










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkUniversitas Psychologica
versão impressa ISSN 1657-9267
Univ. Psychol. v.5 n.2 Bogotá jan./maio 2006
ALMA GABRIELA MARTÍNEZ, ANTONIO LÓPEZ-ESPINOZA Y HÉCTOR MARTÍNEZ*
UNIVERSIDAD DE GUADALAJARA
almagabrielamm@yahoo.com.mx anton779@megared.net.mx
ABSTRACT
Six albino rats were divided in two experimental groups and one control. The experiment began with fifteen days of free access; subsequently experimental groups maintained available three concentrations of glucose: high, middle and low. First group received high-middle-low-high-middle-low sequence and second group received low-middle-high-low-middle-high sequence during six days. Control group not received glucose concentrations. Caloric concentration of food never changed. Results suggest that modification of caloric concentration in water affects feeding behavior. Nevertheless, water with glucose consumption did not change body weight.
Keywords: Glucose, body weight, water and food consumption, calories, rats, feeding behavior.
RESUMEN
Seis ratas Wistar se dividieron en dos grupos experimentales y uno control. Se inició con quince días de libre acceso, posteriormente los sujetos experimentales recibieron tres concentraciones de agua con glucosa denominadas alta, media y baja. El primer grupo recibió la secuencia: alta-media-baja-alta-mediabaja y el segundo grupo la secuencia baja-media-alta-baja-media-alta durante seis días. El grupo control no recibió glucosa. Los resultados mostraron que variar el contenido calórico del agua modifica la conducta alimentaria. No se observaron cambios en el peso corporal a pesar de los grandes consumos de agua con glucosa.
Palabras clave: Glucosa, peso corporal, consumo de alimento, consumo de agua, calorías, ratas, conducta alimentaria.
Introducción
La evidencia experimental disponible demuestra que: a) los organismos son capaces de ajustar su ingesta calórica en presencia de alimentos y comidas con diferente contenido energético (Collier, Hirsh & Kanareck, 1983; Dimitrou, Rice & Corwin, 2000), y, b) que el peso corporal presenta un aumento consistente ante dietas altamente palatables o con niveles elevados en la cantidad de grasa (Beatty, 1978; Treit, Spetch & Deustch, 1982).
De manera particular, aún no es posible conocer los mecanismos por los cuales las ratas equilibran exitosamente su consumo de alimento en situaciones de elección ante dietas de cafetería, pues los resultados obtenidos hasta hoy son contradictorios. Por un lado, se ha señalado que los animales no son capaces de adecuarse acertadamente a las demandas de su ambiente en situaciones particulares de alimentación (por ejemplo ante cambios abruptos en su contexto) (Galef, 1996, Woods & Seeley, 2003). Por otro lado, también se han reportado algunos estudios que demuestran que las ratas son capaces de seleccionar aquellas comidas que convienen a sus necesidades energéticas (Bolles, 1983; Collier, Hirsh & Kanareck, 1983; Corwin, Wojnicki, Fischer, Dimitrou, Rice & Young, 1998; Dimitrou, Rice & Corwin, 2000).
Teiltelbaum y Campell (1958) señalaron que el registro del número y tamaño de las comidas ha demostrado que las ratas consumen entre 50 y 60 calorías diariamente. Por su parte, Collier, Hirsch y Kanareck (1983) demostraron que el consumo de calorías se mantiene constante a pesar de exponer a los sujetos experimentales a una variedad de alimentos.
Diversos autores han afirmado que la periodicidad del patrón alimentario en la rata es producto de la regulación de energía (Collier, Hirsh & Kanareck, 1983; Davis & Levine 1977; Keesey, 1986; Lane, Ingram & Roth, 1999; Mayer, 1955; Staddon & Zanutto, 1998; Staddon, 2003). Tanto Weingarten (1990) como Strubbe y Woods (2004) señalaron que la comprensión del fenómeno de la regulación de energía desde una perspectiva conductual, debe enfocarse en determinar cómo el organismo controla y distribuye la conducta de comer para mantener su homeostasis y sobrevivir. Por su parte, Galef (1996) sugirió que no es del interés psicológico si los humanos o las ratas son capaces de seleccionar dietas adecuadas, sino que el interés debe enfocarse en reportar las características de las situaciones de elección en sus patrones de alimentación. A partir de estos planteamientos se han elaborado numerosos estudios que han evaluado el patrón alimentario de la rata a partir de la disponibilidad de alimentos con diferente contenido calórico bajo diferentes condiciones experimentales (Capaldi, Campell, Sheffer & Bradford, 1987; Young & Shuford, 1955). Sin embargo, no está claro cuál es el efecto de modificar el contenido energético del agua sobre el patrón alimentario de las ratas.
