











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroducción
El noroccidente de Sudamérica es un área clave para el estudio del poblamiento inicial del subcontinente, debido a su posición geográfica, el marcado gradiente ambiental y su diversidad biocultural. En dicha región se han recuperado importantes evidencias a nivel arqueológico (véase síntesis en López 2008; Aceituno et al. 2013; Delgado et al. 2015a, 2015b) y bioantropológico (véase síntesis en Delgado 2012c; Delgado 2015). Igualmente, a partir de la evidencia palinológica, glaciomorfológica, de diatomeas e isotópica, se han llevado a cabo reconstrucciones sobre las condiciones ambientales prevalentes durante los últimos 15.000 años (véase síntesis en Marchant et al. 2002; Van der Hammen y Hooghiemstra 1995). La evidencia arqueológica en un contexto paleoambiental, junto con otras evidencias bioantropológicas y bioarqueológicas, ha permitido la formulación de distintas hipótesis sobre el tiempo, patrón y proceso del poblamiento inicial, así como sobre el número de grupos que ingresaron, su diversidad biocultural y su microevolución holocénica. En este trabajo se presenta un panorama de las investigaciones realizadas en torno al poblamiento inicial de Colombia. Primero se describe el registro arqueológico temprano, su contexto cronológico, paleoambiental y las relaciones interregionales; luego se exponen algunos resultados del estudio de la evidencia bioantropológica en grupos actuales y prehispánicos que incluyen variantes moleculares y rasgos fenotípicos. Por último, se bosqueja un escenario de poblamiento unificado a partir de los datos disponibles, se discuten brevemente algunos aspectos puntuales de las diferentes líneas de evidencia consideradas y se proponen algunos énfasis de trabajo a futuro.
El contexto arqueológico
El registro arqueológico temprano del noroccidente de Sudamérica
La información sobre el primer poblamiento humano de Colombia es aún escasa. El registro arqueológico de las ocupaciones tempranas se ubica en la cordillera Oriental, el valle del Magdalena, las cordilleras Central y Occidental, el Valle de Pubenza y la cuenca del Amazonas (figura 1). A la fecha no existe información relevante de regiones como las tierras bajas del Caribe, la cuenca del Pacífico, los Llanos Orientales y gran parte de la cuenca del Amazonas (Aceituno et al. 2013).
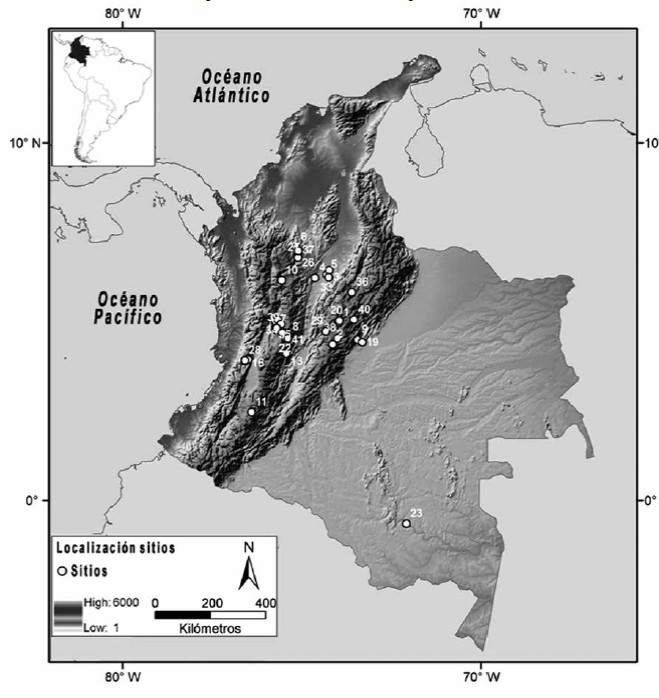
Fuente: Elaboración del autor empleando ArcGIS 9.3, licencia UNLP
Figura 1 Mapa de la distribución geográfica de los sitios arqueológicos tempranos (rango 12.000-8000 años 14C a. P.) de la región de estudio Nota: 1) Tibitó, 2) Tequendama I, 3) La Palestina 2, 4) Nare, 5) San Juan de Bedout, 6) PIII0I-52, 7) El Guatín, 8) El Jazmín, 9) Sueva I, 10) La Morena, 11) San Isidro, 12) La Palestina 1, 13) El Jordán, 14) 66PER001, 15) Salento 24, 16) Sauzalito, 17) La Trinidad corte I, 18) La Selva, Risaralda, 19) Gachalá, 20) El Abra II, 21) La Trinidad corte II, 22) La Pochola, 23) Peña Roja, 24) Génova, 25) La Montañita, 26) Sitio 045, 27) Sitio 021, 28) El Recreo, 29) Galindo I, 30) Nuevo Sol, 31) La Selva, 32) 39-El Recreo Cancha, 33) Peñones de Bogotá, 34) Salento 21, 35) El Antojo, 36) Neusa, 37) PIII0P-59, 38) Páramo Peña Negra, 39) La Chillona, 40) Checua, 41) San Germán II.
La cordillera Oriental
Los fechados más tempranos de la región provienen de dos abrigos rocosos, El Abra II y Tequendama I, así como de un sitio al aire libre (Tibitó). En El Abra se hallaron lascas y herramientas unifaciales de tipo expeditivo, lo cual le dio el nombre de industria abriense o tradición lítica de bordes erodados (edge-trimmed tool tradition) (Correal 1986; Hurt, Correal y Van der Hammen 1977). Dicho registro lítico fue hallado en los niveles más profundos (ca. 12.400 años 14C a. P.) junto con restos óseos de fauna reciente (e. g. venado, Odocoileus virginianus; conejo, Sylvilagus brasilensis; curí, Cavia porcellus). Varias de las fechas se ubican entre el Pleistoceno Final y el Holoceno Temprano. En Tequendama I, Correal y Van der Hammen (1977) excavaron una serie de abrigos rocosos cuyos niveles inferiores fueron datados entre ca. 12.500 y 10.900 años 14C a. P., los cuales contenían restos de fauna similares a los hallados en El Abra II y lascas tipo abriense. Entre ca. 11.000 y 10.000 años 14C a. P., una tecnología lítica distinta aparece en el registro; es denominada tequendamiense y difiere de la abriense en el uso de materias primas alóctonas y la presencia de raspadores, lascas reducidas y un fragmento de punta de proyectil (Aceituno et al. 2013; Correal y Van der Hammen 1977; Delgado et al. 2015b). Recientemente, Nieuwenhuis (2002) identificó cambios importantes en la manufactura y uso de artefactos abrienses, así como la aparición de artefactos tecnológicamente más complejos no atribuibles a la tecnología tequendamiense, el uso de herramientas en contextos y actividades no especializadas (economías de amplio espectro) y la importancia del trabajo de materiales y recursos vegetales (para una opinión distinta, véase Correal 2003).
