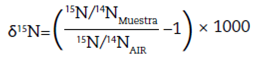Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
Las investigaciones arqueológicas en la microrregión de Antofagasta de la Sierra (ecorregión puna, provincia de Catamarca, noroeste argentino) han permitido proponer, con base en distintas líneas de evidencia, cambios en las estrategias económicas a lo largo del Holoceno Tardío. Al respecto, se postula la consolidación, a partir del ca. 2500 años a. P., de grupos agropastoriles que incrementaron progresivamente el cultivo de vegetales, pasando de estrategias intensivas a extensivas en la producción agrícola, proceso que se habría profundizado aún más con la llegada del Imperio incaico a la región (Olivera 1998). El presente trabajo tiene como objetivo establecer si los cambios económicos propuestos para la microrregión tuvieron un correlato en la composición de la dieta humana. En particular, nos preguntamos si el incremento y la posterior complejización de la producción agrícola postulados para alrededor de los 1000 años a. P. concuerdan con una disminución progresiva del componente cárnico en la dieta y con un aumento del consumo de maíz u otros recursos agrícolas característicos del desierto de altura (e. g., tubérculos y quínoa).
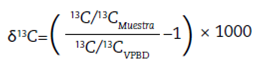
El análisis de isótopos estables del carbono (13C/12C) y nitrógeno (15N/14N) es una metodología ampliamente utilizada y adecuada para dar cuenta de esta problemática, pues permite conocer, por un lado, el patrón fotosintético dominante en la dieta asimilada y, por otro, el lugar que ocupan los individuos bajo análisis dentro de la cadena trófica1 (Ambrose 1993; De Niro y Epstein 1978). Con este fin realizamos un análisis paleodietario mediante el estudio de las relaciones isotópicas en el registro óseo humano, tanto en la fracción orgánica como en la inorgánica (δ13Cco, δ13Cap, δ15Ν) de individuos arqueológicos recuperados de diversos sitios de la microrregión. Este estudio generó nuevos datos, a la vez que interpretó valores isotópicos ya publicados en otras investigaciones (Araníbar et al. 2007; Babot et al. 2009). Asimismo, dada la importancia en la reconstrucción paleodietaria de las variables ambientales en la distribución de valores isotópicos en cada área, se realizaron análisis sobre una muestra de camélidos del área de estudio y se integraron los datos generados ya publicados para plantas cultivadas (Killian y Salminci 2014).
Área de estudio
La ecorregión puna del noroeste argentino (NOA) conforma el extremo sur del altiplano andino y ocupa porciones de las actuales provincias de Jujuy, Salta y Catamarca. Se emplaza entre 22° y 27° S y 65° 10' y 36° 50' O, con una altitud entre los ca. 3.000 y 4.700 m s. n. m. La ecorregión, en general, presenta una gran amplitud térmica, una intensa radiación solar debida a la altitud, una marcada estacionalidad y un sistema de precipitación estival. Las lluvias son a menudo escasas e impredecibles y pueden adquirir un carácter torrencial; son fuertemente influidas por las anomalías climáticas ENSO-El Niño (Tchilinguirian 2009). La distribución de los recursos no es homogénea y se distinguen áreas de alta concentración frente a otras donde se encuentran muy dispersos o no existen. Los recursos se concentran en los microambientes de fondos de cuenca y quebradas protegidas que cuentan con disponibilidad de agua relativamente estable.
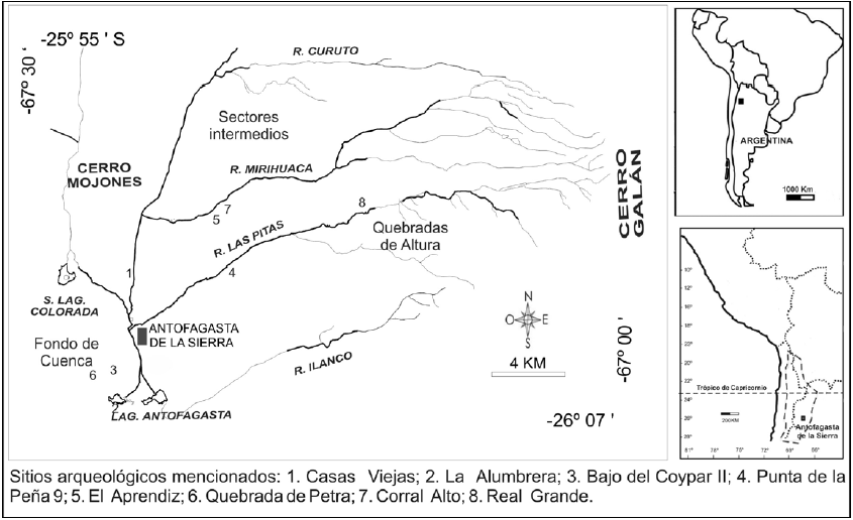
En el caso específico de Antofagasta de la Sierra (en adelante AS), la microrregión se caracteriza por tener un paisaje con alta heterogeneidad, en el que se distinguen tres microambientes con oferta diferencial de recursos (figura 1): 1) fondo de cuenca (3.400 -3.550 m s. n. m.), que tiene las mejores posibilidades para la agricultura en virtud de su topografía abierta y disponibilidad de agua; 2) sectores intermedios (3.550-3.800 m s. n. m.), que incluye los cursos inferiores y medios de los af luentes del río Punilla, con disponibilidad de forraje, agua y tierras aptas para la producción agropastoril, aunque con menos extensión que en el primer sector, e importantes extensiones de campo-tolar; 3) quebradas de altura (3.800-4.600 m s. n. m.), protegidas y estrechas en los cursos medios y superiores de los af luentes del Punilla, con agua permanente y forraje diverso, adecuadas para actividades de caza y pastoreo (Olivera y Podestá 1993), asociadas a los pajonales de altura con importante aporte de forraje. Las precipitaciones en la región tienen un promedio de 124 mm anuales (aunque en ciertos años son inexistentes en el fondo de cuenca), la temperatura media es de 10° C y la humedad relativa, del 30 %.
Antecedentes arqueológicos
A partir de ca. 5000 años a. P. (Arcaico Tardío) existen evidencias de procesos de circunscripción territorial en la región, disminución en la distancia de los movimientos logísticos y, quizá, aumento del sedentarismo. En este sentido, se identificaron bases residenciales con evidencias de actividades múltiples en cuevas y abrigos rocosos de los sectores intermedios y quebradas de altura de la cuenca (Aschero y Hocsman 2011, entre otros). Desde ca. 4500 años a. P. existen evidencias de un posible proceso local de domesticación de camélidos y el hallazgo de microrrestos correspondientes a cultígenos (quínoa, papa, oca), en contextos de cazadores-recolectores en transición a pastores en AS (Aschero y Hocsman 2011; Babot 2005). Hacia ca. 3000 años a. P., habrían empezado a consolidarse en la microrregión sociedades sedentarias con vida aldeana, estrategias mixtas agrícolas-pastoriles y tecnología cerámica (Olivera 1998).
