










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Antropología
versão impressa ISSN 0486-6525
Rev. colomb. antropol. vol.51 no.2 Bogotá jul./dez. 2015
Variación métrica y morfológica de la población prehispánica de Colombia. Implicaciones para la problemática del poblamiento temprano de América
Metric and Morphological Variation of the Precolumbian Populations of Colombia. About the Early Peopling of America
José Vicente Rodríguez
Universidad Nacional de Colombia, Colombia
jvrodriguezc@unal.edu.co
Clemencia Vargas
Universidad Nacional de Colombia, Colombia
mcvargasv@unal.edu.co
Recibido; 4 de Febrero de 2015 Aprobado: 24 de Agosto de 2015
Resumen
La morfología craneal de los paleoamericanos del Nuevo Mundo se caracteriza por la dolico-hipsicefalia y dientes grandes, mientras que los amerindios (indígenas tardíos) poseen una bóveda craneal más ancha y dientes más pequeños. Esta dicotomía se ha interpretado mediante dos hipótesis: a) como resultado de dos oleadas migratorias, la más antigua desde el sureste de Asia, de tipo australomelanesio y la segunda, más tardía, de origen mongoloide; b) como resultado de la diferenciación genética regional. Mediante un análisis multivariado de variables craneométricas, odontométricas y morfológicas dentales y craneales, se estudian diecinueve muestras prehispánicas de Colombia, con resultados convergentes.
Palabras clave: Andes orientales, Colombia, craneometría, rasgos no métricos, odontometría, morfología dental.
Abstract
The morphology of early paleoamericans of the New World is characterized by a long, narrow and height cranial vault, and a big size of teeth, whereas the amerinds (more recent indians) exhibit a shorter and wider cranial vault and more little teeth. Two hypotheses have been proposed: (a) As the result of two different migrations, in the final Pleistocene from Southeast of Asia by population remarkable similar to modern Australo-Melanesian, and by a later Holocene migration from Northeast Asia with mongoloid morphological pattern; (b) by the result of local genetic diversification. We evaluated these hypotheses by the multivariate analysis of craniometrical, odontometrical and morphological traits from skull and teeth of 19 pre-Columbian groups of Colombia. Both quantitative craniometrical and odontometrical techniques generated convergent results.
Keywords: Eastern Andes, Colombia; craniometry, nonmetric traits, odontometry, dental morphological traits.
Introducción
Las poblaciones aborígenes del Nuevo Mundo varían ampliamente desde el punto de vista craneométrico (Castillo 1981; Cocilovo y Guichón 2000; González et al. 2003, 2008; Howells 1989; Hubbe, Harvati y Neves 2011; Neves, Hubbe y Correal 2007; Powell y Neves 1999; Powell et al. 1999; Pucciarelli 2004; Rodríguez 2007; Rothhammer y Silva 1990), morfológico-craneal (rasgos discretos) (Conner 1990; Sciulli 1990), morfológico-dental (Cucina et al. 2003; Griffin, Lambert y Monahan 2001; Ha-ydenblit 1996; Pompa y Padilla 1990; Sutter 2005; Turner 1993; Turner y Scott 2006; Vargas 2010) y odontométrico (Kieser 1990; Perzigian 1976; Rodríguez y Vargas 2010; Scott 1979). Esta variación se puede agrupar, a grandes rasgos, en dos troncos morfométricos: 1) el paleoamericano, caracterizado por la dolico-hipsicefalia, robustez del aparato masticatorio, dientes grandes y mesomorfia facial; 2) el amerindio, que se destaca por la braquicefalia (con algunas excepciones, como Pericú en California y Botocudo en Brasil), aplanamiento facial (mongolización), más gracilidad y dientes más pequeños. Esta dicotomía se ha explicado mediante dos hipótesis: 1) la migracionista, que plantea dos oleadas migratorias distanciadas cronológica (paleoamericanos más tempranos) y espacialmente (paleoamericanos desde el sureste de Asia y amerindios provenientes desde el noreste de Asia) (Hubbe, Harvati y Neves 2011; Powell y Neves 1999; Powell et al. 1999; Pucciarelli, Pérez y Politis 2010); 2) la microevolutiva, cuyos autores sugieren que diferentes factores evolutivos (sistemáticos y aleatorios) y culturales condujeron a la reestructuración genética de la población originaria que penetró por Beringia (Chatters et al. 2014; Cocilovo et al. 2001; Cocilovo y Guichón 2000; Manríquez et al. 2011; Rodríguez 2011; Rothhammer y Dillehay 2009).
Algunas de estas investigaciones adolecen de varios sesgos metodológicos: 1) no se incluye una visión integral que aborde la evolución craneofacial en el ámbito tanto morfológico como métrico, teniendo en cuenta que el comportamiento de estos rasgos puede ser independiente y reflejar distintos momentos y ritmos evolutivos; 2) no se vincula la amplia variación espacial (desde Alaska hasta la Patagonia) y temporal (desde los paleoamericanos hasta los más tardíos) de América y de regiones vecinas (Siberia, Australia-Melanesia y Oceanía). Por ejemplo, el interés por resaltar rasgos negroides conduce a comparar las muestras paleoamericanas básicamente con africanos (Neves et al. 2003; Neves, Hubbe y Correal 2007). Al respecto, hay que destacar que las principales tendencias evolutivas en el Nuevo Mundo se expresan en la braquicefalización, el aplanamiento facial (mongolización), la gracilización del aparato masticatorio y la reducción del tamaño dental, sin que se evidencien cambios sustanciales en la estructura genética (Chatters et al. 2014).