Tibitó, un sitio de desposte ubicado al aire libre excavado por Correal (1979), fue datado en 11.740 ± 110 años 14C a. P. (GrN-9375). Allí se recuperaron herramientas abrienses en asociación con restos óseos de proboscídeos (Haplomastodon sp. y Cuvieronius hyodon), caballo americano (Equus sp.) y venado (Odocoileus virginianus) (Correal 1981). Aparte, en la sabana de Bogotá, la investigación de otros sitios ubicados en abrigos o al aire libre, tales como Sueva I, Gachalá, Galindo I, Neusa y Checua, permitió el descubrimiento de artefactos abrienses, restos óseos humanos y restos de fauna holocénica datados entre ca. 10.000 y 8000 años 14C a. P.
El suroccidente de Colombia
En la región Calima (cordillera Occidental) se excavaron dos sitios ubicados al aire libre -Sauzalito y El Recreo- pertenecientes al Holoceno Temprano, en donde se recuperó un repertorio lítico compuesto por lascas unifaciales, azadas, manos de moler, golpeadores y yunques fabricados con rocas metamórficas locales (Salgado 1988-1990). En el Valle de Pubenza, departamento del Cauca (cordillera Central), en el sitio San Isidro, se recuperaron evidencias en un nivel datado hacia la frontera Pleistoceno/Holoceno; entre ellas, una tecnología lítica compuesta por cientos de artefactos manufacturados en chert y obsidiana, como lascas retocadas y no retocadas, bifaces lanceoladas y manos de moler sin asociación a restos de fauna (Gnecco 2000). También se hallaron semi llas carbonizadas de Persea spp. y Erythrina, así como granos de almidón de Xanthosoma, Ipomoea, Manihot y Maranta arundinacea, además de otros pastos y leguminosas (Mora y Gnecco 2003).
La cuenca del Amazonas
Estudios arqueológicos en la cuenca del Amazonas permitieron el hallazgo de Peña Roja, un sitio al aire libre ubicado en el Caquetá medio y datado a inicios del Holoceno (Cavelier et al. 1995; Gnecco y Mora 1997). En este sitio, el conjunto lítico estuvo compuesto por hojas unifaciales, raspadores, raederas, lascas concoidales, taladros, choppers y cuñas fabricadas en cuarzo, chert, rocas ígneas y metamórficas, así como placas de molienda, cantos rodados con bordes desgastados, golpeadores y yunques (Cavelier et al. 1995; Gnecco y Mora 1997). Adicionalmente, se hallaron cientos de semillas carbonizadas pertenecientes a diferentes géneros de palmas (Morcote et al. 1998), junto con fitolitos de Lagenaria spp., Calathea allouia y Cucurbita spp. (Mora y Gnecco 2003), lo que sugiere la importancia de la explotación de recursos vegetales entre los grupos amazónicos tempranos. Recientemente, Morcote, Aceituno y León (2014) presentaron nuevas evidencias que incluyen fechados ubicados en el 9000 a. P., así como restos arqueobotánicos y herramientas líticas que expanden los hallazgos previos.
El Magdalena Medio
En las tierras bajas tropicales del Magdalena Medio, una serie de sitios tempranos, incluyendo Nare, La Palestina 1 y 2, San Juan de Bedout y Peñones de Bogotá, fueron descubiertos y excavados durante las décadas pasadas (para una síntesis, véase López 2008). En estos sitios estratificados localizados al aire libre y datados entre ca. 10.400 y 8500 años 14C a. P., se halló un repertorio lítico variado que incluye lascas simples, raspadores planoconvexos, puntas de proyectil cola de pez con alas rectas, oblicuas y redondeadas, y puntas delgadas manufacturadas en chert o cuarzo. Estas herramientas fueron halladas sin asociación con algún tipo de recurso vegetal o animal. Otero y Santos (2002) indicaron que estas fueron empleadas en la explotación de fauna local, compuesta por mamíferos, reptiles y aves de tamaño mediano y pequeño. López y Cano (2011) concuerdan al indicar que la prevalencia de ambientes boscosos y espacios ribereños durante la transición Pleistoceno/Holoceno indicaría la explotación de fauna aún existente. Ranere y López (2007) sugieren que sesgos tafonómicos e índices bajos de conservación pueden estar influyendo en la falta de relación entre la tecnología lítica, la fauna y evidencias arqueobotánicas.
El Cauca Medio
Recientemente, en el Cauca Medio y el valle del río Porce (cordillera Central, ca. 1.650-2.100 m s. n. m.), se excavaron una serie de sitios estratificados localizados al aire libre cuyos fechados llegan al Pleistoceno Final (Aceituno et al. 2013). En sitios como el Jazmín (Cauca Medio) (Aceituno y Loaiza 2015) y La Morena (Porce Medio) (Santos, Monsalve y Correa 2015), la tecnología lítica, consistente principalmente en lascas unifaciales, hachas, azadas, bases de molienda y cantos rodados con bordes desgastados, indica una clara orientación hacia la explotación de recursos vegetales (Aceituno y Loaiza 2015; Aceituno et al. 2013; Delgado et al. 2015a). Asimismo, Dickau et al. (2015) presentaron nuevas evidencias (fechas y sitios) para esta región, que complementan los resultados obtenidos hasta la fecha.
Sitios aún en discusión
En el contexto del poblamiento inicial de la región deben mencionarse dos sitios ubicados cronológicamente en el Pleistoceno Final, aunque su relación con el resto no puede establecerse aún. El primero es un sitio al aire libre llamado Pubenza, ubicado en las tierras bajas del valle del Magdalena en Cundinamarca, en donde se encontraron restos de mastodonte junto con lascas en un nivel datado en 16.400 ± 420 14C a. P. (GrN-19857) (Correal et al. 2005). Aspectos relacionados con la asociación entre artefactos y los restos de megafauna, así como su depósito en un contexto de alta energía y un fechado que se desprende del total de los sitios tempranos a escala regional, impiden por el momento y de manera fehaciente su integración al registro arqueológico temprano de Colombia. En el otro sitio denominado El Jordán (cordillera Central), localizado también a cielo abierto, se hallaron unos pocos artefactos en su nivel inferior, datados en 12.910 ± 60 (Beta-111972) (Salgado 1998). Si bien el fechado puede corresponder a la fase inicial de exploración, la escasa información arqueológica recuperada impide evaluar la integridad del sitio, y por lo tanto su utilidad en la discusión.