Para estas nuevas sociedades agropastoriles se ha propuesto un modelo de sedentarismo dinámico, según el cual los grupos humanos habrían utilizado recursos de diversos microambientes en forma integrada, en un sistema de asentamiento-subsistencia que apuntaba a la disminución del riesgo a través de la ampliacion del espectro de recursos y la utilización de variadas estrategias (pastoreo, agricultura, caza, recolección e intercambio intra e interregional). Las poblaciones tendrían asentamientos bases en los que parte de sus integrantes permanecerían el año completo en los fondos de valle de las cuencas endorreicas o en quebradas protegidas, zonas aptas para la producción agrícola-pastoril y asentamientos de ocupación temporaria o semipermanente. Estos últimos se encontrarían ubicados en otros microambientes, a los que parte del grupo se trasladaría con una periodicidad variable, relacionada con el pastoreo, la caza y la extracción de otros recursos -por ejemplo, minerales o líticos- (Olivera 1998). Sin embargo, esto no implicó que no se produjera, con el paso del tiempo, un profundo proceso de cambio en las sociedades de la microrregión.
En un momento inicial (ca. 3000-1700 años a. P.) se sugirió un modo logístico de pastoreo con agricultura, puesto que el pastoreo era el principal eje económico productivo -para ca. 2400 años a. P. se registra la presencia de una alta proporción de camélidos de igual tamaño a la llama actual- complementado con una agricultura de pequeña escala. Por su parte, la caza de vicuñas fue un componente importante en la subsistencia de estos grupos y quizá el mayor aporte de proteína cárnica a su dieta (Olivera 1998; Olivera y Grant 2008).
Desde el comienzo de la era cristiana, la producción agrícola habría comenzado a tener mayor incidencia. Sin embargo, el pastoreo de camélidos continuó ocupando un papel esencial en la economía, en especial en los sectores intermedios de la cuenca. Este segundo momento se caracterizó por la presencia de una mayor influencia de grupos de los valles mesotermales más bajos, en particular, de Hualfín y Abaucán (provincia de Catamarca) (Olivera y Podestá 1993). En esta etapa se registró un aumento demográfico que coincide con una optimización en el uso de los espacios del fondo de cuenca y la ocupación más estable de los denominados sectores intermedios (cursos bajos y medios de las quebradas laterales) (Olivera y Vigliani 2000-2002). Los resultados osteométricos obtenidos2 de la comparación entre los componentes tempranos y tardíos de la aldea Casa Chávez Montículos (ca. 2400-1300 a. P.) muestran una disminución en la proporción de vicuñas a medida que pasa el tiempo, desde el componente más temprano hasta el más tardío. Esta tendencia ha sido confirmada por análisis estadísticos multivariados que permitieron asignar un 42 % de los especímenes medidos del componente inferior a vicuña, mientras que para el componente superior ese porcentaje se redujo a un 17 % (Olivera y Grant 2008).
Hacia ca. 1200-1000 años a. P., se infirió una intensificación y expansión de la producción agrícola y se aprovecharon los terrenos fértiles del piedemonte del cerro del Coypar, incorporando mejoras en la construcción de acequias y tras la delimitación de canales y parcelas (Tchilinguirian y Olivera 2000). Posteriormente, hacia ca. 700 años a. P., se observaron modificaciones en la organización social y política, de lo cual es paradigmático el sitio urbano La Alumbrera. Durante esta etapa, la importancia del cultivo en la economía se profundizaría, lo que se evidencia con la tecnología cerámica utilizada para el almacenaje (Olivera y Vigliani 2000-2002), la numerosa cantidad de marlos de maíz hallados en Quebrada de Petra (Olivera y Vigliani 2000-2002) y el mayor carácter expedito en la producción de los conjuntos líticos (Elías 2012). No obstante y respecto a esto último, los instrumentos asociados con la labranza no son frecuentes en los sitios adscritos a este periodo (Elías 2012). Asimismo, en los conjuntos faunísticos, continúa la importancia de Lama glama, pero se observa una persistencia, incluso el incremento, hasta momentos tardíos de las evidencias de caza (e. g., Vicugna vicugna). Finalmente, con la ocupación incaica se introdujeron cambios en la infraestructura de cultivo que dieron lugar a un sistema productivo agrícola aún más complejo (Tchilinguirian y Olivera 2000).
También se observan variaciones en las condiciones climáticas asociadas a los cambios organizacionales. Olivera y Tchilinguirian (2006) señalan que en la microrregión de Antofagasta de la Sierra se produce una fase húmeda entre ca. 3400-1600 a. P. con buenas condiciones en la cuenca. Las condiciones se tornan más áridas e inestables a partir de los 1650-1700 años a. P., lo que coincide con la optimización de la ocupación de los espacios en toda la cuenca y la incorporación de nuevas tecnologías para la agricultura de siembra y cosecha. Esta tendencia se habría profundizado durante el periodo de la Anomalía Climática Medieval (ca. 1000 a. P.), lo que se evidencia en la infraestructura agrícola de sitios como Bajo del Coypar I y Campo Cortaderas (Olivera y Tchilinguirian 2006).
Isótopos estables en la puna y en Antofagasta de la Sierra
Para la reconstrucción de la ecología isotópica de la ecorregión de puna en el NOA,hasta el momento contamos con estudios detallados sobre valores δ 13C en pasturas de la puna de Jujuy (Fernández y Panarello 1999-2001) y con los resultados obtenidos de investigaciones que han ampliado el conocimiento sobre camélidos en el área de Susques, también en la puna de Jujuy (Samec, Morales y Yacobaccio 2014; Yacobaccio, Morales y Samec 2009; Yacobaccio, Samec y Cata 2010). En cuanto a estos últimos, se ha podido identificar una correlación negativa entre la altitud y los valores δ13C en colágeno, tanto en llamas como en vicuñas actuales (Fernández y Panarello 1999-2001; Yacobaccio, Morales y Samec 2009; Yacobaccio, Samec y Cata 2010). Asimismo, se han presentado los primeros valores δ15N de Camelidae que muestran una correlación positiva entre dichos valores y la altitud durante el Holoceno Temprano y Medio, lo que se asocia con una mayor disponibilidad de humedad en las áreas más altas (Samec, Morales y Yacobaccio 2014).
Para la microrregión de AS resulta oportuno destacar el estudio de isótopos estables en camélidos para la transición entre el Holoceno Temprano y Medio (Mondini, Panarello y Zangrando 2010)3. Este estudio resultará relevante a la hora de discutir los datos del Holoceno Tardío, aunque no será integrado como insumo en el momento de modelar la estimación paleodietaria humana. También son de interés las mediciones isotópicas realizadas sobre pelo de camélido4 en el sitio punta de la Peña 9 (PP9), emplazado en sectores intermedios sobre la margen sur del río Las Pitas (Araníbar et al. 2007).
