FRECUENCIA DE APELLIDOS