El tiempo del poblamiento temprano: inferencias a partir del análisis de fechados 14 C
Delgado et al. (2015b) compilaron y analizaron la base de fechados radiocarbónicos disponible para la región correspondiente a la transición Pleistoceno/Holoceno (ca. 12.000-8000 años 14C a. P.), con el fin de hacer inferencias sobre el tiempo y el patrón de poblamiento. Geográficamente, este estudio mostró que los sitios con fechas más tempranas están distribuidos en el altiplano cundiboyacense (cordillera Oriental) y en el valle del Magdalena Medio. Al occidente, la ocupación de las cordilleras Central y Occidental, así como del valle del río Cauca, es un fenómeno más tardío. Al oriente, el único sitio descubierto en Peña Roja, en la cuenca del Amazonas, sugiere la ocupación del bosque tropical durante el Holoceno Temprano. Asimismo, la distribución altitudinal de los sitios (0-500 m s. n. m., 1.000-2.000 m s. n. m. y 2.500-3.000 m s. n. m.) indica que notablemente los fechados más tempranos provienen del rango de 2.500-3.000 m s. n. m. (Tibitó y El Abra, altiplano cundiboyacense). La ocupación del piso térmico más bajo, representado por la cuenca del Magdalena Medio, es más tardía, alrededor de los 10.500 años 14C a. P. en sitios como Nare, La Palestina 2 y San Juan de Bedout. El segundo piso, ubicado entre los 1.000 y 2.000 m s. n. m. en las cordilleras Central y Occidental, registra ocupaciones hacia el inicio del Holoceno.
Un aspecto interesante que se desprende de dicho análisis es que los sitios más tempranos se ubican en las tierras altas, lo cual indica un sesgo metodológico debido a la mayor investigación en ciertas áreas; esto produce una distribución asimétrica del registro. De manera alternativa, cuestiones relacionadas con transformaciones drásticas del paisaje a causa de eventos climáticos y geológicos (e. g. vulcanismo) pudieron generar hiatos en las secuencias estratigráficas y producir también distribuciones asimétricas del registro. El análisis del patrón temporal de distribución de los fechados correspondientes a la transición Pleistoceno/Holoceno, tanto calibrados (2σ) como no calibrados, exhibe una misma tendencia: una señal débil e interrumpida antes de los 11.000 años 14C a. P. y fuerte y continua posterior a esa fecha, particularmente en la última parte del Pleistoceno.
En términos del escenario paleoambiental en el cual ocurrió el poblamiento inicial, si se aceptan las dos fechas previas a los 11.000 años 14C a. P., entonces la colonización del área probablemente comenzó durante el Interstadial Guantiva (ca. 12.500-11.000 años 14C a. P.), un periodo que presentó condiciones relativamente cálidas y húmedas que permitieron, en algunos casos, la expansión de biomas boscosos y un movimiento ascendente de la línea superior del bosque. Por el contrario, si ambas fechas se consideran provisionalmente como dudosas debido a su desprendimiento del resto, a pesar de que no se comportan como valores atípicos en un sentido estadístico, entonces el proceso de colonización empezó durante el Estadial El Abra contemporáneo con la Cronozona Younger Dryas (~ 11.000-10.000 años 14C a. P.). En cualquier caso, fue durante este pulso frío que la primera población fundadora alcanzó una indiscutible visibilidad arqueológica, representada por una diversidad de contextos depositados en diferentes escenarios ambientales en el altiplano cundiboyacense y la cuenca del Magdalena Medio.
Relaciones interregionales tempranas inferidas a partir de los conjuntos líticos
Los datos disponibles sugieren que, a escala regional, la diversidad en los conjuntos líticos puede ser explicada en términos geográficos y cronológicos que dan cuenta de las probables relaciones interregionales. Así, el flujo de materias primas desde el valle del Magdalena hasta el altiplano cundiboyacense, como se demostró con el conjunto lítico tequendamiense, indicaría patrones de alta movilidad (Correal y Van der Hammen 1977). Otros autores, sin embargo, sugieren interpretaciones alternativas que indican que los dos conjuntos de la sabana de Bogotá pertenecerían al mismo grupo cuyo repertorio lítico reflejaría economías de amplio espectro, y que la presencia de materia prima del Magdalena en la sabana revelaría relaciones intersocietales entre ambas regiones y redes comerciales amplias (Gnecco 2000; Nieuwenhuis 2002). Delgado (2016a) mostró que la presencia de individuos con dietas mixtas C3/C4 en momentos tempranos podría interpretarse como un indicador de grupos móviles foráneos o bien de relaciones entre grupos de la sabana y de tierras bajas. Asimismo, la relación entre los conjuntos líticos abriense y del valle del Magdalena no puede confirmarse dada la ausencia de contextos en la última región antes de los 10.500 años 14C a. P. Aun así, López y Cano (2011) sugirieron la necesidad de considerar la industria tequendamiense como parte de un conjunto mayor y de una tradición mucho más amplia y compleja, que se desarrolló durante varios milenios en el valle del Magdalena y se expandió de manera más bien restringida hacia otras localidades, como la sabana de Bogotá. Las puntas de proyectil encontradas en los ríos Porce y Medellín sugieren, según Aceituno et al. (2013), que dichos restos fueron descartados por cazadores-recolectores del Magdalena durante exploraciones en busca de nuevos territorios y recursos durante la frontera Pleistoceno/Holoceno, un periodo muy inestable en términos ambientales y climáticos, que afectó profundamente la distribución de los recursos (Van der Hammen y Hooghiemstra 1995).
Las relaciones entre el área central, integrada por el altiplano cundiboyacense, el valle del Magdalena y el suroccidente de Colombia, se ven opacadas por la escasez de información, aunque la evidencia proveniente de Pubenza y El Jordán probablemente indica que otros grupos habitaban en la región a fines del Pleistoceno (Correal et al. 2005; Salgado 1998). En términos generales, el registro arqueológico sugiere una conexión entre las cordilleras Oriental y Central, articuladas principalmente por el valle del Magdalena, pero con el uso de la cuenca entera y de pequeños valles interandinos que servían de conexión entre regiones a lo largo del gradiente altitudinal en ambas cordilleras (Aceituno et al. 2013; Delgado 2012c). Asimismo, en San Isidro, Gnecco (2000) recuperó lascas bifaciales, entre ellas varias puntas de proyectil manufacturadas en chert y obsidiana, las últimas con muchas similitudes morfológicas y tecnológicas a las de otros sitios del Ecuador, lo que indica posibles relaciones entre cazadores-recolectores andinos.
El incremento de la visibilidad arqueológica durante el Holoceno Temprano (ca. 10.000-8000 años 14C a. P.) refleja cambios poblacionales importantes, por ejemplo el aumento demográfico, lo cual habría permitido, de acuerdo con modelos como la geografía evolutiva (Lahr y Foley 1998), expansiones y dispersiones especialmente a lo largo de los Andes, donde se encuentra el grueso de los sitios. Las nuevas condiciones ambientales debido al incremento de la temperatura y la humedad permitieron la expansión de los bosques de tipo lluvioso principalmente y generaron, a su vez, múltiples respuestas dependiendo de las características cambiantes de los ecosistemas, como el manejo de recursos vegetales en el bosque tropical y premontano (Aceituno y Loaiza 2015; Cavelier et al. 1995; Delgado 2016a; Gnecco 2000; Santos, Monsalve y Correa 2015), el incremento en la caza de especies menores y la recolección de gasterópodos en la sabana de Bogotá (Correal 1990; Correal y Van der Hammen, 1977), y adaptaciones a ecosistemas de río en la cuenca del valle del Magdalena (López 2008; Otero y Santos 2002). La expansión de los rangos geográficos de diferentes poblaciones de cazadores-recolectores facilitó la compleja dinámica observada durante el Holoceno Temprano.