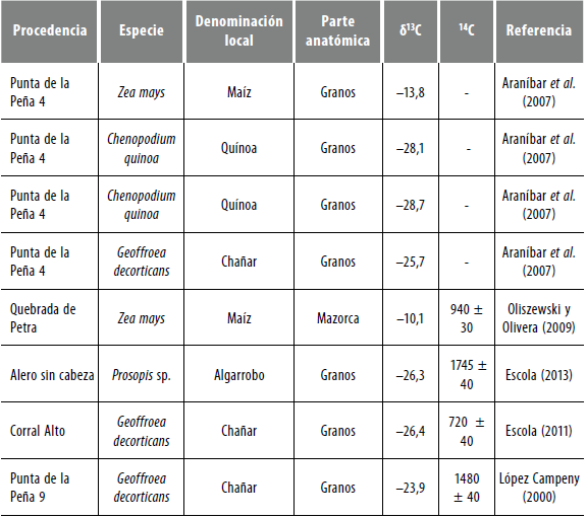
Por su parte, en cuanto a los valores isotópicos de vegetales, se cuenta con un estudio sobre recursos vegetales de consumo humano directo (quínoa, maíz y papa) realizado en AS por Killian y Salminci (2014) (véase la síntesis del estudio en la tabla 1). Finalmente, otro recurso que aporta a la reconstrucción ecológica del área es la consideración de los valores δ13C de los ejemplares vegetales arqueológicos que han sido publicados previamente o que acompañaron fechados radiocarbónicos (Araníbar et al. 2007; Elías 2010; Escola et al. 2013; López Campeny 2000) (tabla 2).
Tabla 1 Valores isotópicos de recursos vegetales de consumo humano procedentes de Antofagasta de la Sierra
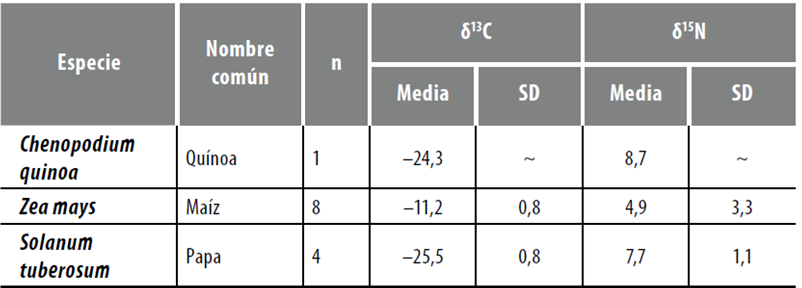
Fuente: Elaboración propia.
Materiales y métodos
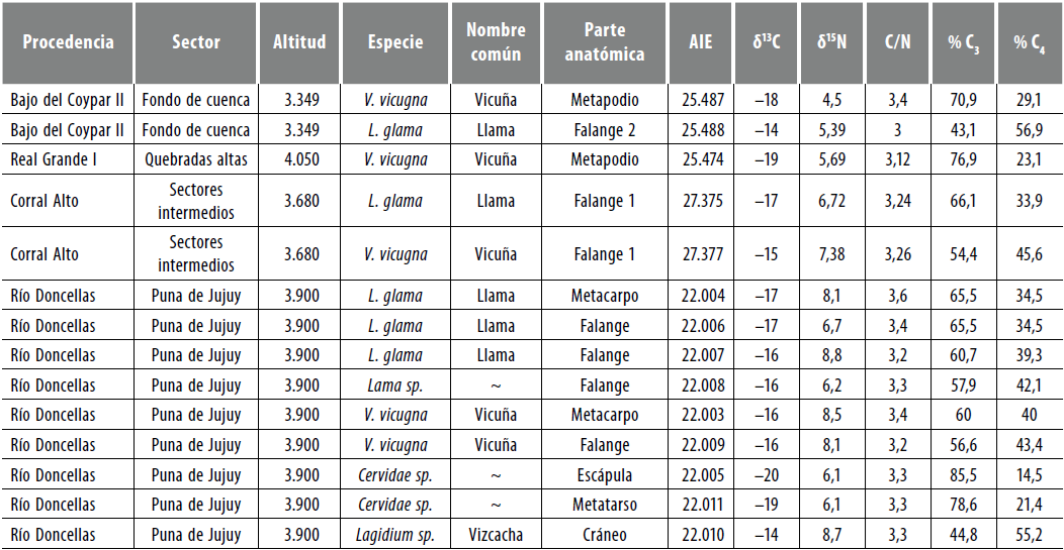
En cuanto a los recursos faunísticos arqueológicos, contamos con dos ejemplares de Lama glama y tres de Vicugna vicugna provenientes de sitios de AS. Por otro lado, dados los escasos valores δ15N para camélidos del Holoceno Tardío en puna, decidimos incluir una pequeña muestra de la localidad arqueológica Río Doncellas (de ahora en adelante LARD), ubicada en la puna de Jujuy, con una cronología asociada al periodo Tardío o de Desarrollos Regionales (Pérez y Killian 2011). Esta consta de dos Lama glama, dos Vicugna vicugna, una Lama sp., dos Cervidae sp. y un Lagidium sp. (probablemente vizcacha).
Con respecto a los restos humanos, las mediciones 513C y δ15N se hicieron sobre 5 individuos, provenientes de los sitios El Aprendiz (quebrada de Miriguaca, sectores intermedios), Casas Viejas 1 y 2 (margen occidental del río Punilla, fondo de cuenca), Bajo del Coypar II (fondo de cuenca), La Alumbrera (fondo de cuenca) y Tambería El Peinado (mina de malaquita asociada con la ocupación inca). Para la discusión de los resultados incluimos los valores publicados y aquellos valores δ13C que acompañan fechados radiocarbónicos que sumaron un total de 14 observaciones (Araníbar et al. 2007; Babot et al. 2009). Excluimos 2 individuos subadultos procedentes de la microrregión que tienen valores δ15N muy positivos, resultado probablemente de su condición de lactantes. Consideramos importante aclarar que, si bien para el noroeste argentino se han postulado intercambios a gran escala que pudieron incluir movimientos migratorios, ninguno de los individuos se encontró acompañado de indicadores que reflejen un origen geográfico diferente.
Modelos de análisis y técnica analítica
Con el fin de poder dar cuenta de los recursos predominantes en las paleodietas, nos basaremos en los fraccionamientos isotópicos propuestos por Newsom et al. (2004) y Bocherens y Drucker (2003). A partir de la propuesta de los primeros autores, no solo se estimó la composición cárnica de las dietas (es decir, la dieta proteica), sino que se incluyó el consumo de vegetales. Para ello se estableció un esquema con los rangos isotópicos de los recursos probablemente consumidos, utilizando datos de fauna y flora generados en este trabajo, así como también los aportados por otros investigadores (Fernández y Panarello 1999-2001; Killian y Salminci 2014; Samec, Morales y Yacobaccio. 2014; Yacobaccio, Samec y Cata 2010).