Con base en este cuestionamiento nos propusimos evaluar experimentalmente los efectos de modificar el contenido energético del agua sobre el peso corporal y el consumo de alimento, agua y calorías. Para alcanzar este objetivo se utilizaron tres soluciones de agua con glucosa. Se utilizó la glucosa porque además de tener un sabor altamente palatable su contenido calórico es muy estable lo que permite un control experimental adecuado. La condición de libre acceso fue el procedimiento empleado durante todo el experimento. El interés principal consistió en evaluar el efecto de incluir la glucosa en la dieta sobre el patrón alimentario de la rata y valorar si alguna o las tres concentraciones de glucosa en agua eran aceptadas o rechazadas por los animales. Es por ello que los sujetos experimentales tuvieron disponibles cada una de las tres concentraciones de glucosa en el agua. Se esperaba que los sujetos disminuyeran su consumo de alimento a partir del número de calorías consumido en cada concentración de glucosa, manteniendo una estabilidad en el peso corporal de acuerdo a la regulación de energía. Adicionalmente, se esperaba encontrar modificaciones de la conducta alimentaria manifestada en cambios sobre el peso corporal y los consumos de agua, alimento y calorías.
Método
Sujetos
Seis ratas de la cepa Wistar, tres machos (ML1, ML2, MC) y tres hembras (HL1, HL2, HC), con 4 meses de edad al inicio del estudio e ingenuas experimentalmente.
Aparatos y materiales
Se utilizaron seis cajas de habitación individual, con medidas de 13 cm de altura por 27 cm de ancho y 38 cm de largo, con una reja metálica en la parte superior con división para comedero y bebedera. Para el registro del consumo de alimento y peso corporal se utilizó una báscula electrónica de precisión. Croquetas de la marca comercial Nutri-cubos, especial para animales de laboratorio, fue el alimento proporcionado. Su fórmula nutricional es la siguiente: 3% de grasas, 7% de cenizas, 1% de calcio, 23% de proteína, 6% de fibra, 49% de E.L.N. (extracto libre de nitrógeno), 6% de fósforo y 12% de humedad.
Como bebida se utilizó agua y durante la manipulación experimental se proporcionó una solución compuesta de glucosa diluida en agua en tres diferentes concentraciones: alta, media y baja. La concentración alta (A) fue de 45g de glucosa diluidos en 200ml de agua, que aportó 90 calorías en total; la concentración media (M) fue de 30g de glucosa en 200ml con 60 calorías, y la concentración baja (B) se conformó por 15g de glucosa por cada 200ml de agua con 30 calorías. El agua sin glucosa proporcionó 0 calorías. El líquido se proporcionó en bebederos graduados de 200ml.
Procedimiento
Los sujetos fueron identificados con un número de registro, fecha de nacimiento y peso corporal al inicio del experimento y fueron colocados en cajas habitación individual. El peso corporal, el consumo de agua y comida se registró diariamente a las 9:00 de la mañana. Para registrar el peso corporal se tomaba la caja habitación y se trasladaba a la mesa de trabajo en la que se encontraba la báscula utilizada para el registro. Se tomaba a la rata y se introducía en el recipiente de la báscula. Al finalizar este procedimiento los sujetos retornaban a su caja habitación y permanecían en el bioterio.
