










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkUniversitas Psychologica
versão impressa ISSN 1657-9267
Univ. Psychol. v.6 n.2 Bogotá maio/ago. 2007
FACILITACIÓN DE RELACIONES BIDIRECCIONALES EN PALOMAS USANDO UNA TAREA DE TRANSFERENCIA TRAS ENTRENAMIENTO EN DISCRIMINACIÓN DE LA PROPIA CONDUCTA1
FACILITATION OF BI-DIRECTIONAL RELATIONSHIPS IN PIGEONS USING A TRANSFER TASK AFTER TRAINING IN DISCRIMINATION OF OWN BEHAVIOR
ANDRÉS GARCÍA, SANTIAGO BENJUMEA
UNIVERSIDAD NACIONAL DE EDUCACIÓN A DISTANCIA, ESPAÑA, Departamento de Psicología Básica I, Facultad de Psicología, Universidad Nacional de Educación a Distancia. C / Juan del Rosal 10, Ciudad Universitaria, 28040-Madrid, España. Número de teléfono: 913987971. Fax: 913987972. E-mail: agargar@psi.uned.es
UNIVERSIDAD DE SEVILLA, ESPAÑA
Recibido: julio 3 de 2006 Revisado: febrero 9 de 2007 Aceptado: abril 18 de 2007
ABSTRACT
An experiment was carried out in which 10 pigeons were trained in a task of conditional discrimination of their own behavior. The birds learned that when the sample had consisted on responding to the left (right) with two white keys, they should choose the red (green) comparison to be reinforced. Once the animals had learned this task, they were passed to a test phase. In short, 5 pigeons were trained in a positive transfer, where the sample was now the red (green) key they had to choose the left (right) comparison to obtain the reinforcer. On the contrary, the other 5 pigeons received a training of negative transfer, where the task consisted in that with a red (green) sample they should choose the right (left) comparison to be reinforced. The obtained data indicate that the performance of those subjects of the group in which there was coherence among the phases went superior to that of those subjects of the group where there was not this coherence. The results are interpreted in connection with the importance that the discrimination of the own behavior has in the derivation of bidirectional relationships.
Key words: stimulus control, symmetry, conditional discrimination, differential response, pigeons.
RESUMEN
Se llevó a cabo un experimento en el que diez palomas fueron entrenadas en una tarea de discriminación condicional de su propia conducta. Los sujetos aprendían que cuando la muestra había consistido en responder a la izquierda (derecha) ante dos teclas blancas, debían elegir la comparación roja (verde) para ser reforzados. Una vez que los animales habían aprendido esta tarea, se les pasaba a una fase de prueba. En concreto, cinco palomas fueron entrenadas en una transferencia positiva, ya que si la muestra era ahora la tecla roja (verde) tenían que elegir la comparación izquierda (derecha) para llegar al reforzador. Por el contrario, las otras cinco palomas recibían un entrenamiento de transferencia negativa, ya que la tarea consistía en que ante una muestra roja (verde) debían elegir la comparación derecha (izquierda) para ser reforzados. Los datos obtenidos indican que la ejecución de los sujetos del grupo en el que había coherencia entre las fases fue superior a la de los sujetos del grupo en el que no había dicha coherencia. Los resultados son interpretados en relación con la importancia que la discriminación de la propia conducta tiene en la derivación de relaciones bidireccionales.
Palabras clave: control de estímulos, simetría, discriminación condicional, respuesta diferencial, palomas.
La situación típica en la que se obtiene el fenómeno de las clases de equivalencia de estímulo es el entrenamiento de los sujetos en un procedimiento de discriminación condicional estándar (Sidman & Tailby, 1982). Durante este entrenamiento, si se establece una asociación consistente entre tres estímulos diferentes, las relaciones aRa, bRb, cRc (reflexividad), bRa, cRb (simetría), aRc (transitividad) y cRa (equivalencia) ocurren en ausencia de entrenamiento específico. La obtención de clases de equivalencia entre estímulos se ha observado en humanos (Dugdale & Lowe, 1990; Hayes & Hayes, 1989; Sidman, 1994), pero todavía no en animales (Dugdale & Lowe, 2000; Zentall, 1996).