El gran interrogante es cuándo, dónde y por qué causas se presentaron estos cambios morfométricos. En este sentido, el territorio de la cordillera Oriental de Colombia, dada su estratégica ubicación en el noroeste de Suramérica, por donde penetraron los primeros pobladores, y la existencia de muestras craneales que dan cuenta de la variación temporal de casi 10.000 años en aproximadamente 30.000 km2, puede arrojar luces sobre estos interrogantes. Al respecto, se ha planteado que la distancia geográfica entre sitios arqueológicos explica gran parte de la variación craneométrica (Cocilovo y Guichón 2000, 17; Rothhammer y Silva 1990, 12) y morfológica (Sciulli 1990, 28). Sin embargo, poblaciones como los aleutianos-esquimales (noroccidente de América) demuestran una gran afinidad con los fueguinos (Tierra del Fuego), distanciados de aquellos por varios millares de kilómetros; por otro lado, grupos tardíos como pericúes (Baja California) y botocudos (Brasil) exhiben similitudes con los paleoamericanos, de los que están separados por varios millares de años, lo que indica que en la diferenciación biológica indígena actuaron fenómenos poblacionales complejos, como la deriva genética, el efecto de cuello de botella, el efecto de fundadores, el efecto de la fusión-fisión de los nuevos territorios, las migraciones tardías y el tipo de recursos alimenticios disponibles (Cocilovo y Guichón 2000, 24-25).
El objetivo del presente reporte es aportar información morfométrica al problema del tiempo, el espacio y los mecanismos de los cambios epigenéticos de la población prehispánica de los Andes septentrionales, y sus implicaciones en la problemática del poblamiento temprano de América. La existencia de diversos ambientes que oscilan entre la Mesa de los Santos, a 900 m s. n. m. (habitada antes de la llegada de los españoles por los guanes); la Sierra Nevada del Cocuy, que posee alturas superiores a los 5.000 m s. n. m. (habitada antiguamente por los laches); la Montaña Santandereana, caracterizada por elevaciones superiores a los 3.000 m s. n. m. (región de los antiguos chitareros); el altiplano cundiboyacense ubicado a 2.500 m s. n. m., habitado por muiscas de tres grandes confederaciones (Bogotá, Tunja y Sogamoso); los valles interandinos (ríos Cauca y Magdalena); la región noreste con sus costas; la península de La Guajira y la Serranía de Perijá, situadas en la frontera con Venezuela, y, finalmente, el altiplano nariñense, en la frontera con Ecuador, proporcionó diversos recursos y generó dinámicas poblacionales particulares que pueden arrojar luces sobre los procesos evolutivos prehispánicos, desde los primeros cazadores recolectores hasta los agroalfareros que encontraron los españoles en el siglo XVI.
Materiales y métodos
En esta investigación se incluyeron muestras de varias regiones y periodos de Colombia, algunas estadísticamente grandes y con adecuado contexto arqueológico (Boyacá, Valle), otras muy pequeñas (Huila, Quimbaya, Perijá, Nariño) (tabla 1). Las mediciones craneométricas siguieron la metodología y nomenclatura de R. Martin (Martin y Saller 1959), incluyendo 31 rasgos (9 de la bóveda craneal, 17 del esqueleto facial y 5 mandibulares) en la comparación de las muestras colombianas (tabla 2) y 16 rasgos en el análisis poblacional continental (tabla 4). En la comparación territorial se incluyeron bases de datos de América del Norte (Howells 1989; Newman 1957), Centroamérica (Romano 1970), Suramérica (Castillo 1981; Cocilovo y Guichón 2000; Neves, Hubbe y Correal 2007; Rodríguez 2007; Rothhammer y Silva 1990), Siberia (Alexeev y Gochman 1978), Australia-Melanesia (Howells 1989) y Oceanía (Hanihara 1996; Howells 1989).

En el ámbito morfológico craneal se incluyó una batería de 32 rasgos discretos (Sciulli 1990, 27) para la comparación regional colombiana, pero se redujo a 18 en la comparación territorial debido a la existencia de pocas publicaciones en este campo.
En el análisis dental se midieron los diámetros mesodistal (MD) y vestibulolingual (VL) de cada diente (maxilar y mandibular de ambos lados), con lo cual se calculó el área por clase de dientes (I, C, P, M) y total (total size), siguiendo la metodología de Kieser (1990) y las bases de datos comparativas de Kieser (1990), Vargas (2010) y Perzigian (1976).
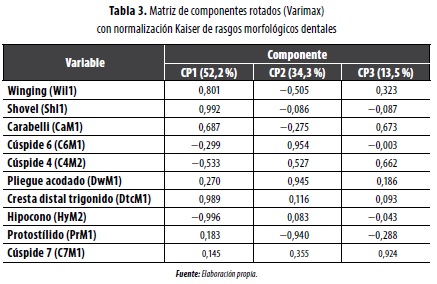
En la evaluación morfológica dental se observaron 10 rasgos (tabla 3), siguiendo la metodología de Turner (1993) y las bases de datos de Turner (1993) y Vargas (2010). Estas variables se pueden ver alteradas por el desgaste dental, especialmente de los incisivos centrales superiores, lo que afecta la observación de la forma en pala, que quizá generó el subregistro observado en muestras mexicanas (Haydenblit 1996) y que caracteriza a todas las poblaciones indígenas de América, por lo que es de poca utilidad en las comparaciones intergrupales (Pompa y Padilla 1990). En los molares el desgaste interproximal acorta el diámetro MD; de ahí que el VL sea más preciso, aunque menos dimórfico (Vargas 2010).
Análisis morfométrico y estadístico
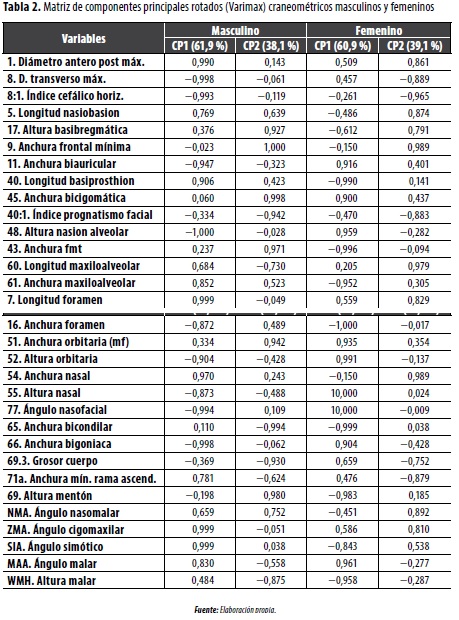
En el análisis craneométrico1 se pudo establecer que las longitudes (1, 5, 40) y anchuras (8, 9, 11, 45, 61, 66) se correlacionan significativamente entre sí (en el nivel 0,01), mientras que algunos segmentos constituyen variables más independientes, como el agujero occipital, las órbitas, la nariz, los ángulos faciales (nasofacial, nasomalar, cigomaxilar, simótico), la longitud maxiloalveolar y la mandíbula (especialmente la anchura bigoniaca que se relaciona solamente con la bicondilar) (Rodríguez 2001, 2007).