Finalmente, es interesante mencionar algunos aspectos sobre las inferencias hechas a partir de la evidencia lítica respecto a las estrategias de obtención de recursos de los cazadores-recolectores tempranos. El registro arqueológico del poblamiento sugiere la existencia de múltiples tradiciones tecnoculturales que reflejan básicamente adaptaciones a un amplio gradiente de escenarios ambientales, que van desde los relativamente especializados en fauna hasta aquellos enfocados en recursos vegetales. No obstante, interpretaciones recientes sugieren que las economías generalizadas o de amplio espectro, con la flexibilidad necesaria para explotar los diversos biomas existentes, seguramente fueron más importantes que las especializaciones (Borrero 2015; Delgado et al. 2015a; Delgado 2016a; Gnecco 2000). Cada vez son más las evidencias provenientes de diferentes regiones, como la sabana de Bogotá (Cárdenas 2002; Delgado 2016a), el Cauca Medio (Aceituno y Loaiza 2015; Dickau et al. 2015; Santos, Monsalve y Correa 2015), el Valle del Cauca (Salgado 1989-1990, 1998), el Valle de Pubenza (Gnecco 2000) y la cuenca del Amazonas (Morcote, Aceituno y León 2014), que corroboran dicha interpretación y resaltan la importancia de los recursos vegetales. Clásicamente, los grupos que habitaron el valle del Magdalena y la sabana de Bogotá han sido caracterizados como altamente móviles, enfocados básicamente en la caza de fauna mayor y menor con poca importancia de los recursos vegetales (Correal y Van der Hammen 1977; López 2008). Sin embargo, López y Cano (2011) sugieren que la relevancia de la explotación de recursos provenientes de ambientes boscosos y espacios ribereños indica economías de amplio espectro. Además, la supuesta especialización económica (i. e. caza de fauna mayor) en la sabana de Bogotá ha sido inferida a partir del estudio de conjuntos líticos, así como de la evidencia proveniente de un solo sitio (i. e. Tibitó). Esto es problemático ya que, por un lado, la industria tequendamiense no da cuenta de dicha especialización debido a su corta duración y, por el otro, la industria ampliamente distribuida en la región (i. e. abriense), presente en la mayoría de sitios, se corresponde con una tecnología unifacial simple, versátil y multifuncional que refleja posibilidades y restricciones ambientales más que especializaciones. Recientes análisis tecnofuncionales del repertorio lítico de la sabana de Bogotá (Nieuwenhuis 2002) corroboran esta inferencia, ya que sugieren el uso de herramientas abrienses en contextos y actividades no especializadas, así como la importancia de los recursos vegetales, tanto en un contexto dietario como tecnológico (i. e. manufactura de herramientas). Es interesante que análisis de isótopos estables de restos óseos humanos del Holoceno Temprano en esta región sugieran dietas basadas principalmente en recursos vegetales locales y, en menor medida, en proteína animal (Cárdenas 2002; Delgado 2016a). Contrario a interpretaciones anteriores, todo esto sugiere que los conjuntos líticos tempranos reflejan la existencia de economías de amplio espectro y dietas generalizadas entre los primeros habitantes.
El contexto bioantropológico
La evidencia genética
Con el estudio de marcadores moleculares de herencia biparental y uniparental (autosómicos, mitocondriales, cromosomas X e Y) en grupos amerindios contemporáneos y prehispánicos (figura 2), varios autores han sugerido hipótesis sobre el tiempo de entrada de los primeros pobladores, el número de dispersiones, las rutas de ingreso y la relación entre diversidad lingüística, geográfica y genética.
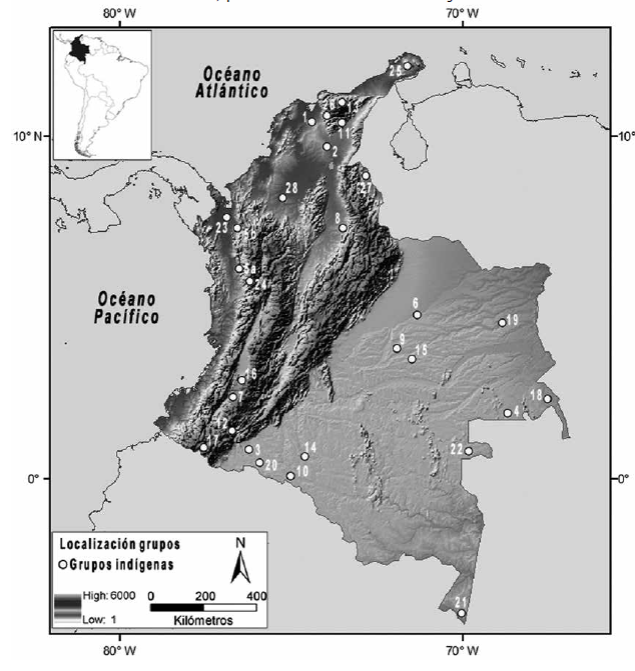
Fuente: Elaboración del autor empleando ArcGIS 9.3, licencia UNLP., a partir de la información publicada en Mesa et al. (2000); Keyeux et al. (2002); Torres et al. (2006); Wang et al. (2007); Usme-Romero et al. (2013)
Figura 2 Mapa de la distribución geográfica de grupos indígenas actuales de Colombia mencionados en el texto, que han sido estudiados desde evidencias genéticas Nota: 1) Arhuaco; 2) chimila; 3) coreguaje; 4) curripaco; 5a) emberá (Wang et al., 2007); 5b) emberá (Keyeux et al. 2002); 6) guahibo-sikuani; 7) guambiano; 8) guane-butaregua; 9) guayabero; 10) huitoto; 11) ijka-arhuaco; 12) inganos; 13) kogui; 14) murui-muinane; 15) nukak; 16) paez; 17) pastos; 18) piapoco; 19) piaroa; 20) siona; 21) ticuna; 22) tucano; 23) tule-cuna; 24) waunana; 25) wayuu; 26) wiwa; 27) yuko-yukpa; 28) zenú.