De las diferentes relaciones derivadas incluidas en las clases de equivalencia, la simetría ha resultado ser crítica con respecto a las diferencias entre humanos y otras especies. Aunque hay alguna evidencia (Hearst, 1989; Honey & Hall, 1989) del desarrollo de relaciones simétricas siguiendo el entrenamiento de condicionamiento pavloviano (asociaciones hacia atrás), los resultados negativos han sido la regla en la prueba de simetría en animales (Gray, 1966; Hogan & Zentall, 1977; Lionello- DeNolf & Urcuioli, 2002; Richards, 1988; Rodewald, 1974; Sidman et al., 1982). Un informe positivo (McIntire, Cleary & Thompson, 1987) podría depender de las cadenas directamente condicionadas de conducta mediada (Dube, McIlvane, Callahan & Stoddard, 1993; Hayes, 1989; Saunders, 1989). Incluso en estudios donde se han encontrado las equivalencias derivadas (por ejemplo, en leones marinos, Kastak, Schusterman & Kastak, 2001; Schusterman & Kastak, 1993. En chimpancés, Yamamoto & Asano, 1995. Y en palomas, Meehan, 1999) los animales tuvieron dificultades iniciales para demostrar relaciones simétricas no entrenadas (por ejemplo, dado el entrenamiento AB, inicialmente, no se encontraron las relaciones BA derivadas) (véase Urcuioli & DeMarse, 1997).
El origen de la derivación de relaciones no entrenadas entre los estímulos en las clases de equivalencia ha constituido el centro de atención de la investigación en la materia. Dos ideas se han destacado en la comunidad científica con respecto a este tema.
Por una parte, Sidman (1990, 1994, 2000) sugirió que la equivalencia de estímulos podría ser un “primitivo”, una función básica no derivada de otra más simple, y concluyó que la equivalencia de estímulos es una propiedad emergente de los procedimientos de entrenamiento de discriminación condicional. Según esta hipótesis del “primitivo”, el fallo en obtener equivalencia en animales habría que buscarlo, en último término, en la filogenia.
Por otro lado, como Dube y colaboradores han señalado (Dube et al., 1993), los resultados positivos con los procedimientos de equivalencia estándar involucran requisitos conductuales previos, que los humanos, pero no los animales de laboratorio, probablemente habrán adquirido a través de su experiencia pre-experimental. Algunos autores han postulado que el lenguaje podría ser este pre-requisito. Los humanos formarían clases de estímulos equivalentes, no porque tengan una habilidad innata específica (“primitivo”), sino porque cuando los estímulos aparecen los nombran, los etiquetan (hipótesis del Naming. Ver, por ejemplo, Dugdale & Lowe, 1990; Hayes & Hayes, 1989).
Catania, Matthews y Shimoff (1990) han sugerido que el carácter bidireccional de la conducta verbal y de la equivalencia de estímulos podría ser una manifestación de la misma competencia conductual, a saber, la habilidad de establecer discriminaciones condicionales basadas en la propia conducta.
Sidman (2000) ha sugerido que las diferencias encontradas entre animales y humanos podrían ser debidas a variables críticas de procedimiento. Aun aceptando que la equivalencia sea un primitivo que se derive directamente de la contingencia de reforzamiento, necesitamos incluir en las clases de equivalencia, no sólo las muestras y los estímulos de comparación, sino también las respuestas y los reforzadores. Visto de esta forma, la hipótesis del Naming podría ser incluida sin controversia dentro de este marco: el factor crucial podría ser que los humanos utilizan diferentes respuestas (nombres, etiquetas), mientras que en los procedimientos habitualmente usados con animales se tendría la misma respuesta para los diferentes estímulos.
Existen evidencias de que cuando se utilizan respuestas diferentes en discriminaciones condicionales con animales se obtienen indicios de equivalencia. Así, en el trabajo de McIntire et al. (1987) los sujetos fueron entrenados para llevar a cabo conductas diferenciales a la muestra y la comparación correcta en cada caso. Otro estudio que presentó resultados positivos, en relación con las relaciones derivadas, fue el de Zentall, Sherburner y Steirn (1992), en el que se usaron como muestras estímulos biológicamente relevantes (estímulos incondicionados). Con este procedimiento lo que podría pasar es que las palomas realmente discriminaran (y escogieran las comparaciones correctas en función de) las respuestas consumatorias (intensas y, por consiguiente, más discriminables) que se llevaban a cabo ante cada estímulo de muestra. En la misma línea, uno de los factores más relevantes en el trabajo de Meehan (1999) fue el uso de reforzadores diferenciales para cada una de las clases que intentó formar en su experimento. Como él mismo mencionó, este uso de reforzadores diferentes generó modelos conductuales distintos a cada muestra. Como ya hemos comentado, esos modelos tenían muchas probabilidades de ser el evento discriminado por las palomas de ese estudio.
En todos los trabajos anteriores se han usado diferentes estímulos de muestra junto con respuestas distintas. Existe, sin embargo, la posibilidad más simple de establecer discriminaciones condicionales basadas exclusivamente en respuestas diferentes como muestras sin acompañamiento de estímulos exteroceptivos diferentes. Aunque existen antecedentes experimentales de este tipo de discriminaciones condicionales (Baninger, Kendall & Vanderwolf, 1974; Lionello-DeNolf & Urcuioli, 2003; Reynolds & Catania, 1962; Shimp, 1982, 1983, 1984), en ninguno de ellos se puso a prueba la obtención de relaciones simétricas.