En la región caribe, la bóveda craneal es muy baja y gruesa de manera natural, tal como lo había anotado Gonzalo Fernández de Oviedo ([1526] 1996). Entre los muiscas de Tunja (Boyacá), se observa un tipo de deformación cefálica fronto-occipital convergente que la alarga (Rodríguez 2001, 262). En las muestras paleoamericanas y en el Herrera Temprano, la forma predominante es la dolico-hipsicefalia, que se extiende cronológicamente hasta el siglo II a. C., y en Chita (Boyacá), hasta el siglo IV d. C. (Rodríguez 2001, 2007). En este lapso de tiempo se aprecia una amplia variación en la forma de la bóveda craneal, que oscila entre la dolico-mesocefalia (Madrid UE-0) y la braquicefalia (Madrid UE-01).
El análisis de componentes principales proporciona una información muy útil de las relaciones entre las variables y las unidades, indicando tendencias en los datos originales y las variables relacionadas con dichas tendencias y comprimiendo las variables que no están relacionadas entre sí en un número menor (Shennan 1992, 261). La rotación Varimax mediante la normalización Kaiser y puntuaciones Bartlett permite acentuar las diferencias entre los grupos.
Por su parte, el análisis de conglomerados jerárquicos hace posible definir grupos afines en un conjunto de datos, en el caso nuestro, empleando la distancia euclídea al cuadrado (previa estandarización de las medidas mediante puntuaciones estándar en todos los análisis), la técnica aglomerativa de Ward (distancias de todos los puntos a las medias de los conglomerados a los que pertenecen, de modo que ellos serán lo más homogéneos posible) (Shennan 1992, 220). Aquí se excluyeron las variables craneométricas más afectadas por la deformación cefálica (1, 8, 8:1, 17, 11, 65) y la pérdida antemortem de dientes (60).
Finalmente, con el fin de observar las diferencias o similitudes temporales y espaciales de las muestras de los Andes orientales, en el ámbito geográfico de América, Siberia, Australia-Melanesia y Oceanía, se aplicó un análisis discriminante. Este divide las observaciones en grupos según unos criterios con la intención de encontrar la forma de distinguir dichos grupos a partir de las variables originales, indicando la contribución de cada una de estas en cada función (Shennan 1992, 285).
En el análisis de componentes principales craneométricos (figura 1), para el CP1 masculino, que explica el 61,9 % del total de la variación, las variables con mayor peso son las longitudes (1, 5, 40, 60, 7), varias anchuras (8, 11, 16, 61, 54, 66, 71a), ángulos faciales (ZMA, SIA, MAA) y alturas (48, 52). En el CP2 (38,1 °%) se incluye el resto de anchuras (9, 45, 43, 65, 60,3) y alturas (17, 69, WMH). En relación con el grupo femenino (figura 2), cuya información es más sesgada debido al tamaño pequeño de las muestras, en el CP1 (60,9 °%) participan básicamente anchuras (11, 45, 43, 61, 16, 51, 65, 66), alturas (52, 55, 69, WMH) y ángulos (SIA, MAA). En el CP2 se vinculan las longitudes (1, 5, 60, 7), algunas anchuras (8, 54, 71a) y ángulos (NMA, ZMA) (tabla 2, figuras 1 y 2). Este comportamiento de las variables significa que, al igual que para diferenciar el sexo, las anchuras constituyen los segmentos que mejor discriminan los grupos poblacionales prehispánicos, fenómeno relacionado con el ensanchamiento tanto de la bóveda craneal como del rostro, conocido como mongolización de los grupos tardíos.
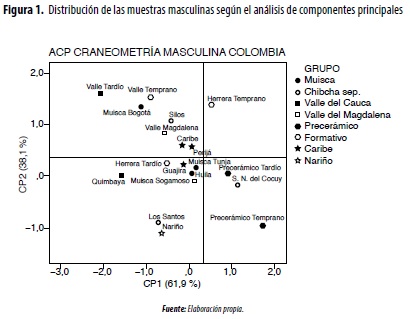
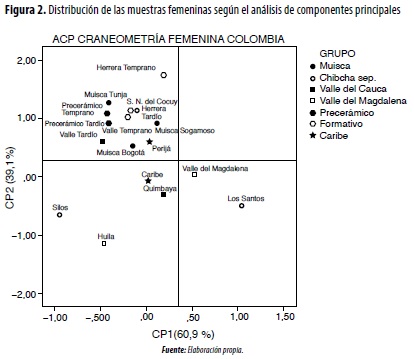
En las comparaciones intergrupales masculinas mediante el análisis de conglomerados jerárquicos, se conforman dos grandes enjambres, y el grupo Quimbaya ocupa una posición intermedia (figura 3): 1) andino, que vincula las muestras más antiguas (precerámicas) y tardías (muiscas de Tunja y Sogamoso, Silos, Sierra Nevada del Cocuy, Los Santos y Nariño); 2) valles interandinos (Cauca y Magdalena), donde además se incluye al Muisca de Bogotá y el Herrera (Temprano y Tardío). En la comparación de las muestras femeninas continúa la tendencia de aproximación entre los grupos del Valle del Cauca (Temprano y Tardío) y los andinos, especialmente del Temprano y el Herrera (figura 4).