Diversidad molecular en grupos contemporáneos
A partir de los patrones de distribución geográfica de haplogrupos del ADNmt en grupos amerindios de Colombia (N = 25; véase la figura 2), Keyeux y Usaquén (2006) y Keyeux et al. (2002) propusieron un modelo que sugiere una importante diferenciación filogeográfica en la región, relacionada tanto con el aislamiento genético causado por la barrera geográfica de la cordillera de los Andes como con el ingreso de dos poblaciones fundadoras durante la frontera Pleistoceno/Holoceno. Una de ellas, proveniente de América Central, atravesó el istmo de Panamá y pobló la región noroccidental de Colombia (NO), integrada por la región andina y por las costas pacífica y atlántica. Otra población ancestral arribó desde el sur de Norteamérica por la cadena insular antillana y colonizó la región sudoriental (SO), conformada por las cuencas del Amazonas y del Orinoco. Aquellos grupos que habitan la región NO presentan altas frecuencias del haplogrupo A y una total ausencia del haplogrupo D, mientras que las que habitan la región SO exhiben frecuencias altas y medias de D y bajas frecuencias de A. A partir del estudio de haplogrupos del ADNmt en grupos chibcha-hablantes (koguis, arsarios e ijkas) y de lengua arawak (wayúus), Melton et al. (2007) encontraron un patrón de afinidades similar al propuesto por Keyeux et al. (2002). Asimismo, Melton et al. sugirieron que las diferencias entre grupos chibchas del noroccidente de Colombia y de Centroamérica pueden indicar una diáspora chibcha temprana (ca. 7000-10.000 a. P.), cuyo origen se remontaría a los 15.000 años a. P., según las fechas de coalescencia obtenidas.
Otros estudios basados en un espectro más amplio de marcadores (ADN-mt, autosómicos y del cromosoma-Y) sugieren la probable existencia de una, dos o más poblaciones fundadoras que habrían seguido una ruta de poblamiento única a través del istmo de Panamá y resaltan la relevancia de la costa pacífica en el proceso de dispersión inicial. Dichos grupos no presentarían una diferenciación clara a escala regional o de estructuración espacial a causa de los efectos de la deriva y el flujo génico, los cuales actuaron desde el Holoceno Temprano (Mesa et al. 2000; Ruiz-Linares et al. 1999; Torres et al. 2006). Ruiz-Linares et al. (1999) estudiaron varios marcadores del cromosoma-Y (microsatélites) y encontraron alta diversidad en la región, compatible con al menos dos poblaciones fundadoras. Asimismo, hallaron que la mayoría de los cromosomas DYS199C son autóctonos, lo que sugiere que dos haplotipos fundadores (DYS199T y DYS199C) estuvieron presentes en los pobladores iniciales y su amplia distribución en América señala que fueron introducidos por los fundadores del continente. Los análisis de coalescencia indicaron fechas con un rango muy amplio (DYS199T ca. 3700 y 13.000 años a. P. y DYS199C ca. 5200 y 18.600 años a. P.). En esa misma dirección, a partir de haplogrupos del ADNmt, Mesa et al. (2000) corroboraron varios de los hallazgos con base en el cromosoma Y (Ruiz-Linares et al. 1999). Esto significa concordancia entre los marcadores maternos y paternos y un índice similar de migración entre hombres y mujeres, aunque sin estructuración geográfica. Otro hallazgo importante a partir de análisis del ADNmt es la presencia de un linaje revertante del haplogrupo C en grupos del oriente de Colombia de lengua tucano-ecuatorial, que podría reflejar la acción de la deriva relacionada con el aislamiento poblacional posterior a la llegada de los grupos fundadores (Torres et al. 2006).
Usme-Romero et al. (2013) estudiaron la diversidad del ADNmt en Colombia en una gran cantidad de grupos amerindios (N = 21) con amplia distribución geográfica. Los autores encontraron alta diversidad genética y distribución clinal de los marcadores, donde el haplogrupo A posee altas frecuencias en el norte y bajas en el sur; el C y el D tienen un patrón contrario y el B es más frecuente en el occidente y disminuye hacia el oriente. Los patrones de diversidad a nivel intrapoblacional reflejaron diferencias entre el norte y el sur de Colombia, respectivamente. El estudio sugiere alta diversidad antes del ingreso de los fundadores a Sudamérica y llama la atención sobre procesos de diferenciación a nivel intrarregional y continental, al igual que el escaso rol de la cadena montañosa de los Andes en la diferenciación observada.
Diversidad molecular entre grupos prehispánicos (ADN antiguo)
Algunos autores han obtenido información genética (básicamente del ADNmt) de muestras prehispánicas tardías de Colombia (Barreto et al. 2007; Casas-Vargas et al. 2011). Sin embargo, recientemente Díaz-Matallana et al. (2015, 2016) obtuvieron resultados confiables (i. e. secuenciación en laboratorios independientes, controles por contaminación, etc. de una muestra con un rango temporal amplio de la sabana de Bogotá. De acuerdo con los resultados, las muestras tardías de la región nororiental, del altiplano cundiboyacense y del valle del río Cauca presentaron frecuencias variables de los cuatro macrohaplogrupos americanos A, B, C y D (Barreto et al. 2007; Casas-Vargas et al. 2011). Casas-Vargas et al. (2011) reportaron coincidencias a nivel haplotípico entre guanes prehispánicos y grupos del noreste de Asia, cuya relación parental con los nativos americanos ha sido establecida previamente y su alta diversidad ha sido explicada por medio de altos índices de flujo génico con grupos muiscas. Los análisis de biodistancias mostraron un patrón geográfico similar al actual, es decir, diferenciación regional y alta diversidad. Con respecto a la muestra temprana, Díaz-Matallana (2015) y Díaz-Matallana et al. (2016) hallaron cuatro haplotipos del ADNmt, entre ellos A2, B2, C1 y D4h3a. Tomando en cuenta otras líneas de evidencia, proponen un modelo de poblamiento que resalta la importancia de la costa pacífica, los valles interandinos y rutas ribereñas como vías de expansión, así como una continuidad genética desde momentos tempranos. Estos resultados descartarían, al menos durante el lapso considerado, componentes genéticos extraamericanos en la región en tiempos prehispánicos (contraNeves, Hubbe y Correal 2007).
La evidencia bioarqueológica
El estudio bioarqueológico de restos óseos humanos desde diferentes perspectivas metodológicas y poblacionales ha generado también algunas ideas interesantes en torno a la problemática discutida. Así, a partir del estudio de los patrones de diversidad morfológica a nivel dental y craneofacial entre grupos prehispánicos (figura 3), se han propuesto varias hipótesis de poblamiento y microevolución.