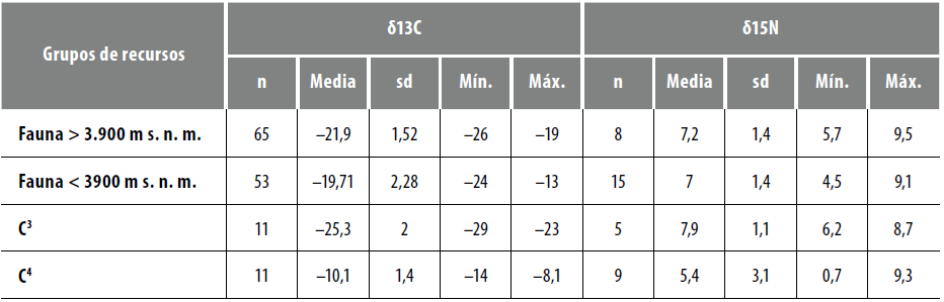
Uno de los requerimientos en la estimación paleodietaria es utilizar grupos de alimentos que sean diferentes a escala isotópica, por lo tanto, se generaron 4 grupos (tabla 3): 1) recursos cárnicos a altitudes mayores a los 3.900 m s. n. m.; 2) recursos cárnicos por debajo de los 3.900 m s. n. m.; 3) vegetales C3, y 4) vegetales C4. En el caso de los recursos cárnicos, se agrupó lo generado en el marco de esta investigación y lo ya publicado. La mayor cantidad de mediciones se hicieron sobre camélidos (n = 106 para valores δ13C y n = 20 para valores δ15Ν), sin embargo, se han agrupado de manera conjunta diferentes recursos faunísti- cos (incluyendo 12 valores δ 13C de Octodontomys gliroides, Felis sp., Dusicyon sp., Ctenomys sp. y Puma concolor). La estratificación espacial mencionada para los recursos cárnicos responde a la expectativa teórica que indica que por encima de los 3.900 m s. n. m. habría una mayor disponibilidad de humedad (con consecuentes valores δ15Ν más bajos) y mayor proporción de plantas C3. A su vez, se deja constancia de que bajo el rótulo de C3 incluimos diferentes recursos con señales isotópicas similares (Solanum tuberosum, Chenopodium quinoa, Prosopis sp. y Geoffroea decorticans), mientras que bajo el rótulo C4 incluimos los valores de maíz (Zea mays) y un ejemplar de amaranto (Amaranthus caudatus), procedente de puna de Jujuy, inédito (δ13C -11,9 ± 0,04; δ15N + 4,9 ± 0,07).
Tabla 3 Descripción de los valores medios, desvíos estándar, valores mínimos y máximos de los conjuntos de recursos probablemente consumidos
Fuente: Elaboración propia.
Asimismo, discutiremos los valores de 13C sobre hidroxiapatita (δ 13Cap), con el fin de no incurrir en sesgos interpretativos (Krueger y Sullivan 1984). De este modo, se hará referencia a la inferencia paleodietaria realizada sobre la fracción mineral o hidroxiapatita como dieta total, y se analizará su aporte energético (lípidos y carbohidratos). Al respecto, consideramos que la diferenciación entre dieta proteica y dieta total resulta pertinente, pues de no considerar una vía complementaria, la señal isotópica de un recurso tan relevante como el maíz puede verse subrepresentada dado el bajo contenido proteico de este cereal. Para comprender la información procedente de la fracción mineral, recurriremos a un modelo construido a partir de la observación de dietas controladas propuesto por Kellner y Schoeninger (2007) 5.
Las mediciones isotópicas y el pretratamiento de las muestras se realizaron en el Laboratorio de Isótopos Ambientales del Instituto de Geocronología y Geología Isotópica (Ingeis/UBA-Conicet). Para la extracción de colágeno, se siguieron las recomendaciones metodológicas de Tykot (2004). El procedimiento consta de dos fases, la desmineralización y la eliminación de partículas posdepositacionales. El primer paso requiere un ataque con ácido clorhídrico (HCl 2 %) por 72 horas, cambiando el reactivo cada día. En etapas anteriores y posteriores a este paso, el material se sumerge por 24 horas en hidróxido de sodio con el fin de eliminar ácidos húmicos. Luego, la muestra es enjuagada y secada en una estufa a < 60 °C.
En el pretratamiento de la fracción inorgánica se utilizaron los protocolos propuestos por Tykot (2004) y Garvie-Lok et al. (2004). El pretratamiento de vegetales consta del lavado ultrasonido, el secado en un horno a 60 °C y luego la molienda manual. Para la medición de valores δ13C y δ15Ν en colágeno y vegetales, se utilizó un analizador elemental Carlo Erba EA1108, acoplado a un espectrómetro de masas para la determinación de relaciones isotópicas (IRMS) Thermo-Scientific Delta V Advantage, utilizando una interfaz ConFlo IV. La incertidumbre analítica es de 0,2 %% en ó13C y δ15N. El análisis de la composición isotópica del carbono en hidroxiapatita se realizó con el método de ácido fosfórico desarrollado por McCrea (1950) y modificaciones posteriores (Panarello et al. 1980). El espectrómetro empleado en Ingeis es un Delta S Finnigan Mat triple colector. La incertidumbre analítica es de 0,1 %% (±2σ) en δ13C.
Resultados
Fauna
Todas las piezas óseas analizadas presentaron relaciones C/N aceptables. Los valores isotópicos se encuentran en la tabla 4. Si consideramos los conjuntos de Camelidae de AS y LARD se debe destacar que no hallamos diferencias entre los conjuntos respecto a los valores δ13C, cuya la media fue ± SD -16,5% ± 2 y -16,40/oo ± 0,5, respectivamente. No obstante, en el primer conjunto se evidencia una mayor variabilidad debido probablemente a que la muestra se compone de especímenes procedentes de tres sitios ubicados a diferentes altitudes, y se halla un valor muy positivo en un sitio de fondo de cuenca (Lama glama de Bajo del Coypar II, δ13C -13,7 %o). Por su parte, la diferencia entre los conjuntos es mayor respecto a los valores δ15N. Para el primero, la media ± SD es + 5,9 /% ± 1,1 y para el segundo, es + 7,7/ ± 1, con una variabilidad similar en el interior de estos dos casos. A su vez, al considerar las especies en total, se halló una diferencia menor a 1 /% entre llamas y vicuñas, tanto en 13C (media ± SD δ13C -16,2% ± 1,4; -17 % ± 1,4) como en 15N (media ± SD ó15N + 7,1% ± 1,3; + 6,4 % ± 1,6). Los ejemplares de cérvido provenientes de LARD presentan valores δ13C más bajos que el conjunto de camélidos, aunque en el caso del 15N no se hallan diferencias. En los valores de Lagidium sp. no se observan diferencias para ninguna de las relaciones isotópicas.