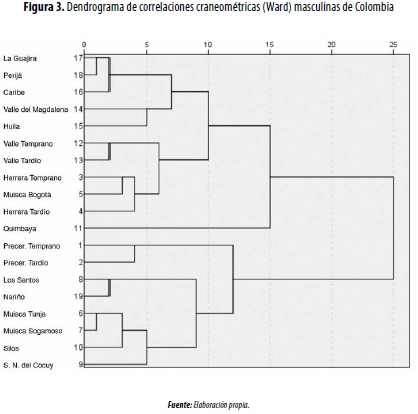
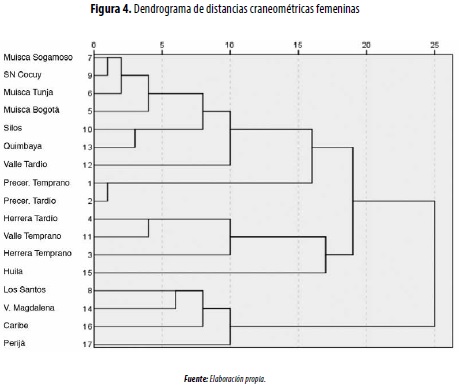
Estas similitudes fenotípicas podrían estar indicando una influencia de los valles interandinos en la configuración poblacional de los muiscas sureños (Bogotá), especialmente desde el periodo Formativo (Herrera).
Los rasgos discretos del cráneo han sido considerados de alto valor por su penetración genética; presentan poco dimorfismo sexual, por lo que ambos sexos se pueden integrar (Sciulli 1990). Infortunadamente, son pocas las publicaciones que incluyen esta batería de rasgos. En el análisis comparativo de las muestras colombianas mediante la prueba Kruskal-Wallis, las diferencias entre los grupos no son significativas (significación asintótica de 0,450, nivel de significancia de 0,05, por lo que se conserva la hipótesis nula). Mediante el análisis de conglomerados jerárquicos se conforman dos grandes enjambres: 1) andinos, que incluye todos los grupos tardíos de la cordillera Oriental; 2) valles interandinos, que vincula los grupos del Valle del Cauca y Magdalena, además de los precerámicos. Sin embargo, en la matriz de proximidades, estos últimos presentan las menores distancias con los grupos tardíos de la cordillera Oriental.
En el análisis de componentes principales de los rasgos discretos se configuran tres CP, de los cuales el CP1 explica el 42,8 °% de la variación e incluye los huesecillos adicionales o wormianos (ossicula) en lambda, bregma, sutura coronal y supramastoideo. El CP2 explica el 32,7 °% e integra los agujeros (foramen) ovale, espinoso, palatino, ethmoidal y los tubérculos (torus) en palatino y maxilar. El CP3 explica el 24,5 °%, con los huesecillos en la sutura lambdoidea y asterion. El grupo de Silos es el más diferente, pues todos los demás se integran en una gran agrupación.
De esta manera, estos rasgos no son muy útiles en los análisis intergrupales regionales, aunque sí sirven en las comparaciones continentales, pues separan tajantemente las muestras colombianas de las brasileñas, norteamericanas y siberianas, pero las integran con otras muestras suramericanas.
Los rasgos morfológicos dentales, al igual que los craneales, tampoco muestran diferencias significativas entre los grupos locales, particularmente en rasgos como los incisivos en pala (SHI1), pues el 100 °% de las muestras los poseen. En la comparación mediante conglomerados jerárquicos, al igual que los rasgos discretos, se diferencian los grupos andinos de los valles interandinos.
El análisis de componentes principales construye tres grandes componentes (tabla 3): el CP1, que explica el 52,2 % de la variación, incluye tres rasgos marcadamente mongoloides (Will, ShI1, DtcMl) y uno sin orientación específica (HyM2), y separa a los grupos de África, Australia-Melanesia, Europa y afro-colombianos contemporáneos (Cartagena, Guapi, Providencia, Tumaco), por un lado, de los de América, Siberia, Amur, Japón, Mongolia, Vietnam y mestizos colombianos (Bogotá, Cali, Guatavita, Tunja), por otro; el CP2 (34,3 °%) incluye la cúspide 6 (C6M1) y el protostílido (PrMl); el CP3 (13,5 °%) tiene la cúspide 7 (C7M1, rasgo africano y de grupos precerámicos) como variable con el mayor peso. Los rasgos Carabelli (CaM1) y especialmente la cúspide 4 (C4M2, rasgo caucasoide) se ubican en los tres componentes con un peso similar.
Las dimensiones dentales constituyen una excelente fuente de información sobre las tendencias en la variación temporal y espacial de las poblaciones prehispánicas de los Andes septentrionales y de América en general. Podemos apreciar que el tamaño dental, a juzgar por el TS (total size) se ha reducido a partir de los cazadores recolectores, cuyo promedio en el Precerámico Temprano de Colombia (1.335 mm2) (Rodríguez y Vargas 2010) es similar al del Indian Knoll (1.333 mm2) (Perzigian 1976), grupo arcaico de Norteamérica. Este tamaño se reduce notablemente a 1.179 mm2 en el Precerámico Tardío, pero se incrementa a 1.239 mm2 en el periodo Herrera (Formativo) y se estabiliza en 1.250 mm2 en promedio en los grupos agroalfareros tardíos. Algunas muestras conservaron dientes un poco más grandes, como Valle Temprano (1.292 mm2) y Silos (1.283 mm2), pero son menores que los esquimales (1.322 mm2), cuya dentición es la más grande de América, y aún más pequeños que las muestras de Australia-Melanesia (1.448 mm2), que se caracterizan por ser macrodontes (Kieser 1990).
En el análisis de componentes principales de muestras de África, América, Asia, Australia-Melanesia y Oceanía, se conforman tres componentes: CP1 (51,3 % de la variación), que incluye las áreas de premolares inferiores y caninos (superiores e inferiores); CP2 (30,1 %), con los molares (superiores e inferiores) e incisivos (superiores e inferiores) con mayor peso; CP3 (18,6 °/0), que vincula los premolares superiores (figura 5).