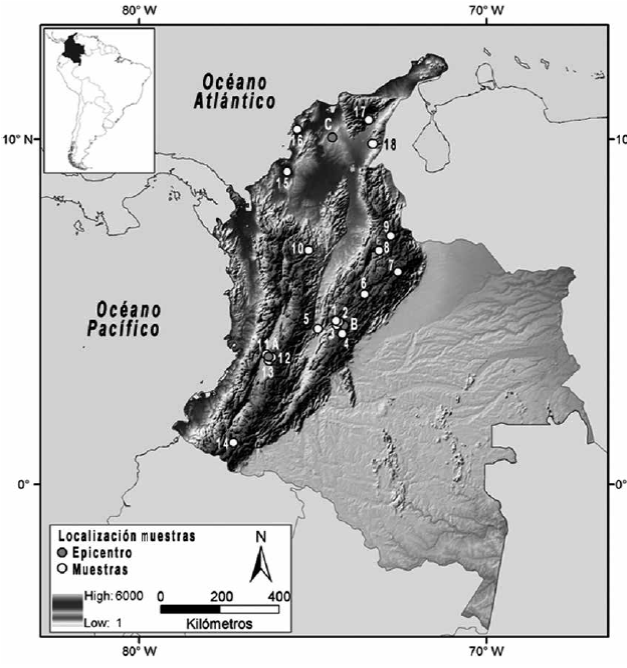
Fuente: Elaboración del autor empleando ArcGIS 9.3, licencia UNLP., a partir de Delgado (2015)
Figura 3 Mapa de la distribución geográfica y temporal de algunas de las muestras osteológicas comúnmente investigadas en estudios bioarqueológicos Nota: 1) Holoceno Temprano/Medio, sabana de Bogotá; 2) Holoceno Medio, sabana de Bogotá; 3) Holoceno Tardío Inicial, sabana de Bogotá; 4) Holoceno Tardío Final; 5) Panche; 6) Muiscas Boyacá; 7) Lache; 8) Guane; 9) Chitarero; 10) Porce (Holoceno Temprano/Medio); 11) Guacarí; 12) El Cerrito; 13) Palmira (Coronado, Estadio Deportivo Cali; Malagana; Santa Bárbara); 14) Nariño; 15) Cueva La Trementina; 16) Turbaná; 17) Sierra del Perijá; 18) Gamarra. Las letras A, B y C corresponden a los epicentros de la distribución de algunas muestras dispersas en una región específica empleados en análisis estadísticos (véase Delgado 2012c; 2015).
Los estudios iniciales
A pesar de que Paul Broca (1876) realizó uno de los primeros estudios con material esquelético prehispánico de la región, los estudios sistemáticos se remontan a la década de 1940, cuando Paul Rivet (1943) analizó varias muestras y sugirió la existencia de una alta diversidad biológica compatible con múltiples orígenes, algunos ubicados fuera del continente. Correal y Van der Hammen (1977) analizaron la morfología craneofacial en muestras tempranas de la sabana de Bogotá y sugirieron una única población ancestral diversa culturalmente que arribó a Colombia y se dispersó por diferentes regiones. Estos autores indican que la historia poblacional de la sabana de Bogotá es compleja, es decir, que hay diferencias notables desde el Pleistoceno Final y cambios importantes durante el Holoceno Temprano relacionados con fluctuaciones climáticas. Solo a partir de este momento ocurre un importante incremento demográfico y expansiones poblacionales hacia diferentes áreas, como las cordilleras Central y Occidental y el suroccidente de Colombia. Por su parte, Sacchetti (1988) analizó la variabilidad craneofacial de grupos prehispánicos en Colombia a nivel estadístico y, desde un enfoque tipológico, propuso hipótesis sobre el origen de la variabilidad de corte dispersionista. Según este autor hubo diferentes demotipos (i. e. linajes) que indican el ingreso de múltiples grupos a la región, provenientes de Centroamérica así como del Caribe y del Orinoco. También sugiere una diferenciación entre grupos tempranos “precerámicos” y tardíos agrícolas, según la cual los primeros generaron parte de la variabilidad observada posteriormente, pero complementada con expansiones locales y extrarregionales.
Estudios recientes
Desde un enfoque teórico-metodológico más sofisticado, algunos investigadores adelantaron estudios que pueden enmarcarse dentro de una perspectiva bioarqueológica contemporánea. En distintos trabajos, J. V. Rodríguez y colegas han propuesto un escenario de poblamiento conservador de evolución gradual, con base en los patrones de variación a nivel craneofacial y dental en muestras prehispánicas de la región (J. V. Rodríguez 2001, 2007; Rodríguez y Vargas 2010; Vargas 2010; para una opinión contraria, véase Delgado 2015, 2016b). Los aspectos más relevantes de dicho modelo se resumen a continuación: 1) grupos de cazadores especializados pertenecientes a una única población ancestral poco diversa, procedentes del istmo de Panamá, ingresaron y se expandieron por toda la región a través de valles interandinos durante el Pleniglacial (ca. 26.000-14.000 años 14C a. P.); 2) entre ca. 14.000-12.000 años 14C a. P., grupos de cazadores especializados en megafauna provenientes del valle del Magdalena arribaron a la sabana de Bogotá y se asentaron en los abrigos rocosos de El Abra y Tequendama; 3) las muestras más tempranas de esta región que presentaban un patrón craneofacial paleoamericano serían las representantes de la población antecesora que se habría dispersado por el valle del río Sogamoso-Chicamocha y por la sabana de Bogotá a través del valle del río Bogotá. Después, al traspasar la cordillera Central, se habrían expandido hacia el valle del río Cauca y el sur del país, y 4) en la región las poblaciones evolucionaron in situ con pocas modificaciones.
Por otro lado, Neves, Hubbe y Correal (2007), mediante el estudio de algunas colecciones óseas de la sabana de Bogotá, formularon hipótesis sobre las fuentes de diversidad, el número de poblaciones fundadoras y la divergencia morfológica (para una opinión contraria, véase Delgado 2015, 2016b). Neves, Hubbe y Correal (2007) analizaron 52 cráneos procedentes de 7 localidades, cuya cronología fue dividida entre paleocolombianos (11.000-6000 años 14C a. P.) y colombianos arcaicos (5000-3000 años 14C a. P.). Para la presente discusión, los hallazgos más interesantes son la existencia de marcadas similitudes entre paleoamericanos de Brasil, australomelanesios, africanos y europeos modernos, y los paleocolombianos y colombianos arcaicos. Esto sugeriría el poblamiento de la región por dos grupos biológicos distintos y una mayor dispersión geográfica de la morfología paleoamericana; es decir, los grupos tempranos son parte de los llamados paleoamericanos, cuya morfología generalizada debió originarse fuera de América. También detectaron una continuidad morfológica de 8.000 años desde el 11.000 hasta el 3000 14C a. P., cuando arribaron grupos con una morfología craneal distinta denominada amerindia. Esta transformación no habría sido producto de una adaptación a cambios ambientales o en la economía de subsistencia, sino de una dispersión proveniente de distintas regiones, fundamentalmente del valle del Magdalena, que reemplazó (o asimiló) a las poblaciones preexistentes.
Mediante el análisis de la morfología dental de muestras prehispánicas de la región, D. Rodríguez (2012) y Rodríguez y Colantonio (2015) realizaron algunas interpretaciones del proceso de poblamiento, según las cuales hubo más diversidad dental que la explicada por un origen único a escala regional y existieron diferencias marcadas respecto al patrón dental a nivel cronológico. Asimismo, hubo diferentes procesos de discontinuidad poblacional básicamente entre grupos del Holoceno Temprano/Medio, Medio y Tardío Inicial y una continuidad durante el Holoceno Tardío Final.