El objetivo del presente trabajo fue, pues, comprobar la obtención de simetría en una situación de discriminación condicional con la propia conducta del sujeto como estímulo (propioceptivo) de muestra, usando palomas como sujetos experimentales.
Método
Sujetos
Se usaron diez palomas (Columba Livia) experimentalmente ingenuas, mantenidas aproximadamente al 80% de sus pesos con alimento libre. Los cuartos en los que fueron alojadas se mantuvieron en un ciclo 12:12 luzoscuridad. El agua estuvo continuamente disponible.
Aparatos
Se usaron cinco cámaras experimentales estándar con tres teclas. Cada cámara tenía 27.5 cm de alto, 24 cm de profundidad y 32 cm de ancho. Una pared de la cámara contenía dos teclas de estímulo/respuesta (luces blancas, rojas, o verdes de 6 W), una luz general, y una apertura (5 cm por 6 cm) para proporcionar el acceso al grano mixto de la paloma. Cada tecla estaba situada aproximadamente a 5 cm de la pared más cercana, y a 13 cm de la otra tecla (distancias entre los centros). Se usó un ordenador para controlar y registrar los eventos experimentales.
Procedimiento
El experimento se dividió en tres fases: preentrenamiento, entrenamiento en discriminación condicional y prueba de simetría.
Fase de preentrenamiento. Después del entrenamiento en el comedero, los sujetos realizaron tres sesiones de automoldeamiento (estímulo condicionado (EC) de 8’’ e intervalo entre ensayos (IEE) de 52’’) con razón fija 1 (RF1) para que las palomas adquiriesen el picoteo en todas las combinaciones de colores (blanco, rojo y verde) y posiciones (izquierda y derecha) de las teclas. Además, los animales fueron expuestos a un programa de RF10 reforzado con comida para aumentar el número de respuestas a las teclas en todos los colores y posiciones. Después, los sujetos se asignaron al azar a dos condiciones de entrenamiento de cinco pájaros cada uno.
Fase de entrenamiento de discriminación condicional. Cada ensayo de entrenamiento consistió en dos componentes consecutivos: un componente de muestra seguido por un componente de comparación (Figura 1). El componente de muestra comenzaba con la iluminación de dos teclas con luz blanca. En cada ensayo una tecla fue asignada como “correcta” y la otra como “incorrecta”, con una probabilidad de 0.5. Cada vez que una paloma picaba en la tecla incorrecta el contador de la tecla correcta se reiniciaba. Así, cinco picotazos consecutivos en la tecla correcta eran la única manera de acabar este componente, que era seguido por el de comparación, después de un intervalo de 2’’ en el que ambas teclas estaban apagadas. El componente de comparación consistía en las dos teclas iluminadas, una de verde y una de rojo, en posición (izquierda-derecha) aleatorizada.
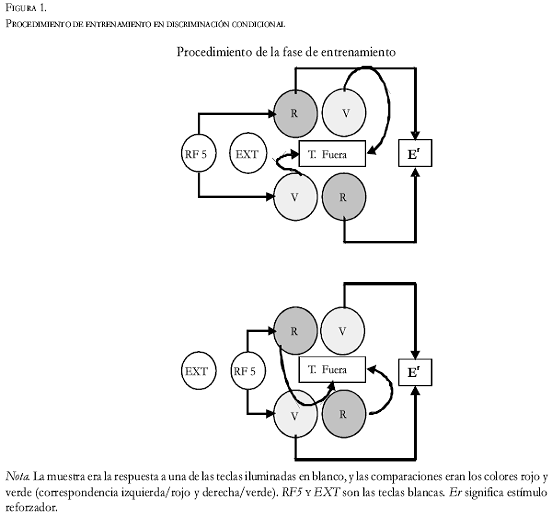
La mitad de los sujetos fueron reforzados por picar la tecla roja después de picar la tecla blanca izquierda, y por picar la tecla verde después de picar la blanca derecha (correspondencia izquierda-rojo; derecha-verde. Ver la Figura 1). La otra mitad de los sujetos eran reforzados por picar la tecla verde después de picar la tecla blanca izquierda, y por picar la tecla roja después de picar la tecla blanca derecha (correspondencia izquierdaverde; derecha- rojo).