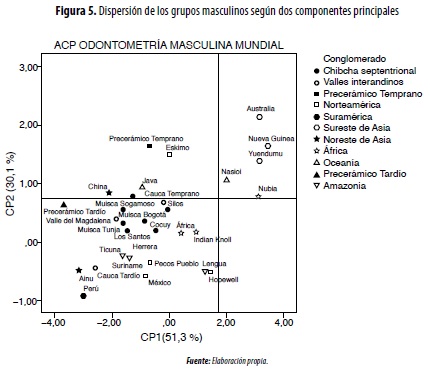
Por el CP1 se separan de manera significativa los grupos macrodontes del mundo como Australia-Melanesia y África antigua (Nubia); por el CP2 se diferencian estos mismos macrodontes y los grupos con dentición grande de América (Precerámico, esquimales, Valle Temprano) y Asia (China, Java) (figura 5).
Con el fin de brindar una noción más amplia de las relaciones fenéticas de las poblaciones precolombinas, se aplicó un análisis discriminante entre varias regiones de América (norte, centro, sur), Siberia, Oceania y Australia-Melanesia.
El análisis conforma varias fórmulas discriminantes (lambda de Wilks con significado de 0,00 para las 13 funciones calculadas), la primera de las cuales explica el 35,6 % de la variación e incluye con mayor peso la anchura nasal; la segunda explica el 21,1 °% y vincula el diámetro anteroposterior máximo y el ángulo cigomaxilar; la tercera (11,0 °%) incluye la longitud nasiobasion. El sistema clasifica correctamente el 87,6 °% de los casos agrupados originalmente. Los grupos más diferentes son el Paleolitico y Australia-Melanesia, mientras que América, Siberia y Oceania se integran alrededor de un centroide. Las menores distancias Mahalonobis (F < 3,0 con sig. 0,0) se aprecian entre el grupo PaleoSur y el PaleoNorte, Amazonia, Orinoquia, Patagonia y Oceania (Chatters 2000, 307), y las mayores, con relación a Australia-Melanesia (F = 9,7). Llama la atención que el grupo Amazonia registra las menores distancias con todos los demás de América y es el que más se aproxima a Australia-Melanesia (F = 6,5) y Oceania (F = 2,5) (tabla 4).
De esta manera, la población precolombina de América, tanto precerámica como agroalfarera tardia, se diferencia significativamente, en cuanto a la morfologia dental (especialmente por la forma en pala de los incisivos, cúspide de Carabelli y protostílido), el tamaño de todos los dientes (I, C, P, M, TS) y la craneometria (sobre todo por la anchura nasal y el ángulo cigomaxilar, que se relaciona con el prognatismo facial), de los aborigenes de Australia-Melanesia, pero se aproxima a Siberia y Oceania.
Análisis y discusión
El análisis morfométrico y estadístico multivariado de 19 muestras prehispánicas de los Andes septentrionales evidencia que las variables métricas (craneometria y odontometria) poseen un mayor potencial discriminante que las morfológicas (craneales y dentales) en la evaluación de las diferencias y similitudes de los grupos locales. En las primeras, las anchuras que describen la bóveda craneal (rasgos 8, 9, 11, 16), el esqueleto facial (rasgos 45, 51, 54), la mandíbula (rasgos 65, 66, 71a) y los dientes (áreas totales TS) son más informativas que las longitudes y alturas, comportamiento que también se ha observado en la diferenciación sexual. Algunas variables morfológicas dentales son tan homogéneas en los indigenas americanos como los incisivos en pala, pues se expresan en un 100 °% de las muestras analizadas (los reportes con bajas frecuencias deben estar sesgados por la incidencia del desgaste dental), pero sirven para diferenciar los grupos mongoloides de los de origen europeo, africano y australomelanesio.
A grandes rasgos, el análisis métrico separa los grupos antiguos (Precerámico y Formativo) de los más recientes (agroalfareros), en cuanto a las dimensiones de la bóveda craneal (dolico-hipsicefalia), el esqueleto facial (rasgos mesomorfos), la mandíbula (más robusta) y los dientes (macrodoncia) que caracterizan a los grupos de cazadores recolectores (VIII-III milenios a. C.), horticultores (II milenio a. C.) y alfareros tempranos (I milenio a. C.) de la cordillera Oriental de Colombia. Este proceso evolutivo debe estar relacionado con los cambios ambientales que se produjeron entre el III y el II milenio a. C., que condujeron a un incremento en la temperatura y a la reducción de la pluviosidad (Hammen 1992). Esos mismos cambios indujeron una transformación en el patrón de subsistencia con la aparición de la horticultura de tubérculos de altura, datada en Aguazuque (Soacha, Cundinamarca) hacia el II milenio a. C. (Correal 1990), y del consumo de maíz a inicios del I milenio a. C. (Cárdenas 2002).
Espacialmente, el análisis morfométrico separa a los grupos andinos de aquellos de los valles interandinos, aunque en el Herrera (I milenio a. C. a I milenio d. C.) y el Muisca de Bogotá (siglos X-XVI d. C.) se percibe alguna influencia de estos últimos, sobre todo del Valle Temprano (siglos V a. C. a V d. C.). Esto podría estar relacionado con el hecho de haber compartido un tronco ancestral común, con grupos de cazadores recolectores que ascendieron por el valle del río Magdalena, desde donde se habría generado una escisión hacia la cordillera Oriental y otra hacia la cordillera Central, y de ahí hacia el valle del río Cauca. Al respecto, el registro arqueológico evidencia la influencia de la región del valle del Magdalena en sitios precerámicos tempranos (Tequendama) y tardíos (Aguazuque) (Correal 1990; Correal y Pinto 1983; Correal y Hammen 1977) y Herrera Temprano (Rodríguez y Cifuentes 2005), consistente en materia prima lítica (chert) y animales de tierras cálidas, en los sitios precerámicos, y entre los alfareros tempranos de cerámica (Cardale y Paepe 1990).
Este proceso diferencial de poblamiento de la cordillera Oriental de Colombia, desde el norte (posiblemente siguiendo la ruta de los ríos Sogamoso-Chicamocha u Opón) y desde el sur (por el río Bogotá u otra ruta), daría origen a las diferencias biológicas y culturales que se observan entre los muiscas del norte (Tunja, Sogamoso) y sur (Bogotá), en cuanto a las prácticas funerarias, los estilos cerámicos (Rodríguez 2011) y la lengua y costumbres anotadas por los cronistas (Simón [1621] 1981).