Para calcular las calorías consumidas en el agua con glucosa y en el alimento se determinó el número de calorías que aporta el alimento por gramo a partir de la media estandarizada en nutrición animal (Subcommittee on Laboratory Animal Nutrition, Committee of Animal Nutrition, Board on Agriculture, National Council, 1995) de 3,4 calorías por gramo consumido. Esta cantidad se multiplicó por el número de gramos consumidos diariamente por el animal. El agua proporcionó 0 calorías. Cuando se adicionó glucosa al agua, se calculó el número de calorías que aportaba cada concentración de glucosa, el cual se multiplicó por el número de mililitros consumidos diariamente (Tabla 1). Se sumó la cantidad de calorías consumidas en el alimento más el número de calorías consumidas en la bebida para obtener el consumo total diario de calorías.
Diseño experimental
Se formaron dos grupos de sujetos experimentales y uno control, cada grupo se integró con una hembra y un macho. Los sujetos ML1 y HL1 fueron asignados al primer grupo, los sujetos ML2 y HL2 se asignaron al segundo grupo y los sujetos MC y HC al tercer grupo (control). La asignación de todos los sujetos a los grupos experimentales se realizó de forma aleatoria.El experimento se dividió en siete fases. Las fases 1, 3, 5 y 7 tuvieron una duración de quince días cada una. En estas fases se proporcionó 50g de nutricubos (chow) y 200ml de agua sin glucosa a todos los sujetos. Las fases 2, 4 y 6 tuvieron una duración de seis días cada una y proporcionaron 50g de nutricubos (chow) y 200ml de agua con glucosa en sus tres tipos de concentración a los sujetos experimentales. El grupo 1 recibió la secuencia de agua con glucosa de la siguiente manera altamedia- baja-alta-media-baja, durante las fases 2, 4 y 6. El grupo 2 recibió la secuencia opuesta, bajamedia- alta-baja-media-alta durante las fases 2, 4 y 6. El grupo control recibió durante estas fases 50g de nutricubos (chow) y 200ml de agua, es decir, se mantuvo en condiciones de libre acceso sin glucosa durante todo el experimento.
Resultados
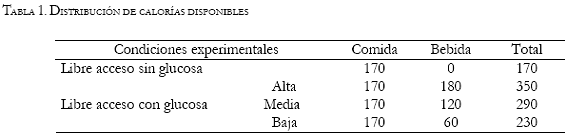
Las Figuras 1, 2, y 3 muestran los datos individuales del registro diario del peso corporal, del consumo alimento y del consumo de agua respectivamente. Las líneas continuas representan las fases de línea base y libre acceso sin glucosa. Las fases de libre acceso con glucosa están representadas por tres figuras, correspondientes a cada una de las tres concentraciones proporcionadas. El círculo representa la concentración alta de glucosa (45g por cada 200ml), el cuadrado representa la concentración media (30g por cada 200ml) y el triángulo la concentración baja (15g por cada 200ml). Los dos paneles superiores muestran los datos del grupo 1, los centrales el grupo 2, y los inferiores al grupo control. Los datos de los machos y las hembras de cada grupo se ubican en las columnas izquierda y derecha respectivamente. La Figura 4 muestra el promedio de consumo de calorías durante cada fase del experimento.
Las gráficas de la Figura 1 muestran el peso corporal de todos los sujetos. Los sujetos experimentales y los controles presentan una curva de crecimiento del peso corporal similar. Las fases en las que se adicionó la glucosa no muestran diferencias respecto a las fases de libre acceso sin glucosa. Los sujetos ML2 y HL1 presentaron un decremento en el peso corporal en las fases 3 y 5 de libre acceso sin glucosa respectivamente, que posteriormente fue recuperado durante ese mismo periodo, para continuar con su patrón ascendente.