Cada picotazo (RF1) al color correcto se reforzó con comida. Picar la tecla del color incorrecto fue seguido por un intervalo de 10’’ de tiempo fuera (con la luz general apagada) y otra presentación del mismo componente de comparación. De esta forma, garantizamos que cada color recibió la misma cantidad de reforzamiento. Cada sesión concluyó cuando se entregaron 40 reforzadores. Este entrenamiento continuó hasta que el sujeto respondía correctamente, al menos al 90% de los ensayos en tres sesiones consecutivas (media se sesiones: 48.7, rango: 35-65).
Fase de transferencia (prueba de simetría). Conforme los sujetos respondían adecuadamente en la tarea de discriminación de su propia conducta, pasaban a la tercera fase. En ésta, fueron repartidos en dos grupos: uno de transferencia positiva (T+) y otro de transferencia negativa (T-). Estos grupos (de cinco sujetos cada uno) se igualaron tanto en la velocidad de adquisición en la primera fase, como en el estado final de discriminación (las cinco palomas del grupo de transferencia positiva obtuvieron como promedio un 95% de aciertos, durante las últimas cinco sesiones de la primera fase, mientras que los sujetos del grupo de transferencia negativa obtuvieron un 96% de aciertos).
En esta fase de prueba, los eventos que actuaron anteriormente como muestra (responder izquierda o derecha) y comparación (rojo o verde) intercambian sus papeles (ver Figura 2). Tras el IEE inicial, los sujetos se encuentran con una tecla roja y otra verde (su posición fue aleatorizada). Tras responder a una de ellas (seleccionada al azar para cada ensayo) aparecían las dos teclas iluminadas de blanco. Todas las demás variables fueron idénticas a las de la fase de entrenamiento.
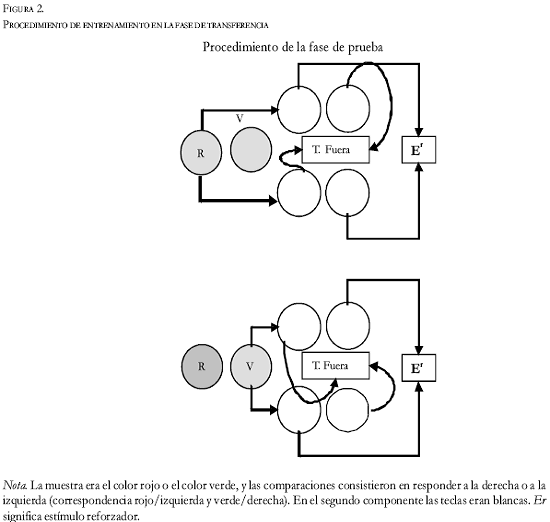
Grupo de transferencia positiva: Para los sujetos S1 y S4 funcionó la siguiente lógica: si para pasar de una situación (R-V o V-R) a la otra (B-B) el sujeto debió responder cinco veces consecutivas al rojo, tendrá ahora que elegir la tecla izquierda para ser reforzado. Si, al contrario, pasó gracias a cinco respuestas al verde, tendrá que elegir la tecla derecha en ese ensayo para conseguir la comida. Para los sujetos S13, S24 y S31 la contingencia era ésta: si para pasar de una situación (R-V o V-R) a la otra (B-B) tuvieron que responder cinco veces consecutivas al rojo, ahora deberán elegir la tecla derecha para ser reforzados. Si pasaron gracias a cinco picotazos al verde, la opción reforzada será picar en la tecla izquierda.
Vemos, pues, que entre ambas fases hay “coherencia”, ya que a los sujetos a los que se les entrenó en la primera fase en las secuencias “tras responder izquierda - elegir rojo” y “tras responder derecha - elegir verde”, se les enseña en la segunda las secuencias “tras responder rojo - elegir izquierda” y “responder verde - responder derecha”. También, a los sujetos a los que en la primera fase se les reforzó si seguían estas secuencias: “tras responder izquierda - elegir verde” y “tras responder derecha- elegir rojo”, se les refuerza en la segunda fase responder así: “tras responder verde - elegir izquierda” y “tras responder rojo - elegir derecha”. Llamamos, pues, a este grupo de TRANSFERENCIA POSITIVA.
Grupo de transferencia negativa: Este grupo fue igual que el anterior, con la excepción de que en éste no hay “coherencia” entre las fases de entrenamiento y prueba. Así, a los sujetos S14 y S30, que en la primera fase fueron entrenados en las secuencias “tras responder derecha - elegir rojo” y “tras responder izquierda - elegir verde”, se les pedía en la segunda fase que aprendiesen las secuencias simétricas contrarias: “tras responder rojo - elegir izquierda” y “tras responder verde - elegir derecha”. Del mismo modo, a los sujetos S2, S3 y S5, que en la primera fase fueron entrenados en las cadenas conductuales “tras responder izquierda - elegir rojo” y “tras responder derecha - elegir verde”, se les entrena ahora para que aprendan que para ser reforzados deben seguir estas secuencias: “tras responder rojo - elegir derecha” y “tras responder verde - elegir izquierda”. Entonces, a este grupo lo llamamos de TRANSFERENCIA NEGATIVA.