Tabla 4 Valores δ 13C y δ15Ν generados en el marco de esta Investigación para fauna arqueológica de Antofagasta de la Sierra y la localidad arqueológica Río Doncellas (departamento de Cochinoca, puna de Jujuy)
Fuente: Elaboración propia
En lo que respecta a la estimación paleodietaria6, observamos que los ejemplares de cérvido tienen una dieta dominada por especies C3. Una llama (Lama glama) proveniente del sitio Bajo del Coypar II y el caso de Lagidium sp. integran de manera más equilibrada especies C3 y C4. El resto de los ejemplares integra menos del 35 % de especies C4 en su dieta.
Resulta pertinente dar cuenta de la presencia de dos ejemplares de Cameli- dae hallados en el sitio Bajo del Coypar II, donde se advierte que uno de ellos tuvo una ingesta con un gran aporte de pasturas C4 propias, probablemente de la altitud donde se emplaza el sitio (fondo de cuenca), en tanto que de otra muestra se infiere una dieta con un mayor aporte de C3, correspondiente a áreas de pastura en cotas más elevadas (considerando los análisis de Fernández y Panarello 1999-2001).
Restos esqueletarios humanos
Todos los individuos pudieron ser analizados en ambas fracciones del material y contaron con relaciones C/N aceptables, con excepción de uno de Bajo del Coy- par II que no tuvo un rendimiento suficiente de colágeno para poder realizar la medición isotópica y solo pudo medirse la fracción inorgánica. Los resultados de los análisis de las muestras de material óseo se encuentran en la tabla 5, junto con los valores publicados por otros investigadores.
Tabla 5 Valores δ13C y δ15N en restos humanos generados en el marco de esta investigación y anteriores1;2Olivera y Yacobaccio (1999);3Araníbar et. al. (2007);4Babot et al. (2009)
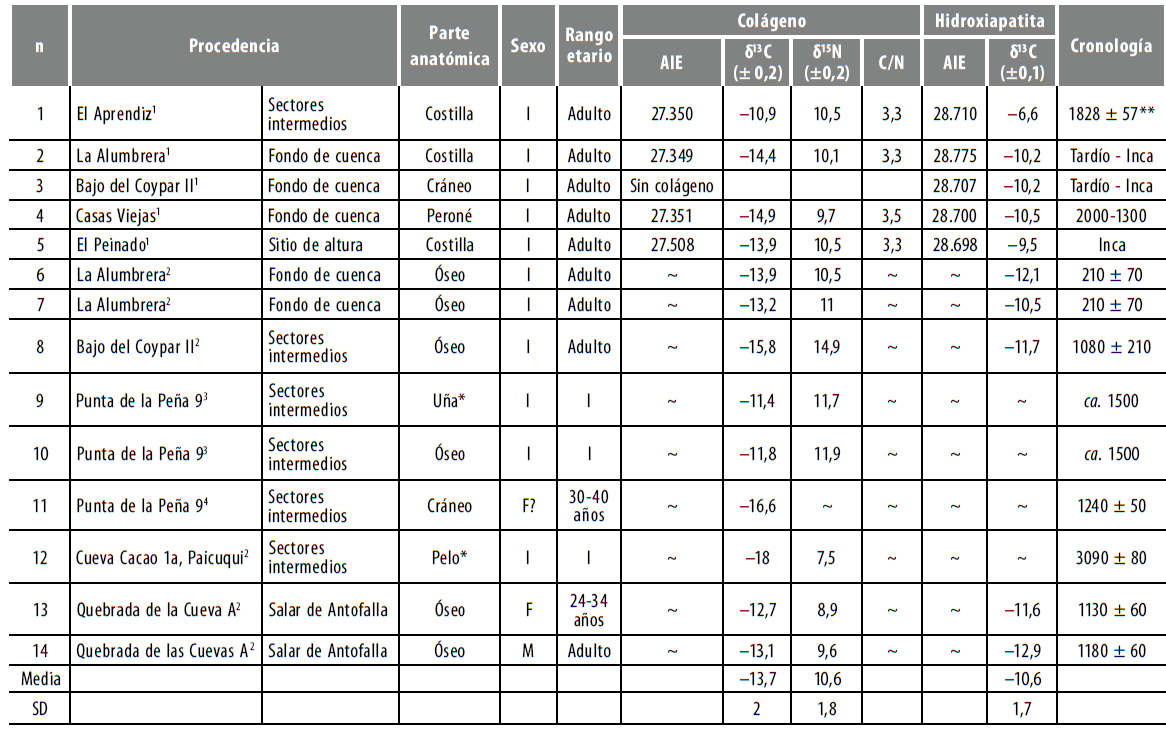
* Los valores en una corresponden al promedio de 7 mediciones; el DE de los valores de δ13C es igual a 1,16 y a 0,58 en el caso del δ15N. Los valores fueron corregidos para ser comparados con colageno oseo.
** AMS Arizona (ESCO-001); δ13C -11,4.
Fuente: Elaboración propia.
Considerando todos los valores disponibles para AS, se halla un rango pequeño de variación en el interior de cada sistema isotópico, con una media de ± SD para δ13Cco -13,7 ± 2 %; δ15Ν 10,5 % ± 1,8 y para δ13Cap, -10,5 ± 1,7 <%>. En el caso de los valores 513Cco, la amplitud del rango de variación está dada principalmente por la medición realizada en pelo proveniente del sitio Cueva Cacao 1 (δ13C -18 %o). Mientras que en los valores δ15N la amplitud del rango se debe a dicha muestra (δ15Ν + 7,5 %), pero también al individuo que sí pudo medirse en Bajo del Coypar II (δ15N + 14,9 %o). Es importante recalcar que, aunque los valores δ13Cap se encuentran en general empobrecidos, hay un individuo con un valor alto en el sitio El Aprendiz, que mide ó13Cap = -6,6 %o.