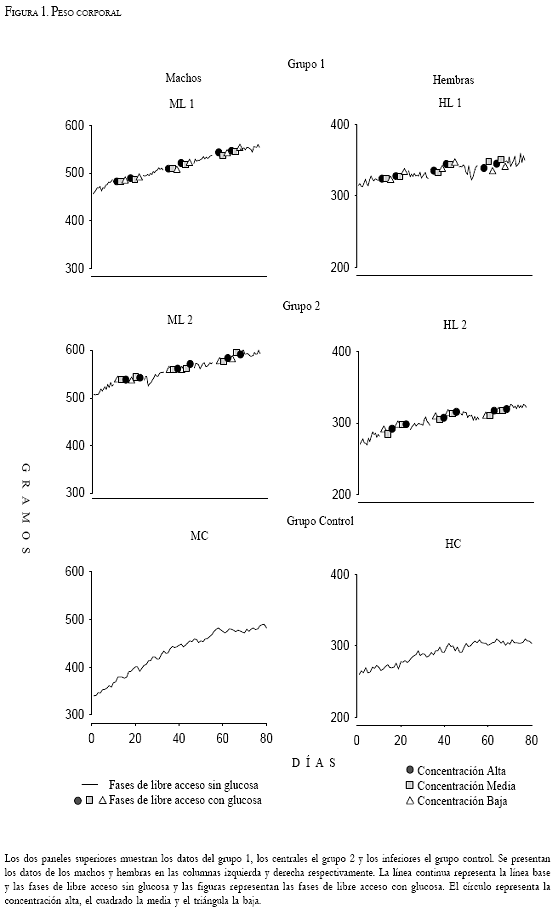
En la Figura 2 se presenta el consumo de alimento. Ambos grupos experimentales muestran diferencias durante las fases de línea base y de libre acceso sin glucosa respecto a las fases de libre acceso con glucosa. El consumo promedio durante la línea base se registró entre los 22 y 32g en los machos y entre los 15 y 20g en la hembra HL1. La hembra HL2 inició su consumo de alimento entre los 25 y 30g, este posteriormente disminuyó y se situó entre los 15 y 22g. Durante las fases 3, 5 y 7 se observó el mismo promedio de consumo, excepto en el sujeto ML2 cuyo consumo osciló de los 12 hasta los 32g, sobre todo en la fase 7. Cuando se adicionó glucosa al agua (fases 2, 4 y 6) el consumo de los sujetos experimentales disminuyó respecto al registrado durante la línea base y las fases de libre acceso sin glucosa. Los machos consumieron un promedio de 10, 15 y 18g y las hembras consumieron 9, 10 y 12g durante la concentración alta, media y baja respectivamente. El grupo control no presenta diferencias de consumo entre las fases, el macho consumió un promedio de 28g y la hembra 20g. Una característica particular del sujeto HC es que durante las cuatro primeras fases mostró una variabilidad de +/- 10g en el consumo de alimento, a partir de la fase 5 su consumo muestra una variabilidad de +/-5g.
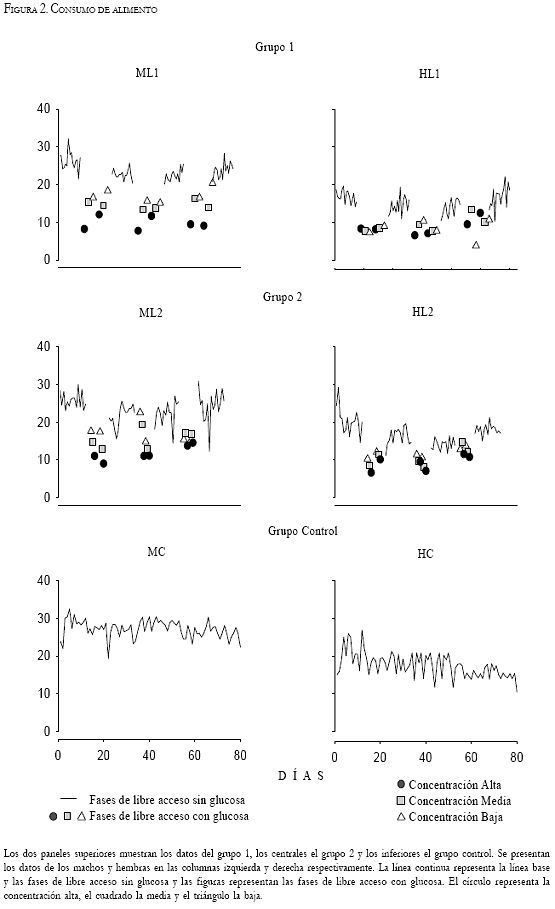
La Figura 3 muestra el registro del consumo de agua y de agua con glucosa. Durante la línea base y las fases de libre acceso sin glucosa se muestra una variabilidad en el consumo de agua de +/ -15ml en los machos y de +/-20ml en las hembras de los grupos experimentales, consumiendo un promedio de 60 y 45ml diarios respectivamente. Se observa un incremento en el consumo de agua durante las fases de libre acceso con glucosa respecto a las fases de libre acceso sin glucosa: el consumo se incrementó en promedio 20, 50 y 100ml durante la concentración alta, media y baja respectivamente. Los sujetos ML1 y HL2 duplicaron y los sujetos ML2 y HL1 triplicaron su consumo de agua con la concentración baja de glucosa respecto al promedio registrado durante las fases de libre acceso sin glucosa. El grupo control no mostró diferencias de consumo a lo largo de todo el estudio. El macho consumió entre 50 y 60ml y la hembra entre 40 y 50ml.