En las dos fases del experimento se utilizó el mismo procedimiento de corrección.
Resultados
Fase de discriminación condicional
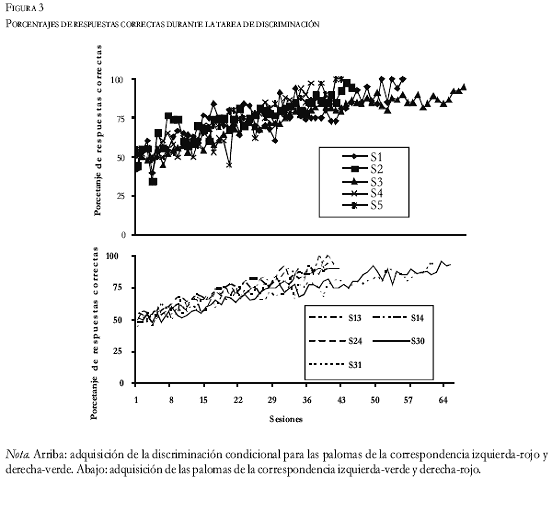
Todas las palomas consiguieron un índice muy alto de aciertos (Figura 3) en la tarea de discriminación. Durante las primeras diez sesiones, su conducta estaba en el nivel de azar. Cuando llevaban 25 sesiones mostraron índices de acierto cercanos al 70%. Después, mejoraron más lentamente: subieron del 85% de aciertos, en la sesión 40, al 95% al final del entrenamiento.
Prueba de simetría
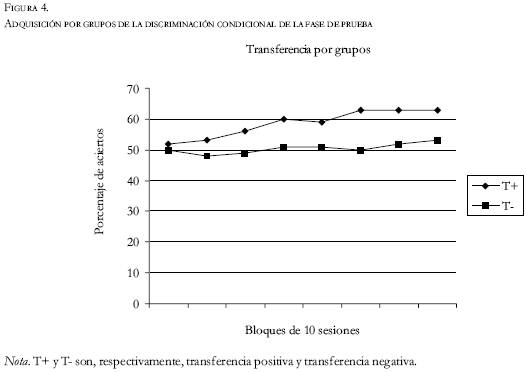
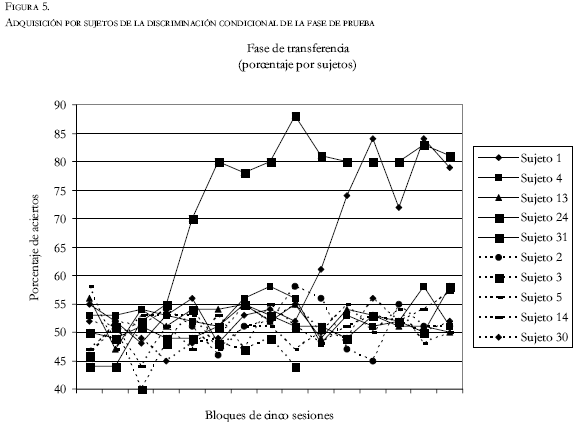
Como se puede ver en las figuras 4 y 5, durante las veinte primeras sesiones de la tercera fase (de transferencia), todos los sujetos de ambos grupos se movieron entre el 45% y el 55% de aciertos en la tarea. Sólo en cuatro de estas veinte sesiones los sujetos de transferencia negativa obtuvieron mejores resultados como grupo que los sujetos de transferencia positiva. Es decir, aunque con escasas diferencias, en el 80% de la primera parte de la prueba el grupo de transferencia positiva se situó por encima del grupo de transferencia negativa. A partir de este momento, las diferencias no hicieron más que aumentar, y siempre se situó como promedio el grupo de transferencia positiva con un índice de discriminación más alto que el de transferencia negativa. Además, la magnitud de esta diferencia se ampliaba conforme avanzaban las sesiones. Así, la diferencia era de siete puntos en la sesión 30, nueve en la sesión 40, y 14 en la sesión 60. Por lo que respecta al análisis estadístico, no se encontraron diferencias significativas (T = 0.5; P = 0.62) entre el promedio del índice de aciertos de los sujetos de transferencia positiva y de transferencia negativa durante las primeras 15 sesiones. Por el contrario, sí se encuentran diferencias (T = 7.11; P = 0.000) al comparar las últimas 15 sesiones. Del mismo modo, las últimas 15 sesiones del grupo de transferencia positiva se diferenciaron estadísticamente (T = 9.04; P = 0.000) de las 15 primeras sesiones de dicho grupo. Esta misma comparación no fue significativa (T = 1.54; P=0.144) para los sujetos del grupo de transferencia negativa.