En el ámbito continental, los grupos de los Andes septentrionales se aproximan a los de los Andes meridionales (especialmente Perú), Amazonia (Brasil), Orinoquia (Venezuela), Circuncaribe (Cuba), Centroamérica (México) y suroccidente de Norteamérica (Pueblo). En América se conforman tres grandes conglomerados morfométricos: 1) circunártico, que incluye a los aleutianos-esquimales, grupos de la GNWC (atapasco, haida, tlingit) y fueguinos (ona, yamana, alacaluf), afines a los de Siberia (chukchi, evenk); 2) Norteamérica (noreste, sureste), con similitud a Oceanía; 3) Centro-Suramérica, que incluye el suroccidente de Norteamérica (Pueblo), Centroamérica, Circuncaribe, Andes, Orinoquia, Amazonia y Patagonia, con rasgos similares a las poblaciones de Siberia (aplanamiento facial) y Oceanía (mesomorfia). Vale la pena resaltar que ninguno de estos grupos registra proximidad morfométrica significativa, por las variables empleadas en la comparación dental y craneal, con el de Australia-Melanesia. Quizá el que más se aproxima por la bóveda craneal es el de la Amazonia.
Los estudios de ADNmt han demostrado igualmente que existe una estrecha relación genética entre los indígenas americanos y los de Siberia, a juzgar por la predominancia de los haplogrupos A2, B2, C1 y D1, donde el D4h3 se ha registrado en muestras del Holoceno Temprano tanto en Norteamérica como en Suramérica (Bolnick et al. 2014; Chatters et al. 2014; Díaz et al. 2014). Esto indica que los amerindios son descendientes de los paleoamericanos que provienen del noreste de Asia, y que lo más probable es que la ruta para el ingreso inicial desde la región de Beringia haya sido la costera por el Pacífico (Cui et al. 2013; Rasmus-sen et al. 2014). Sin embargo, se ha hallado en indígenas de la Amazonia (Surui) una proporción muy baja (1-2 %) de genes de origen de la denominada Población Y (Australia, Nueva Guinea, Islas Andaman), procedente posiblemente de un tronco antiguo que compartieron paleoamericanos y australomelanesios y que arribó a América en uno de los "pulsos migratorios" tempranos (Balter 2015, 355). Al respecto, se ha planteado que el tronco ancestral que dio origen a los paleoamericanos surgió de una población protomongoloide, no especializada, mesomorfa, o mezclada con rasgos caucasoides (Kozintsev, Gromov y Moiseyev 1999, 194).
Conclusiones
El análisis morfométrico de muestras de los Andes orientales de Colombia evidencia un cambio significativo hacia el II milenio a. C., consistente en la reducción del aparato masticatorio (reducción del tamaño dental, de la anchura de la rama ascendente, del grosor del cuerpo mandibular). Esta tendencia se incrementa durante el periodo Herrera Temprano (I milenio a. C.), que se relaciona con cambios drásticos en el clima (reducción de la pluviosidad e incremento de la temperatura) y patrón de subsistencia (desarrollo de la horticultura de tubérculos de altura).
Ya para el Herrera Tardío (I milenio d. C.), con una población agroalfarera dependiente del maíz, se observa una marcada braquicefalización y gracilización del aparato masticatorio, tendencia que se extiende hasta la llegada de los españoles (área de muiscas, guanes, laches y chitareros). Este periodo es el de mayor variación poblacional, pues el índice cefálico oscila entre la dolicocefalia (Chita, Boyacá, siglo IV d. C.) y la braquicefalia (generalizada en los grupos tardíos), y no hay evidencias en el registro arqueológico de migraciones masivas tardías a los Andes orientales, como se había sugerido anteriormente (Lleras 1995). Resaltan los rasgos particulares del grupo de la Mesa de los Santos, Santander (ocupada a la llegada de los españoles por los guanes), porque poseen la nariz más angosta (23,4 mm en varones), y el Muisca de Tunja, con el rostro más angosto (135,6 mm en masculinos) y la bóveda craneal más alargada (índice cefálico de 78,9).
En el Valle del Cauca se observa una tendencia similar (aunque el análisis no se puede remontar a la etapa de cazadores recolectores por la ausencia de muestras), en la que el grupo Temprano (siglos V a. C. a V d. C.) es más robusto (con dientes más grandes e individuos dolicocéfalos) que el Tardío (siglos IX a XVI d. C.). Se registran similitudes morfométricas entre ambas regiones (Andes orientales y Valle del Cauca), que pueden estar relacionadas con un origen ancestral común.
Respecto a las otras regiones (valle del Magdalena, región Quimbaya, Orinoquia, Nariño), no es mucho lo que se puede afirmar debido al pequeño tamaño de las muestras. En estos grupos se aprecia la práctica cultural generalizada de la deformación cefálica, fenómeno que incide especialmente en la bóveda craneal produciendo hiperbraquicefalia. Su nariz es más ancha (26,7 mm), la región alveolar es más pronunciada y la mandíbula, más robusta.
En fin, en los Andes septentrionales se evidencia una amplia variabilidad biológica en el tiempo y el espacio, y la influencia de diferentes procesos evolutivos en la conformación de la estructura poblacional: inicialmente, las migraciones de cazadores recolectores que se remontaron desde el Caribe por el valle del río Magdalena hacia finales del Pleistoceno, se dispersaron por la región andina y los valles interandinos, dando lugar a distintos procesos adaptativos y estocásticos.
Con respecto a la problemática de los orígenes de la dualidad paleoamericana (dolico-hipsicefalia, mesomorfia facial, macrodoncia) / amerindia (braquicefalia, mongolización, microdoncia), se puede argüir que en los Andes septentrionales se generó por procesos microevolutivos, es decir, mediante reestructuración genética regional, influenciada por fenómenos ambientales y culturales complejos, diacrónicos y espacialmente diferentes.
Notas
1 Las medidas originales ocupan un amplio espacio, por lo que no se incluyen en este reporte, pero varias de ellas han sido publicadas originalmente (Rodríguez 2001, 2007; Rodríguez y Vargas 2010; Vargas 2010).