Valores isotópicos y procedencia de los individuos
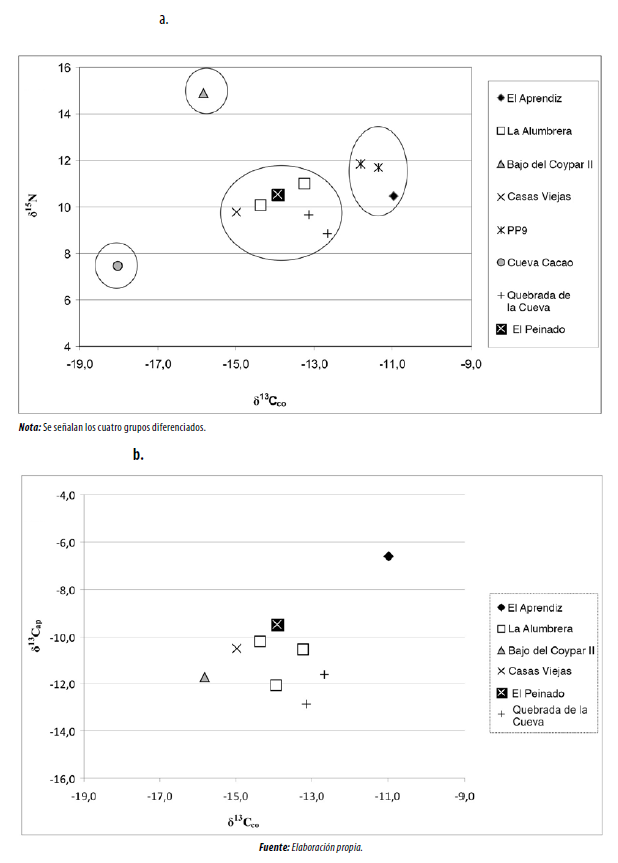
A partir de la información isotópica en el colágeno óseo, es posible diferenciar al menos cuatro grupos, aunque dos de ellos se componen cada uno de un solo individuo (figura 2a). El primero es el caso del individuo hallado en Cueva Cacao 1, con la cronología más temprana del conjunto, que se aleja de este principalmente por el bajo valor en 15N. Esta medición se hizo sobre pelo y fue representativa de una observación estacional y no de los últimos diez años de vida, como es el caso del colágeno óseo. El otro grupo comprende uno de los dos individuos de Bajo del Coypar II, que tiene un valor llamativamente alto en 15N, en comparación con el resto de los valores de la microrregión. Sin embargo, cabe resaltar que dicho valor no resulta anómalo si consideramos otros provenientes de puna en el NOA (Pérez y Killian 2011). El tercer grupo se compone de tres individuos de sitios emplazados en los sectores intermedios, es decir, Punta de la Peña 9 y El Aprendiz, los cuales muestran valores más positivos en 13C. El cuarto grupo, compuesto por individuos hallados en sitios de fondo de cuenca, el área de Antofalla y un individuo hallado en un sitio incaico ubicado a gran altitud (> 4.500 m s. n. m.), tiene valores más bajos respecto al 13C.
Al cruzar la evidencia en 13C de ambas fracciones del registro óseo (figura 2b), resulta más dificultoso diferenciar grupos y no se establece una correlación entre ambas mediciones, lo cual implica la existencia de patrones fotosintéticos dominantes diferentes en la dieta proteica y total en cada individuo. La única excepción a la mencionada tendencia resulta del individuo hallado en los sectores intermedios de El Aprendiz, que muestra un aporte importante de recursos bajo patrón fotosintético C4, tanto en lo que respecta al componente cárnico como vegetal de la dieta asimilada.
Tendencias temporales
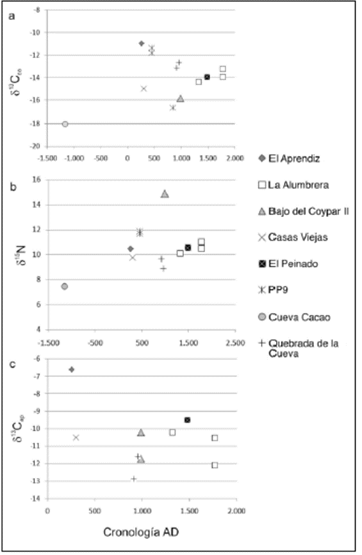
Con el fin de analizar las tendencias temporales, se consideraron los valores isotópicos de 13C (en colágeno e hidroxiapatita) y 15N de manera individual. Una parte de la muestra (n = 9) posee fechados directos, lo cual facilitó su adscripción a un rango temporal. Por su parte, los individuos restantes (n = 5) fueron ubicados temporalmente mediante fechados asociados y, en menor medida, a partir de una asignación cronológica relativa, dadas las características contextuales de los hallazgos. Consideraremos como un límite, algo arbitrario, los 1000 años a. D. para diferenciar a los individuos adscritos al periodo de consolidación agropas- toril de aquellos del periodo Tardío.
Como se observa en la figura 3 (a, b y c), no se registran tendencias claras en los sistemas isotópicos analizados, es decir, no hay una relación lineal entre el paso del tiempo y un incremento en la señal isotópica del 13C o el 15N. Los individuos con cronologías cercanas al 1000 a. D., o sea, en el momento transicional a las sociedades tardías, presentan valores con diferencias entre el 3 y 4 %%, tanto en los valores δ13C como en δ15Ν, con una diferencia más acentuada en el caso del nitrógeno (figuras 3a y 3b). Es probable que esto resulte de las diferencias geográficas en las áreas de emplazamiento de los sitios con una consecuente repercusión a escala isotópica, pues el periodo señalado cuenta con los individuos provenientes del área cercana al Salar de Antofalla, que tiene mayor humedad y precipitaciones respecto al resto de AS.
Fuente: Elaboración propia.
Figura 3 Valores δ13C en colágeno (a), valores δ13C en hidroxiapatita (c), valores δ15Ν (b) y cronología en años calendáricos (a. D.) Nota: Para calibrar los fechados radiocarbónicos 14C y homogeneizar la información, se utilizó la curva ShCal04 (McCormac et al. 2004) y el software OxCal v4.1.6 (Bronk Ramsey 2009).
Por otro lado, es alrededor del 500a. D. y no posteriormente cuando se hallan los valores δ13Cco más positivos. Esto también coincide con que se encuentra el individuo de El Aprendiz con un valor de 13C extremo en hidroxiapatita (figura 3c). Este aspecto resulta llamativo porque el resto de los valores en esta fracción se halla siempre al menos un 4 % más empobrecido. El resto del conjunto no muestra diferencias al considerar su adscripción temporal. Asimismo, teniendo en cuenta los valores δ13Cap y el fraccionamiento de dieta estimada de -9,4 %% (Ambrose y Norr 1993) para hidroxiapatita, el aporte de recursos C3 en la dieta es dominante en todos los casos.