Si en lugar de llevar a cabo un análisis por grupos, lo hacemos por sujetos (Figura 5), nos daremos cuenta de que las diferencias observadas están motivadas principalmente por dos sujetos pertenecientes al grupo de transferencia positiva. Entre las sesiones 20 y 25, el sujeto 24 comenzó a subir su nivel de acierto hasta un 70%. Desde este momento hasta el final del entrenamiento (75 sesiones), nunca bajó del 80% de discriminación. Cuando llevábamos aproximadamente 50 sesiones de entrenamiento en esta tercera fase, el sujeto 1 comenzó a subir su nivel de aciertos. Diez sesiones más tarde se situó en el 80% de aciertos y se mantuvo a ese nivel hasta el final del estudio.
Discusión
Los principales hallazgos del presente trabajo han sido los siguientes: a) las palomas demostraron la habilidad para discriminar y etiquetar su propia conducta; b) se ha facilitado el aprendizaje de la inversión en una tarea de discriminación de su propia conducta con palomas como sujetos.
Esta facilitación, no obstante, es relativa. Aunque ningún sujeto del grupo de transferencia negativa la ha mostrado, en el de transferencia positiva sólo dos sujetos han obtenido resultados positivos. Posiblemente la estrategia de evaluación mediante una fase de entrenamiento y otra de prueba no sea la más adecuada, y puede ser sustituida por una estrategia de evaluación que incluya ensayos de prueba intercalados con los de entrenamiento (García, 2000).
Por tanto, aunque con las limitaciones comentadas, se han obtenido indicios de que la discriminación de la propia conducta facilita la simetría con palomas como sujetos. Cuando la relación entre la muestra y la comparación era del tipo respuesta-estímulo en lugar del habitual estímulo-estímulo, esta relación se hizo bidireccional. El argumento que M. Sidman utilizó para defender que la equivalencia era una función básica no derivable de otra, era la importancia de agregar en una discriminación condicional la relación E-E a la relación E-R existente en una discriminación simple. Al trabajar con humanos difícilmente se puede descartar la hipótesis del Naming para explicar la emergencia de nuevas relaciones. Este Naming (conducta oral diferencial a la muestra) estaría convirtiendo la discriminación del evento externo (E-E) en una discriminación de la propia conducta (R-E) del sujeto. Por otro lado, la evidencia de simetría con nohumanos usando discriminaciones del tipo E-E es escasa. Al emplear una discriminación en la que la conducta del sujeto es el evento a discriminar, hemos aumentado esa evidencia.
Cuando decimos que una relación de control es simétrica, queremos expresar que el evento que controla en un momento y una situación determinada es después el evento controlado. La conducta que controla una primera elección es luego controlada (y evocada) por la etiqueta que se le asignó. Podemos decir que somos conscientes de nuestros actos cuando existe una completa (y bidireccional) relación entre nuestra conducta y los símbolos que le son asociados.
Parece ser que hay una relación estrecha entre el uso de discriminaciones con procedimientos que generan conductas diferenciales en los sujetos y la aparición de estas relaciones bidireccionales. Por una parte, tenemos los estudios con seres humanos (sobre todo con niños) en los que la presencia del Naming (conducta verbal diferencial) posee una gran importancia para la emergencia de simetría. Se ha comentado previamente la fuerte relación empírica entre el lenguaje y las relaciones emergentes de estímulos. De hecho, la investigación conductual en las relaciones derivadas ha estado desde el principio estrechamente unida al entrenamiento y el uso del lenguaje (Sidman, 1971). Existen también otros datos (Horne & Lowe, 1996) que indican la no-emergencia de relaciones en los niños pequeños.
Por otro lado, en algunos de los trabajos llevados a cabo con animales no-humanos que han mostrado buenos resultados en este tema, la discriminación de la propia conducta podría estar jugando una función crucial (García, 2000). En dichos trabajos, lo que el sujeto podría estar aprendiendo era a relacionar una respuesta emitida por él con una etiqueta. Y todo parece indicar que ahora la etiqueta podría evocar la aparición de la conducta correspondiente. Al contrario que los eventos no verbales, (apretar palancas y picar teclas no son intercambiables con las luces y los tonos), los eventos verbales funcionan como estímulos y como respuestas. Y esa bifuncionalidad parece vital para la aparición de la bidireccionalidad de las relaciones. Este tipo de relación simétrica que tiene lugar entre una conducta verbal y la conducta (gobernada-por-la-regla) hace simbólica la conducta verbal.