Figura 4. Se muestra el consumo de calorías del grupo 1 (panel superior) y 2 (inferior). Las barras negras representan el promedio de consumo durante las fases 1, 3, 5 y 7. El conjunto de barras bicolores representa el consumo de calorías en el alimento (negro) y en el agua (blanco) durante los seis días de las fases 2, 4 y 6. Los datos del grupo control están sobrepuestos mediante la línea gris.
La Figura 4 muestra el consumo de calorías. Las barras de color negro representan el promedio de consumo de calorías durante las fases de línea base y libre acceso sin glucosa (1, 3, 5 y 7), los conjuntos de barras bicolores representan el promedio de consumo diario de calorías en que se adicionó glucosa a la bebida (fases 2, 4 y 6). Las calorías consumidas en el alimento y en la bebida se representan con los colores negro y blanco respectivamente. Los datos de los sujetos control se representan por la línea continua sobrepuesta en los datos de los sujetos experimentales. La línea representa de igual forma, el promedio de consumo de calorías durante las fases 1, 3, 5 y 7 y el consumo diario de calorías durante los 5 días de duración de las fases 2, 4 y 6.
Durante las fases 1, 3, 5 y 7, los sujetos experimentales consumieron un promedio de 90 calorías (machos) y 70 calorías (hembras). Los sujetos control mostraron el mismo promedio de consumo durante las fases ya mencionadas y se mantuvo durante los 6 días de duración de las fases 2, 4 y 6. Por su parte, los sujetos experimentales incrementaron su consumo de calorías en estas fases (2, 4 y 6) con respecto a las fases sin glucosa. El sujeto ML1 mostró un incremento promedio de 10 calorías. El sujeto ML2 mostró un incremento promedio de 30 calorías, sin embargo, su consumo de calorías se duplicó en los días 2 y 6 de las fases 4 y 6 respectivamente. Las hembras consumieron entre 15 y 20 calorías más como promedio. El sujeto HL1 también duplicó su consumo el día 5 de la fase 4. En la secuencia de concentraciones alta-media-baja y baja-media-alta en la que recibieron el agua con glucosa se puede observar que a mayor número de calorías en el agua, menor consumo de calorías en el alimento respecto a las otras concentraciones, y a menor número de calorías en el agua, mayor el consumo de alimento respecto a las otras concentraciones.
Discusión
Los datos mostraron que: 1) la curva de crecimiento del peso corporal se mantuvo constante; 2) el consumo de alimento (chow) disminuyó durante las fases de libre acceso con glucosa respecto al consumo registrado durante las fases de libre acceso sin glucosa; 3) el consumo de agua con glucosa fue mayor que el consumo de agua; 4) el consumo de agua con glucosa se incrementó gradualmente en el grupo 1(que recibió la secuencia alta-media-baja) y disminuyó gradualmente en el grupo 2 (que recibió la secuencia baja-media-alta) en cada una de las dos secuencias en las que se adicionaron las tres concentraciones de glucosa en agua; y, 5) los grupos 1 y 2 consumieron más calorías que el grupo control.