Componente predominante de la dieta proteica
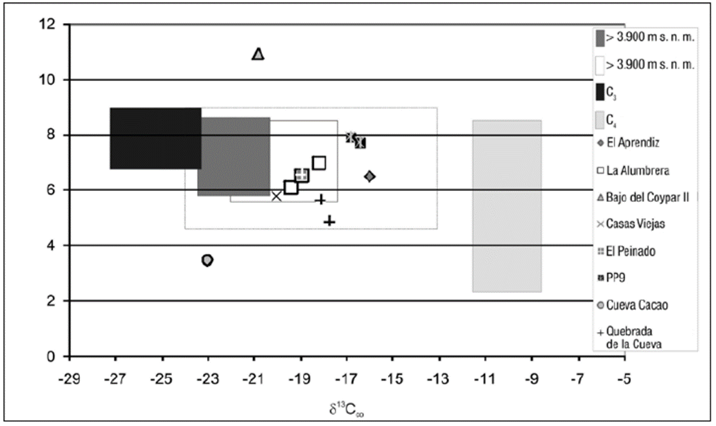
A partir de la información de la ecorregión argentina de puna y la que arroja esta investigación, se establecieron cuatro grupos de recursos para la estimación paleodietaria. En la tabla 3 se describe la estadística descriptiva de cada uno de ellos y en la figura 4 se observan los valores de las dietas humanas estimadas los rangos isotópicos establecidos para los conjuntos de recursos. Resulta oportuno señalar que al establecer el rango de distribución de vegetales de consumo humano C3, en lo que respecta al 15N, este es similar al de los recursos cárnicos. Probablemente esto se deba, en parte, a los requerimientos de abono de las tierras cultivadas, como se ha mencionado en Killian y Salminci (2014).
Fuente: Elaboración propia.
Figura 4 Valores δ 13C y δ15Ν de recursos corregidos y dietas humanas estimadas Nota: El recuadro en línea punteada delimita los valores máximos y mínimos del grupo fauna por debajo de los 3.900 m s. n. m. Se corrigieron los valores obtenidos en colágeno óseo de fauna en un 4 ‰ en el caso del 13C, y se pasaron de este modo a lo directamente consumido (músculo). Las dietas estimadas de los humanos suponen un fraccionamiento de 5 y 4 ‰ para los valores δ13C y δ15N, respectivamente. Los valores δ13C de muestras animales y vegetales modernas se han corregido considerando el denominado efecto Suess (+ 1,5 ‰, Craig 1957).
Como puede observarse, la distribución de los valores estimados de las dietas proteicas coinciden en mayor medida con los rangos establecidos para los recursos faunísticos de puna, al ubicarse de manera equidistante respecto a los recursos vegetales, tanto C3 como C4 (figura 4). Solo en dos casos no hay coincidencias. Se trata, por un lado, de la medición efectuada sobre pelo humano hallado en Cueva Cacao 1 y, por otro, del caso del individuo hallado en el sitio Bajo del Coypar II que tiene un valor 15N extremadamente positivo, que si bien no puede explicarse a partir de los valores en fauna de este trabajo, sí podría coincidir con los rangos de distribución obtenidos en puna de Jujuy que se encuentran en proceso de publicación (Samec, comunicación personal). En el caso de Cueva Cacao 1, debemos recordar que el mechón de pelo analizado, una trenza humana, se halló con un conjunto de elementos manufacturados con materias primas provenientes de camélidos, por lo cual, esta falta de coincidencia con el componente cárnico debe entenderse a partir del material observado. Es decir, una observación estacional (pues no es un promedio de observaciones, producto de diferentes mediciones) poco representativa del ciclo anual alimentario. Ahora bien, las dietas estimadas que sí encuentran coincidencias con los recursos cárnicos lo hacen si consideramos los valores extremos, tanto en el conjunto correspondiente a recursos faunísticos de sectores altos como bajos. De igual modo, existe una coincidencia entre cuatro dietas estimadas y la fauna procedente de sectores por debajo de los 3.900 m s. n. m., al tener en cuenta los valores máximos y mínimos de este conjunto. De no considerarse los valores extremos de los recursos faunísticos, tendríamos incertidumbre sobre cuatro dietas estimadas, en adición a las dos ya mencionadas. Podría tratarse de individuos que tuvieron un mayor componente de recursos C4 en su dieta, como el maíz y el amaranto, dado que se presentan señales δ13C más positivas que el resto y más cercanas a la distribución de estos recursos. Por lo tanto, es necesario realizar el análisis isotópico sobre hidroxiapatita, ya que el valor allí obtenido refleja tanto el componente proteico de origen animal, como el componente energético proveniente de recursos como los cereales o pseudocereales.
El aporte de vegetales
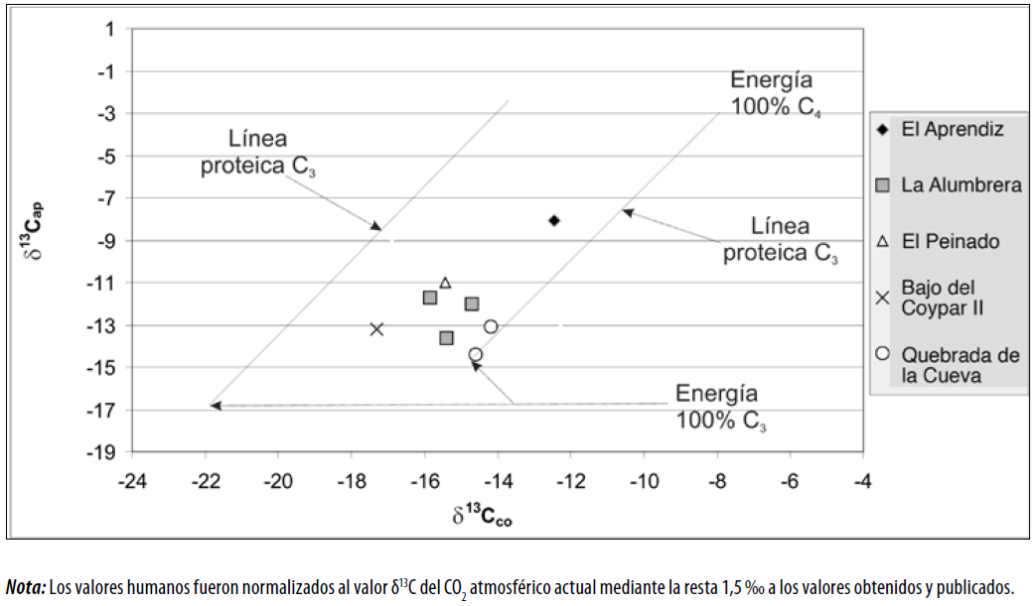
En la figura 5 se exponen los valores δ 13C en colágeno e hidroxiapatita emplazados en las líneas de regresión propuestas por Kellner y Schoeninger (2007). Con respecto a la dieta inferida a partir del registro inorgánico, se trata de individuos que no tuvieron un aporte sustancial de energía (carbohidratos y lípidos) bajo el patrón fotosintético C4 en su dieta, tal como queda evidenciado por su ubicación en la mitad inferior de las líneas de regresión propuestas por las autoras. Por lo tanto, puede decirse que, en la estimación de la dieta total, no se hallan representados el maíz ni el amaranto, incluso en los casos de Punta de la Peña 9 y Quebrada de las Cuevas, en Antofalla. Sin embargo, bajo este modelo, el individuo que proviene del sitio El Aprendiz, ubicado en los sectores intermedios de AS con una cronología ca. 500 a. D., sí posee una dieta energética con un mayor componente C4 en su dieta respecto a los demás.