Como Skinner afirma (1953), parece que las respuestas discriminativas a la propia conducta y las variables de las que son la función, son producto exclusivo del ambiente social. El hecho de que un sujeto sea o no consciente de sí mismo e introspectivo depende de la intensidad con la que el grupo haya insistido en obtener respuestas a preguntas como “¿qué estás haciendo?” o “¿por qué lo hiciste?”.
Lo que realmente hemos hecho en nuestro estudio ha sido situar a una paloma en una situación experimental que coloca la propia conducta de uno en el lugar del estímulo condicional, y también pone una comparación correcta y una incorrecta. Hemos situado a las palomas en un contexto de aprendizaje y evaluación, en el que sólo los miembros de la especie humana (dentro de los dominios ontogénicos y culturales de su desarrollo) se han localizado tradicionalmente. Y los resultados han sido, en un cierto sentido, convergentes: la apariencia de relaciones simétricas a las directamente entrenadas. Cuando Sidman (1990) consideró el desarrollo de clases de la equivalencia como un primitivo no derivable o función básica, y también exclusivo de la especie humana, estaba localizando este fenómeno dentro del dominio de desarrollo filogenético de la especie humana. Al trabajar con individuos de otra especie dentro de un episodio genuino de los dominios ontogénicos y culturales de los humanos, hemos dado otro paso en la comprensión de la génesis de esta capacidad.
1 Esta investigación ha sido financiada en parte por los proyectos del Ministerio de Ciencia y Tecnología (BSO2002-04322 a Andrés García y BSO2003-02182 a Santiago Benjumea) y del proyecto de la Junta de Andalucía a Santiago Benjumea (HUM-633).
Referencias
Baninger, R. J., Kendall, S. B. & Vanderwolf, C. H. (1974). The Ability of Rats to Discriminate Their Own Behaviours. Canadian Journal of Psychology, 28 (1), 79-91. [ Links ]
Catania, A .C., Matthews, A. A. & Shimoff, E. (1990). Properties of Rule-governed Behavior and Their Implications. En D. E. Blackman & H. Lejeune (Eds.), Behaviour Analysis in Theory and Practice. Contributions and Controversies (pp. 215-230). Hove: Erlbaum. [ Links ]
Dube, W. V., McIlvane, W. J., Callahan, T. D. & Stoddard, L. T. (1993). The Search for Stimulus Equivalence in Nonverbal Organisms. The Psychological Record, 43, 761-778. [ Links ]
Dugdale, N. A. & Lowe, C. F. (1990). Naming and Stimulus Equivalence. En D. E. Blackman & H. Lejeune (Eds.), Behavior Analysis in Theory and Practice. Contributions and Controversies (pp. 115- 138). Hove: Erlbaum. [ Links ]
Dugdale, N. A. & Lowe, C. F. (2000). Testing for Symmetry in the Conditional Discriminations of Language-trained Chimpanzees. Journal of the Experimental Analylsis of Behavior, 73, 5-22. [ Links ]
García, A. (2000). Discriminación de la propia conducta y emergencia de simetría en palomas. Disertación doctoral no publicada, Universidad de Sevilla, Sevilla. [ Links ]
Gray, L. (1966). Backward Association in Pigeons. Psychonomic Science, 4, 333-334. [ Links ]
Hayes, S.C. (1989). Rule-Governed Behavior: Cognition, Contingencies and Instructional Control. New York: Plenum press. [ Links ]
Hayes, S. C. & Hayes, L. J. (1989). The Verbal Action of the Listener as a Basis for Rule-governance. En S. C. Hayes (Ed.), Rule-Governed Behavior: Cognition, Contingencies and Instructional Control (pp. 153- 188). New York: Plenum Press. [ Links ]
Hearst, E (1989). Backward Associations: Differential Learning about Stimuli that Follow the Presence versus the Absence of Food in Pigeons. Animal Learning and Behavior, 17, 280-290. [ Links ]
Hogan, D. E. & Zentall, T. R. (1977). Backward Associations in the Pigeon. American Journal of Psychology, 90, 3-15. [ Links ]
Honey, R. C. & Hall, G. (1989). The Acquired Equivalence and Distinctiveness of Cues. Journal of Experimental Psychology: Animal Behavior Processes, 15, 338-346. [ Links ]
Horne, P. J. & Lowe, C. F. (1996). On the Origins of Naming and Other Symbolic Behavior. Journal of the Experimental Analysis of Behavior, 65, 119-137. [ Links ]
Kastak, C. R., Schusterman, R. J. & Kastak, D. (2001). Equivalence Classification by California Sea Lions Using Class-specific Reinforcers. Journal of the Experimental Analysis of Behavior, 76, 131-158. [ Links ]