Referencias
Alexeev, V. P. e I. J. Gochman. 1978. Paleoanthropology of World and the Formation of Human Races. Moscú: Nauka. [ Links ]
Baiter, Michael. 2015. "New Mystery for Native American Origins". Science 349 (6246): 354-355. [ Links ]
Bolnick, Deborah A. et al. 2014. "Ancient DNA from Early to Mid-Holocene Burials in Northwestern Argentina: Implications for Understanding the Colonization and Early Population History of South America". American Journal of Physical Anthropology 153 (S58): 81. [ Links ]
Cardale, Marianne y Paul de Paepe. 1990. "Resultados de un estudio petrológico de cerámicas del periodo Herrera, provenientes de la sabana de Bogotá y sus implicaciones arqueológicas". Boletín del Museo del Oro 27: 99-119. [ Links ]
Cárdenas, Felipe. 2002. Datos sobre la alimentación prehispánica en la sabana de Bogotá, Colombia. Informes Arqueológicos 3. Bogotá: Instituto Colombiano de Antropología e Historia. [ Links ]
Castillo, Helia. 1981. "Craneología indígena de Venezuela". Estudios de Antropología Biológica 1: 263-327. [ Links ]
Chatters, James C. 2000. "The Recovery and First Analysis of an Early Holocene Human Skeleton from Kennewick, Washington". American Antiquity 65 (2): 291-316. [ Links ]
Chatters, James C. et al. 2014. "Late Pleistocene Human Skeleton and mtDNA Link Paleoamericans and Modern Native Americans". Science 344 (6185): 750-754. [ Links ]
Cocilovo José A. et al. 2001. "El proceso microevolutivo de la población nativa antigua de Arica". Chungara. Revista de Antropología Chilena 33: 13-20. [ Links ]
Cocilovo, José A. y Ricardo A. Guichón. 2000. "La variación geográfica y el proceso de microdiferenciación de las poblaciones aborígenes de Patagonia Austral y de Tierra del Fuego". Chungara. Revista Chilena de Antropología 15: 9-28. [ Links ]
Conner, Michael D. 1990. "Population Structure and Skeletal Variation in the Late Woodland of West-Central Illinois". American Journal of Physical Anthropology 82: 31-43. [ Links ]
Correal, Gonzalo. 1990. Aguazuque. Evidencias de cazadores, recolectores y plantadores en la altiplanicie de la cordillera Oriental. Bogotá: FIAN, Banco de la República. [ Links ]
Correal, Gonzalo y Thomas van der Hammen. 1977. Investigaciones arqueológicas en los abrigos rocosos del Tequendama. 12.000 años de historia del hombre y su medio ambiente en la altiplanicie de Bogotá. Bogotá: Biblioteca Banco Popular. [ Links ]
Correal, Gonzalo y María Pinto. 1983. Investigaciones arqueológicas en el municipio de Zipacón, Cundinamarca. Bogotá: FIAN, Banco de la República. [ Links ]
Cucina, Andrea et al. 2003. "Las poblaciones caribeñas desde el tercer milenio a. C. a la conquista española: las filiaciones biológicas desde la perspectiva antropológica dental". Estudios de Antropología Biológica 11 (2): 913-927. [ Links ]
Cui, Yinqiu et al. 2013. "Ancient DNA Analysis of Mid-Holocene Individuals from the Northwest Coast of North America Reveals Different Evolutionary Paths for Mitogenomes". Plos One 8 (7): e66948. [ Links ]
Díaz, Marcela et al. 2014. "Molecular Characterization of Early to Mid-Holocene Skeletal Human Remains Belonging to Checua Culture from Colombia - South America". The 22nd Annual Meeting of the Society for Molecular Biology and Evolution (SMBE), 8-12 de junio de 2014. San Juan, Puerto Rico. http://www.smbe.org/annual/2014. [ Links ]
Fernández de Oviedo, Gonzalo. [1526] 1996. Sumario de la natural historia de las Indias. México: Fondo de Cultura Económica. [ Links ]
González, Rolando et al. 2003. "Craniometric Evidence for Paleoamerican Survival in Baja California". Nature 425: 62-65. [ Links ]
González, Rolando et al. 2008. "The Peopling of America: Craniofacial Shape Variation on a Continental Scale and its Interpretation from an Interdisciplinary View". American Journal of Physical Anthropology 137: 175-187. [ Links ]
Griffin, Mark C., Patricia M. Lambert y Elizabeth Monahan. 2001. "Biological Relationship and Population History of Native Peoples in Spanish Florida and the American Southeast". En Bioarchaeology of Spanish Florida. The Impact of Colonialism, editado por C. S. Larsen, 226-273. Gainesville: University Press of Florida. [ Links ]
Hammen, Thomas van der. 1992. Historia, ecología y vegetación. Bogotá: Corporación Araracuara; Fondo FEN Col.; Fondo Promoción de la Cultura. [ Links ]
Hanihara, Tsunehiko. 1996. "Comparison of Craniofacial Features of Major Human Groups". American Journal of Physical Anthropology 99: 389-412. [ Links ]
Haydenblit, Rebecca. 1996. "Dental Variation among Four Prehispanic Mexican Populations". American Journal of Physical Anthropology 100: 225-246. [ Links ]
Howells, William White. 1989. Craniometric Analysis in the Dispersion of Modern Man. Papers of the Peabody Museum of Archaeology and Ethnology. Cambridge: Harvard University. [ Links ]
Hubbe, Mark, Katerina Harvati y Walter Neves. 2011. "Paleoamerican Morphology in the Context of European and East Asian Late Pleistocene Variation: Implications for Human Dispersion into the New World". American Journal of Physical Anthropology 144: 442-453. [ Links ]
Kieser, Julius A. 1990. Human Adult Odontometrics. Cambridge: Cambridge University Press. [ Links ]
Kozintsev, A. G., A. V. Gromov y V. G. Moiseyev. 1999. "Collateral Relatives of American Indians among the Bronze Age Populations of Siberia". American Journal of Physical Anthropology 109 (2): 193-204. [ Links ]