Discusión y conclusiones
A partir de la muestra analizada, que corresponde casi en su totalidad a las ocupaciones de los milenios primero y segundo de la era en AS, los datos obtenidos no nos permiten afirmar que haya existido un correlato claro entre la composición dietaria humana y los cambios económicos registrados con base en otras evidencias arqueológicas en el área de estudio. Específicamente, no encontramos una tendencia que indique un reemplazo del consumo del componente cárnico, proveniente de la caza y el pastoreo, por el maíz. Este planteamiento se basa en la ausencia de evidencias que indiquen un enriquecimiento progresivo en los valores δ13C (figura 3, a y c), o un cambio en los patrones inferidos a partir de la estimación paleodietaria (figuras 4 y 5). Asimismo, se podría considerar como un indicador de la disminución del consumo de carne el empobrecimiento progresivo de los valores δ15N, hecho que tampoco observamos a partir de los datos presentados (figura 3b).
Con respecto a la inferencia paleodietaria, el análisis de la dieta proteica advierte la importancia de los recursos cárnicos por encima de otros, y los recursos faunísticos consumidos tendrían un aporte sustantivo de pasturas bajo el patrón fotosintético C4 (figura 4). Al respecto, si bien en áreas más bajas de la provincia de Catamarca (Izeta et al. 2010) se ha postulado una estrategia de engorde a base de maíz para las llamas, en el caso de AS, esta situación podría haberse dado a partir de la misma composición de las pasturas, con una mayor proporción de especies bajo esta vía metabólica, aunque no se descarta la posibilidad de esta práctica de forraje complementario de maíz en el área de estudio. En concordancia con el planteamiento, debemos recordar la alta proporción de C4 en las dietas de camélidos registrada para cronologías anteriores al Holoceno Tardío (Mondini, Panarello y Zangrando 2010) y, por lo tanto, presumiblemente anteriores a la introducción del maíz en la economía. Debemos tener presente además que, en la pequeña muestra de camélidos analizada en este trabajo, se ha dado cuenta de la alta proporción de dicha pastura en la dieta de uno de los ejemplares procedente de AS.
Por su parte, a la hora de analizar la dieta total, los recursos energéticos predominantes, es decir, lípidos y carbohidratos, se emplazan en la vía C3 (figura 5). Esto indicaría que la energía dietaria estaría aportada principalmente por recursos cultivados, como tubérculos o quínoa, o de recolección, como es el caso del chañar (Geoffroea decorticans) y el algarrobo (Prosopis sp.) (hallados en sitios arqueológicos del área, como se describe en Babot [2009], Babot et al. [2009], entre otros), sin detectarse presencia importante de maíz.
Por otra parte, resulta llamativo que el valor con más aporte de energía C4 en la dieta se halle en un individuo con una cronología del primer milenio (sitio El Aprendiz), a la vez que los individuos con mayor aporte C4 en su dieta en general (sitio Punta de la Peña 9) también se adscriben a una cronología cercana a los ca. 500 a. D. Debemos recordar que para ese momento se ha postulado el incremento de la intensificación agrícola, tanto en los valles de Catamarca como en Antofalla (Quesada 2010), y aunque hay importantes evidencias sobre cultivo en la microrregión, la mayor relevancia de la agricultura dentro de la economía antofagasteña parece demostrarse a partir del inicio del segundo milenio (Olivera y Vigliani 2000-2002; Olivera et al. 2003-2005).
Más allá de lo expuesto, el hecho de no encontrar por el momento claros indicadores de consumo humano de maíz no implica que este no se produjera. Si consideramos el registro arqueológico de diferentes partes del mundo, la importancia de este cereal ha sido señalada más allá de sus propiedades nutricio- nales en la esfera político-social, cuando la alimentación genera y perpetúa la heterogeneidad y jerarquía (Ambrose et al. 2003; Hastorf y De Niro 1985; Hastorf 1990), al emplearse de modo ritualizado (Hastorf 2003) o cuando se convierte en un recurso para la producción de excedentes para el intercambio (Burger y Van der Merwe 1990). Incluso para la microrregión se ha postulado la hipótesis de un uso del cereal como forraje (Olivera 1997). Por otro lado, la puesta en duda de la importancia de este cereal frente a otros recursos también se ha señalado en otro caso de la puna argentina, a partir del análisis de la muestra esqueletaria humana procedente de la localidad arqueológica Río Doncellas. Allí, la evidencia isotópica también señala la relevancia de los recursos cárnicos respecto a los vegetales y la probable predominancia de recursos como tubérculos y quínoa entre estos últimos (Killian et al. 2012; Pérez y Killian 2011). Resulta necesario aclarar que dichas conclusiones se confrontan con las evidencias macrobotánicas y los indicadores de una economía con un extenso desarrollo de la agricultura registrados en esa área. También, en el caso de AS, el hallazgo de una gran cantidad de marlos en la cueva de Quebrada de Petra (Oliszewski y Olivera 2009; Olivera y Vigliani 2000/2002) puede reflejar una situación similar: la valoración social y política del recurso, más allá de una menor incidencia de su consumo cotidiano como alimento base. Por supuesto que estas ideas requerirán profundización en diversas líneas de la investigación para contrastar su validez dentro del proceso regional.
Este trabajo expuso una vía metodológica independiente para el estudio de las diferentes estrategias de consumo en Antofagasta de la Sierra a lo largo del Holoceno Tardío. De este modo, el análisis paleodietario ha permitido avanzar en la jerarquización de los recursos alimentarios, en los que se destaca la relevancia de los recursos cárnicos, producto del pastoreo y la caza. En el caso del maíz, al menos por el momento, no podemos afirmar que este haya sido importante en la dieta habitual de los casos analizados, lo que nos conduce a pensar que la intensificación agrícola debió recaer en otras especies mejor adaptadas a la alta aridez y a la altitud, como es el caso de la quínoa y de los tubérculos. Es decir, la importancia de la producción cerealera quizá no haya sido el corolario de largos procesos de adaptación de la agricultura a este desierto de altura, incluso para el desarrollo de las complejas sociedades del Tardío y aun durante la ocupación incaica.
A partir de ahora tenemos nuevos e interesantes interrogantes para seguir profundizando esta investigación, y se hace necesario no solo ampliar las muestras óseas y dentales humanas para el análisis isotópico, sino ahondar en el conocimiento de la variabilidad a escala isotópica en todos los componentes de la ecología microrregional, en particular, aquellos con incidencia directa en la cadena trófica de la que los humanos formaron parte.