Lionello-DeNolf, K. L. & Urcuioli, P. J. (2002). Stimulus Control Topographies and Tests of Symmetry in Pigeons. Journal of the Experimental Analysis of Behavior, 78, 467-495. [ Links ]
Lionello-DeNolf, K. L. & Urcuioli, P. J. (2003). A Procedure for Generating Differential “Sample” Responding without Different Exteroceptive Stimuli. Journal of the Experimental Analysis of Behavior, 79, 21-35. [ Links ]
McIntire, K. D., Cleary, J. & Thompson, T. (1987). Conditional Relations by Monkeys: Reflexivity, Symmetry and Transitivity. Journal of the Experimental Analysis of Behavior, 47, 279-285. [ Links ]
Meehan, E.F. (1999). Class-consistent Differential Reinforcement and Stimulus Class Formation in Pigeons. Journal of the Experimental Analysis of Behavior, 72, 97-115. [ Links ]
Reynolds, G. S. & Catania, A.C. (1962). Temporal Discrimination in Pigeons. Science, 135, 314-315. [ Links ]
Richards, R.W. (1988). A Question of Bidirectional Associations in Pigeons´s Learning of Conditional Discrimination Task. Bulletin of the Psychonomic Society, 26, 577-579. [ Links ]
Rodewald, H. K. (1974). Symbolic Matching to Sample by Pigeons. Psychological Reports, 34, 987-990. [ Links ]
Saunders, K.J. (1989). Naming in Conditional Discrimination and Stimulus Equivalence. Journal of the Experimental Analysis of Behavior, 51, 379-384. [ Links ]
Schusterman, R. J. & Kastak, D. (1993). A California Sea Lion (Zalopbus Californianuus) is Capable of Forming Equivalence Relations. Psychological Record, 43, 823-839. [ Links ]
Shimp, C. P. (1982). On Metaknowledge in the Pigeon: An Organism´s Knowledge about its Own Behavior. Animal Learning and Behavior, 10 (3), 358-364. [ Links ]
Shimp, C. P. (1983). The Local Organization of Behavior: Disociations between a Pigeon´s Behavior and Self-reports of that Behavior. Journal of the Experimental Analysis of Behavior, 39, 61-68. [ Links ]
Shimp, C. P. (1984). Self-reports by Rats of the Temporal Patterning of Their Behavior: A Dissociation between Tacit Knowledge and Knowledge. En H. L. Roitblat, T. G. Bever & H. S. Terrace (Eds.), Animal Cognition (pp. 215-229). Erlbaum: NJ. [ Links ]
Sidman, M. (1971). Reading and Auditory-visual Equivalences. Journal of Speech and Hearing Research, 14, 5-13. [ Links ]
Sidman, M. (1990). Equivalence Relations: Where do They Come From? En D. E. Blackman & H. Lejeune (Eds.), Behaviour Analysis in Theory and Practice. Contributions and controversies (pp. 93-114). Hove: Erlbaum. [ Links ]
Sidman, M. (1994). Equivalence Relations and Behavior: A Research Story. Boston: Authors Cooperative. [ Links ]
Sidman, M. (2000). Equivalence Relations and the Reinforcement Contingency. Journal of the Experimental Analysis of Behavior, 74, 127-146. [ Links ]
Sidman, M., Rauzin, R., Lazar, R., Cunninghan, S., Tailby, W. & Carrigan, P. (1982). A Search for Symmetry in the Conditional Discriminations of Rhesus Monkeys, Baboons and Children. Journal of the Experimental Analysis of Behavior, 37, 23-44. [ Links ]
Sidman, M. & Tailby, W. (1982). Conditional Discrimination vs. Matching to Sample. An Expansion of the Testing Paradigm. Journal of the Experimental Analysis of Behavior, 37, 5-22. [ Links ]
Skinner, B.F. (1953). Science and Human Behavior. Nueva York: MacMillan. [ Links ]
Urcuioli, P. J. & DeMarse, T. B. (1997). Further Tests of Response-outcome Associations in Differential Outcome Matching-to-sample. Journal of Experimental Psychology: Animal Behavior Processes, 23, 171-182. [ Links ]
Yamamoto, J. & Asano, T. (1995). Stimulus Equivalence in a Chimpanzee (Pan Troglodytes). Psychological Record, 45, 3-21. [ Links ]
Zentall, T. R. (1996). An Analysis of Stimulus Class Formation in Animals. En T. R. Zentall & P. M. Smeets (Eds.), Stimulus Class Formation in Humans and Animals (pp.15-34). Amsterdam: North Holland. [ Links ]
Zentall, T. R., Sherburner L. M, & Steirn J. N. (1992). Development of Excitatory Backward Associations during the Establishment of Forward Associations in a Delayed Conditional Discrimination by Pigeons. Animal Learning and Behavior, 20, 199-206. [ Links ]