El peso corporal de los seis sujetos no presentó modificaciones relevantes en su curva de crecimiento normal, a pesar de que cuatro de los seis sujetos recibieron una mayor cantidad de calorías. Según Staddon (2003) este fenómeno se debe al balance de energía que se mantiene a pesar de la variación en la disponibilidad del alimento o de las calorías, por tal razón no se observan cambios significativos en el peso corporal o en los consumos de alimento en las ratas. Sin embargo, los sujetos experimentales disminuyeron su consumo de alimento cuando tuvieron disponible una fuente de energía alterna en la glucosa. El consumo de alimento decreció durante las fases de libre acceso con glucosa respecto a las fases de libre acceso sin glucosa. Situación opuesta al consumo de bebida, el cual se incrementó durante las fases de libre acceso con glucosa respecto a las fases de libre acceso sin glucosa. Estos resultados sugieren que la variación del contenido energético en el agua modificó los consumos de agua y alimento; esto no representó cambios en el peso corporal.
Esta distribución de los consumos de alimento y agua sugiere que la rata modificó sus consumos mediante un proceso de regulación energética (Collier, Hirsh & Kanareck, 1983; Corwin et al., 1998; Weingarten, 1990; Strubbe & Woods, 2004). Sin embargo, los resultados demostraron que el número de calorías consumidas durante las fases de libre acceso con glucosa fue mayor que el registrado durante las fases de libre acceso sin glucosa. Partiendo de este resultado, sería interesante evaluar el nivel de regulación en el consumo de alimento ante una mayor variedad de concentraciones energéticas. Bolles (1983); Collier, Hirsh y Kanareck (1983); Corwin et al. (1998) y Dimitrou, Rice y Corwin, (2000) afirmaron que las ratas mantienen estable su consumo de calorías a pesar de la disponibilidad de otros alimentos. Sin embargo, nuestros resultados no coinciden con esta postura, probablemente debido al efecto de otros factores como el nivel de actividad de los sujetos. Según Garrow (1986) la cantidad de energía consumida es similar a la cantidad de energía expedida, si la ecuación falla, los organismos desarrollan obesidad, lo que no se observó en ninguno de los sujetos experimentales.
Por otro lado, se observó una relación proporcional entre la presencia de grandes bebidas de agua con glucosa y la concentración utilizada, pues a menor concentración de glucosa, mayor el consumo de bebida, y a mayor concentración de glucosa, menor el consumo de bebida. Mook (1974) señaló que las ratas consumen grandes cantidades de agua endulzada por sus propiedades hedónicas y postingestivas. Realizó algunas manipulaciones utilizando dos soluciones endulzadas y dos dietas líquidas endulzadas para medir la preferencia del animal. No encontró evidencias precisas de las preferencias de soluciones más concentradas en ratas. Sin embargo, su procedimiento consistía en ofrecer al animal las soluciones por un tiempo breve y además, utilizó sacarina para endulzar las soluciones. La sacarina no contiene calorías, por lo tanto, no produce consecuencias postingestivas. Para Mook (1974) la palatabilidad de soluciones o comidas dulces no garantiza que los animales muestren preferencias sólidas. Señaló que las ratas no consumen grandes cantidades de agua con sacarina y no modifican su ingesta diaria de alimento. Cuestionó el papel del sabor dulce en las preferencias alimentarias y afirmó que las ratas no consumen grandes cantidades de agua con sacarina por ausencia de efectos calóricos.
Los resultados de nuestro experimento demostraron que la variación del contenido calórico de los alimentos modificó el patrón alimentario de la rata sin tener efecto sobre el peso corporal, a pesar de que aumentó el número de calorías consumidas. Esta evidencia sugiere la necesidad de realizar nuevos estudios que evalúen el proceso de regulación conductual en situaciones de cambios energéticos alimentarios. Finalmente, es necesario en futuras investigaciones analizar de forma separada el papel del sabor y del contenido energético de los alimentos.