Recientemente, desde diferentes enfoques metodológicos, estadísticos y teóricos que abordan múltiples líneas de evidencia, Delgado (2012a, 2012b, 2012c, 2015, 2016a, 2016b) indagó sobre el poblamiento y la expansión poblacional ocurrida durante la frontera Pleistoceno/Holoceno y esbozó algunas ideas que se resumen a continuación. La evidencia morfológica craneofacial y dental correspondiente al Holoceno Temprano (10.000-7000 años 14C a. P.) sugiere la existencia de una única población fundadora relativamente diversa, con un patrón morfológico craneofacial afín al denominado paleoamericano o al extremo generalizado del vector morfológico americano (Delgado 2016b) y rasgos dentales típicamente amerindios (Delgado 2015). La escasez de restos humanos antiguos fuera del altiplano cundiboyacense -así como su dispar estado de conservación- impide establecer el grado de diferenciación geográfica alcanzado por las poblaciones locales derivadas del único stock fundador. No obstante, diferencias detectadas entre los grupos del Holoceno Temprano/Medio de las cordilleras Central (Porce) y Oriental, respectivamente, indican que es probable el ingreso a la región de más de una población fundadora o, de manera alternativa, la existencia de mayor diversidad biológica en la única que arribó.
El empleo de diferentes métodos genético-cuantitativos para inferir flujo génico, deriva y selección natural (Delgado 2012a c, 2015) señala una variación moderada durante el Holoceno Temprano, y mayor varianza fenotípica que la esperada, por el surgimiento de flujo génico y posiblemente la expansión de más de una población fundadora. Estos resultados concuerdan con la evidencia arqueológica y paleoambiental que indica un aumento demográfico y mayores relaciones intersocietales coincidentes con mejoras climáticas. La evolución morfológica ocurrida durante el Holoceno Temprano no se relaciona con la acción de presiones selectivas en respuesta a cambios ambientales o en la dieta, sino con deriva y flujo génico fundamentalmente.
Discusión: hacia un modelo unificado de poblamiento
En su conjunto, las diferentes líneas de evidencia consideradas sugieren un proceso de poblamiento más complejo que el asumido previamente. Respecto al tiempo del ingreso inicial, si bien existen datos que indican fechas superiores a los 12.000 años 14C a. P., las evidencias más fuertes señalan que la población fundadora alcanzó una notable visibilidad arqueológica después de los 11.000 14C a. P. Desde el punto de vista de la geografía cultural (Borrero 2015), las diferencias pre y pos 11.000 14C a. P. observadas pueden indicar diferentes fases de poblamiento correlacionadas con eventos paleoclimáticos; la más temprana correspondería a la fase de exploración sucedida durante el Interstadial Guantiva o antes y la más tardía, a la fase de colonización propiamente dicha, ocurrida durante el Estadial El Abra. Solo durante el Holoceno Temprano se habría dado lo que Borrero llama ocupación efectiva, relacionada con la ocupación estable del espacio, lo cual también coincide con una mejora importante en las condiciones ambientales. Diferentes archivos polínicos indican un periodo de incremento de temperatura y humedad en comparación con periodos anteriores (Marchant et al. 2002).
En el contexto del poblamiento inicial de la región, el uso de modelos teóricos como la geografía evolutiva y la biogeografía humana (Harcourt 2012; Lahr y Foley 1998) puede ser útil para comprender mejor la importancia de los cambios ecológicos y ambientales pasados como promotores de una amplia cantidad de cambios a escala poblacional; entre ellos, la retracción/extinción y la expansión/dispersión que también generan múltiples respuestas a nivel biológico y cultural. Este marco teórico por supuesto no sugiere una relación simplista y directa entre el cambio ecológico/ambiental y biocultural, sino que revela una compleja red de interacciones entre el ser humano y el medio en diferentes niveles y escalas que deben ser vistos en su conjunto como un proceso coevolutivo y adaptativo (Delgado et al. 2015a). Esto permitiría explicar tanto los incrementos demográficos (e. g. radiación espacial) como los procesos de discontinuidad y diferenciación poblacional ocurridos durante el Pleistoceno Final, pero fundamentalmente durante el Holoceno Temprano. Para ese momento hay un marcado aumento en la cantidad de restos óseos humanos, así como en la señal arqueológica, lo cual indica un importante incremento demográfico y la complejización de la dinámica de poblamiento a escala regional, con mayor cantidad de grupos que se expanden por el territorio (Aceituno et al. 2013; Delgado et al. 2015a; Delgado et al. 2015b). El registro arqueológico muestra grupos foragers tempranos distribuidos por una amplia cantidad de escenarios ambientales que integran la sabana de Bogotá, el valle del Magdalena, el Cauca Medio, el Valle del Cauca, el altiplano de Popayán y la cuenca del Amazonas, con una impresionante diversidad tecnológica y diferentes tendencias económicas (Aceituno y Loaiza 2015; Aceituno et al. 2013; Delgado et al. 2015a; Delgado et al. 2015b; Delgado 2016a; Correal y Van der Hammen 1977; López 2008; Mora y Gnecco 2003; Niuwenhuis 2002).
Esta expansión holocénica también señala la posibilidad de contactos poblacionales a diferentes escalas y, por lo tanto, de intercambio genético (i. e. flujo génico), lo cual explicaría la supuesta homogeneidad biológica vista durante momentos tempranos (Delgado 2015; J. V. Rodríguez 2007). La evidencia genética no tiene aún la resolución necesaria para sugerir fechas de ingreso acotadas; no obstante, varios estudios proponen fechas de coalescencia que oscilan entre los 13.000 y 16.000 años 14C a. P., lo cual se corresponde relativamente con el registro arqueológico más temprano (Ruiz-Linares et al. 1999; Díaz-Matallana 2015). Asimismo, algunos estudios que emplearon métodos de genética cuantitativa en la evaluación del tiempo necesario para la generación de la diversidad morfológica vista sugieren fechas de ca. 13.000 14C a. P. para el ingreso inicial de humanos a la región (Delgado 2015, 2016b).
De acuerdo con la evidencia disponible, resulta difícil establecer si ingresaron uno o varios grupos. Algunos autores sugieren que la impresionante diversidad cultural temprana se generó a partir de una única población ancestral (Correal y Van der Hammen 1977; Correal 1990), aunque otros proponen que probablemente más grupos ingresaron en diferentes momentos, dada la importante variación de sistemas de subsistencia, tecnología lítica, rangos geográficos de acción, entre otros (Aceituno y Rojas 2015; Aceituno et al. 2013; Delgado et al. 2015a, 2015b; Delgado 2016a, 2016b; López y Cano 2011; Ranere y López 2007). Si bien algunas de estas diferencias pueden resultar de adaptaciones a entornos ambientales y sociales distintos, muchas son contemporáneas, lo que da más apoyo a la idea de múltiples grupos con diversos patrones culturales. Otras líneas de evidencia también sugieren propuestas contradictorias; de acuerdo con la evidencia genética, ingresaron una (Usme-Romero et al. 2013), dos (Ruiz-Linares et al. 1999; Keyeux et al. 2002) o más (Mesa et al. 2000; Torres et al., 2006) poblaciones ancestrales. Aun así, la mayoría proponen más de un grupo fundador, lo cual coincide con la interpretación del registro arqueológico.