Lleras, Roberto. 1995. "Diferentes oleadas de poblamiento en la prehistoria tardía de los Andes orientales". Boletín del Museo del Oro 38-39: 3-11. [ Links ]
Manríquez, Germán et al. 2011. "Morphometric and mtDNA Analysis of Archaic Skeletal Remains from Southwestern and South America". Chungara. Revista de Antropología Chilena 43: 283-292. [ Links ]
Martin, Rudolf y Karl Saller. 1959. Lehrbuch der Anthropologie, I & II. Stuttgart: Gustav Fisher Verlag. [ Links ]
Neves, Walter A. et al. 2003. "Early Holocene Human Skeletal Remains from Santana do Riacho, Brazil: Implications for the Settlement of the New World". Journal of Human Evolution 45: 19-42. [ Links ]
Neves, Walter A., Mark Hubbe y Gonzalo Correal. 2007. "Human Skeletal Remains from Sabana de Bogotá, Colombia: A Case of Paleoamerican Morphology Late Survival in South America?". American Journal of Physical Anthropology 133: 1080-1098. [ Links ]
Newman, Russell W. 1957. A Comparative Analysis of Prehistoric Skeletal Remains from the Lower Sacramento Valley. Washington: Smithsonian Institution. [ Links ]
Perzigian, Anthony J. 1976. "The Dentition of the Indian Knoll Skeletal Population: Odontometric and Cusp Number". American Journal of Physical Anthropology 44: 113-122. [ Links ]
Pompa y Padilla, José Antonio. 1990. Antropología dental. Aplicación en poblaciones prehispánicas. Serie Antropología Física. México: Instituto Nacional de Antropología e Historia. [ Links ]
Powell Joseph et al. 1999. "Afinidades biológicas extra-continentales de los dos esqueletos más antiguos de América: implicaciones para el poblamiento del Nuevo Mundo". Antropología Física Latinoamericana 2: 7-22. [ Links ]
Powell, Joseph F. y Walter A. Neves. 1999. "Craniofacial Morphology of the First Americans: Pattern and Process in the Peopling of the New World". Yearbook of Physical Anthroplogy 42: 153-158. [ Links ]
Pucciarelli, Héctor M. 2004. "Migraciones y variación craneofacial humana en América". Complutum 15: 225-247. [ Links ]
Pucciarelli, Héctor M., S. Iván Pérez y Gustavo G. Politis. 2010. "Early Holocene Human Remains from the Argentinean Pampas: Additional Evidence for Distinctive Cranial Morphology of Early South Americans". American Journal of Physical Anthropology 143: 298-305. [ Links ]
Rasmussen, Morten et al. 2014. "The Genome of a Late Pleistocene Human from a Clovis Burial Site in Western Montana". Nature 506: 225-229. [ Links ]
Rodríguez, José Vicente. 2001. "Craneometría de la población prehispánica de los Andes orientales de Colombia: diversidad, adaptación y etnogénesis. Implicaciones para el poblamiento americano". En Los chibchas. Adaptación y diversidad en los Andes orientales de Colombia, editado por José Vicente Rodríguez, 250-310. Bogotá: Universidad Nacional de Colombia; Colciencias. [ Links ]
__. 2007. "La diversidad poblacional de Colombia en el tiempo y el espacio: estudio craneométrico". Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 31 (120): 321-346. [ Links ]
__. 2011. Los chibchas: hijos del sol, la luna y los Andes. Orígenes de su diversidad. Bogotá: Universidad Nacional de Colombia; IDU. [ Links ]
Rodríguez, José Vicente y Arturo Cifuentes. 2005. "Un yacimiento formativo ritual en el entorno de la antigua laguna de La Herrera, Madrid, Cundinamarca". Maguaré 19: 103-131. [ Links ]
Rodríguez, José Vicente y Clemencia Vargas. 2010. "Evolución y tamaño dental en poblaciones humanas de Colombia". Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 34 (133): 423-439. [ Links ]
Romano, Arturo. 1970. "Preceramic Human Remains". Handbook of Middle American Indians, 22-34. Austin: University of Texas Press. [ Links ]
Rothhammer, Francisco y Tom D. Dillehay. 2009. "The Late Pleistocene Colonization of South America: An Interdisciplinary Perspective". Annals of Human Genetics 73: 540-549. [ Links ]
Rothhammer, Francisco y Claudio Silva. 1990. "Craniometrical Variation among South American Prehistoric Populations: Climatic, Chronological, and Geographic Contributions". American Journal of Physical Anthropology 82: 9-17. [ Links ]
Sciulli, Paul W. 1990. "Cranial Metric and Discrete Trait Variation and Biological Differentiation in the Terminal Late Archaic of Ohio: The Duff Site Cemetery". American Journal of Physical Anthropology 82: 19-29. [ Links ]
Scott, Eugenie C. 1979. "Increase of Tooth Size in Prehistoric Coastal Peru, 10.000 B. P.-1.000 BP." American Journal of Physical Anthropology 50: 251-258. [ Links ]
Shennan, Stephen. 1992. Arqueología cuantitativa. Barcelona: Crítica. [ Links ]
Simón, Pedro. (1621) 1981. Noticias historiales de las conquistas de Tierra Firme en las Indias Occidentales. Bogotá: Biblioteca Banco Popular. [ Links ]
Sutter, Richard. 2005. "The Prehistoric Peopling of South America as Inferred from Epigenetic Dental Traits". Andean Past 7: 183-217. [ Links ]
Turner, Christy G. 1993. "Southwest Indian Teeth". National Geographic Research & Exploration 9 (1): 32-53. [ Links ]
Turner, Christy G. y Scott Richard G. 2006. "The Dentition of American Indians: Evolutionary Results and Demographic Implications Following Colonization from Siberia". En Handbook of Paleoanthropology, editado por Winfried Henke e Ian Tattersall, 1901-1941. Springer-Verlag Press. [ Links ]
Vargas, María Clemencia. 2010. "Morfología y odontometría dental de poblaciones de Colombia". Tesis de doctorado, Facultad de Odontología, Universidad de Buenos Aires, Argentina. [ Links ]