Referencias
Beatty. D. (1978). Brief communication: Operant responding in rats with dietary obesity. Physiology and Behavior, 21, 671-672. [ Links ]
Bolles, R. C. (1983). Teoría de la motivación. México: Trillas. [ Links ]
Capaldi, E. D., Campell, D. H., Sheffer, J. D. & Bradford, J. P. (1987). Conditioned flavor preferences based on delayed caloric consequences. Journal of Experimental Psychology: Animal Behavior Processes, 13, 150-155. [ Links ]
Collier, G., Hirsh, E. & Kanareck, R. (1983). La operante vista de Nuevo. En W. K. Honig, y J. E. R. Staddon (eds.). Manual de conducta operante. México: Trillas. [ Links ]
Corwin, R. L., Wojnicki, F. H. E., Fischer, J. O., Dimitrou, S. G., Rice, H. B. & Young, M. A. (1998). Limited access to a dietary fat option affects ingestive behavior but not body composition in male rats. Physiology and Behavior, 65, 545-553. [ Links ]
Davis, J. D. & Levine, M. W. (1977). A model for the control of ingestion. Psychological Review, 84, 379-412. [ Links ]
Dimitrou, S. G., Rice, H. B. & Corwin, J. (2000). Effects of limited access to a fat option on food intake and body composition in female rats. International Journal of Eating Disorders. John Wiley & son´s Inc. [ Links ]
Galef, B. G. (1996). Social influences on food preferences and feeding behavior of vertebrates. En E. D Capaldi (ed). Why we eat what we eat (pp. 207-231). USA: American Psychological Association. [ Links ]
Garrow, J. S. (1986). Physiological aspects of obesity. En K. D. Brownell y J. P. Forext (Eds.). Handbook of eating disorders (pp. 45-62). New York: Basic Books, Publishers. [ Links ]
Keesey, R. E. (1986). A set-point theory of obesity En K. D. Brownell y J. P. Foreyt (Eds.), Handbook of eating disorders (pp. 63-87). New York: Basic Books, Publishers. [ Links ]
Lane, M. A., Ingram, D. K. & Roth, G. S (1999). Nutritional modulation on aging in nonhuman primates. The Journal of Nutrition, Health y Aging, 3, 69-78. [ Links ]
Mayer, J. (1955). Regulation of energy intake and body weight. The glucostatic theory and the lipostatic hypothesis. Annals of the New York Academy of sciences, 63, 15-43. [ Links ]
Mook, D. G. (1974). Saccharin preference in the rat: some unpalatable findings. Psychological Review, 81, 475-490. [ Links ]
Staddon, J. E. R. (2003). Adaptive behavior and learning. (Internet ed.). Nueva York: Cambridge University Press. [ Links ]
Staddon, J. E. R. & Zanutto, B. S. (1998). In praise of parsimony. En D. L. Wynne y J. E. R. Staddon (eds.). Models of action. USA: Lawrence Erlbaum Associates Publishers. [ Links ]
Strubbe, J. H. & Woods, S. C. (2004). The timming of meals. Psychological Review, 111, 148-121. [ Links ]
Subcommittee on Laboratory Animal Nutrition, Committee of Animal Nutrition, Board on Agriculture, National Research Council (1995). Nutrient Requirements of Laboratory. En línea: National Academies Press. [ Links ]
Teiltelbaum, P. & Campell, B. A. (1958). Ingestion patterns in hyperphagic and normal rats. Journal of Comparative and Physiological Psychology, 51, 135-141. [ Links ]
Treit, D., Spetch, M. L. & Deutsch, J. A. (1982). Variety in the flavor of food enhances eating in the rat: a controlled demonstration. Physiology and Behavior, 30, 207-211. [ Links ]
Weingarten, H. P. (1990). Learning, homeostasis and control of feeding behavior. En E. D. Capaldi, y T. L. Powley (eds.). Taste, experience and feeding: development and learning. USA: American Psychological Association. [ Links ]
Woods, S. C., & Seeley, R. J. (2003). Hunger and energy homeostasis. En R. Gallistel y H. Pashler (eds.). Steven´s handbook of experimental psychology (pp. 633-668). Nueva York: John Wiley & Sons. [ Links ]
Young, P. T. & Shuford, E. H. (1955). Quantitative control of motivation through sucrose solutions of different concentrations. Journal of Comparative and Physiological Psychology, 48, 114-118. [ Links ]