Los estudios que abordaron el registro bioarqueológico presentan una significativa limitante para proporcionar datos sobre la cantidad de grupos que ingresaron, ya que la evidencia más temprana proviene de una única región (i. e. la sabana de Bogotá), lo cual no permite evaluar diferencias a nivel geográfico. No obstante, algunos autores señalan que la amplia diversidad dental vista durante el Holoceno Temprano sugiere más de un grupo fundador (Delgado 2012b, 2012c; D. Rodríguez 2012; Rodríguez y Colantonio 2015). Delgado (2012b, 2012c, 2015, 2016b) mostró que, si bien no es posible descartar el ingreso de una única población fundadora a partir de la evidencia disponible, diferencias halladas entre grupos tempranos de las cordilleras Central (i. e. Porce) y Oriental, así como el hallazgo de varianza fenotípica moderada durante el Holoceno Temprano/Medio, indicarían el ingreso de una población fundadora más diversa o, alternativamente, el arribo de varias poblaciones con variados patrones de diversidad fenotípica. Estos análisis señalan igualmente que dichas diferencias se relacionan con la diversidad generada antes del ingreso a la región, e incluso al continente.
El patrón de expansión y de relaciones a escala intra y extrarregional de los grupos cazadores-recolectores tempranos es complicado de establecer dado lo fragmentario del registro arqueológico, aunque en conjunto esta y otras evidencias genéticas, lingüísticas y bioarqueológicas proporcionan una idea sobre dicho proceso. Como se mostró arriba, a partir del análisis de los conjuntos líticos, existió una compleja dinámica de poblamiento. La relación establecida entre el valle del Magdalena y las cordilleras Central y Oriental, pero no con la cordillera Occidental, indicaría, por un lado, una dinámica de expansión multidireccional occidente-oriente, oriente-occidente y norte-sur y, por el otro, probablemente la existencia de barreras geográficas o culturales. Los estudios lingüísticos llevados a cabo por Dahl et al. (2011) podrían corroborar las relaciones mencionadas a partir de los conjuntos líticos, ya que sugieren procesos de dispersión de algunos troncos lingüísticos coincidentes con otras evidencias. Por ejemplo, mientras que la familia chibcha se habría expandido a través del valle del Magdalena, la lengua paez lo habría hecho a través del Valle del Cauca, lo que produjo, según estos autores, la divergencia vista entre los grupos ubicados en ambos valles interandinos. La diferenciación genética y lingüística sugerida por Keyeux et al. (2002) no parece ser corroborada por el registro arqueológico, ya que no existen diferencias en la distribución espacial ni relaciones tecnológicas que sugieran la dispersión de dos poblaciones ancestrales dicotómicamente distribuidas en la región andina y las cuencas del Orinoco/Amazonas; aunque de alguna forma la cordillera andina parece haber representado una barrera permeable durante la fase de poblamiento inicial. Por lo tanto, un patrón de expansión poblacional temprano en dirección nororiental o suroccidental no puede ser corroborado. La divergencia morfológica a nivel dental entre grupos tempranos de las cordilleras Oriental y Central (Delgado 2015) daría soporte a la diferenciación observada a nivel tecnológico y económico entre ambas regiones (Aceituno y Rojas 2015; Aceituno et al. 2013), aunque es claro que las similitudes culturales no necesariamente indican semejanzas biológicas y viceversa. Finalmente, la distribución espacial del registro arqueológico temprano (e. g. cronologías tempranas en el altiplano cundiboyacense y el valle del Magdalena frente a ocupaciones tardías en el Valle del Cauca y la cuenca del Amazonas) sugiere que el proceso de dispersión dentro de la región fue relativamente lento y multidireccional, lo que concuerda con la teoría de colonización de paisajes desconocidos donde se requiere tiempo para conocer un territorio y explotarlo (Hazelwood y Steele, 2003; Meltzer 2003).
Perspectivas futuras
Si bien existen notables avances en la investigación antropológica del primer poblamiento de la región, también hay aspectos que impiden obtener una idea más clara y detallada del proceso. La investigación arqueológica continua en el Cauca Medio (Aceituno y Loaiza 2015; Dickau et al. 2015), la cuenca del Amazonas (Morcote, Aceituno y León 2014) y el valle del Magdalena (López y Cano 2011) proporciona información importante sobre la subsistencia y adaptación de los primeros habitantes a diversos ecosistemas. Asimismo, la reevaluación de la evidencia disponible ha generado aportes en torno a la estructura espaciotemporal del poblamiento y el tiempo de llegada de los grupos tempranos (Delgado et al. 2015b), así como su utillaje lítico y formas de captación de recursos (Aceituno y Rojas 2015; Delgado 2016a). Aun así, en el corto plazo resulta absolutamente necesario enfocar la investigación hacia contextos espaciales relevantes pero poco investigados, como la costa caribe y las cuencas del Amazonas y el Orinoco. También es importante continuar con el estudio de diferentes regiones que han sido clásicamente investigadas, como la sabana de Bogotá, el Valle del Cauca y el altiplano de Popayán. Adicionalmente, la reevaluación de los materiales recuperados hace décadas también debe ser prioridad, incluyendo nuevas dataciones de materiales con la técnica de AMS, preferiblemente de restos óseos humanos, fauna y de elementos que tengan claras huellas de manipulación humana para mejorar la resolución cronológica.
El estudio de la evidencia bioarqueológica ha mejorado sustancialmente con el empleo de enfoques estadísticos, poblacionales y teóricos más robustos y sofisticados (Delgado 2015; Díaz-Matallana 2015; D. Rodríguez 2012; Vargas 2010). Sin embargo, el uso de nuevas técnicas, como análisis de morfometría geométrica en dos y tres dimensiones, de genética cuantitativa y simulaciones a nivel microevolutivo, puede arrojar más información. También se debe estandarizar la utilización de métodos y técnicas numéricas y estadísticas más apropiadas para el estudio de restos óseos antiguos, empleando enfoques exploratorios, correlacionales y explicativos, así como la aplicación de técnicas para inferir la estructura poblacional que se basan en la teoría genética cuantitativa (véase Delgado 2016b). Se requieren nuevos análisis de isótopos estables, enfocados no solo en la dieta sino en la movilidad, como isótopos de oxígeno y estroncio para obtener información detallada de las formas de dispersión y expansión intrarregional.
Todo lo mencionado en este trabajo, tanto las fortalezas como las falencias, sugiere que el estudio del poblamiento inicial del noroccidente de Sudamérica es un campo de investigación fértil y prometedor, tal y como lo indica la publicación de volúmenes recientes sobre la arqueología y la bioantropología del poblamiento inicial a escala nacional (Delgado y Rosique 2012) e internacional (Delgado et al. 2015a).

