










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkUniversitas Psychologica
Print version ISSN 1657-9267
Univ. Psychol. vol.8 no.2 Bogotá May/Aug. 2009
Auditive Discrimination of Equine Gaits by Parade Horses*
Discriminación auditiva de pasos equinos por caballos de paso fino
DUILIO CRUZ-BECERRA **
Universidad Católica de Colombia, Bogotá, Colombia
ENRIQUE BURUNAT-GUTIÉRREZ ***
Universidad de La Laguna, Tenerife, España
ALDO HERNÁNDEZ-BARRIOS ***
Universidad Católica de Colombia, Bogotá, Colombia
ANDRÉS MANUEL PÉREZ-ACOSTA ****
Universidad del Rosario, B ogotá, Colombia
*Research article. Authors wish to thank the support of the Catholic University of Colombia (particularly to Escuela de Tecnologías, and Facultad de Psicología), La Portada (fine-step horse breeders of Colombia), and Fedequinas (Colombian National Federation of Equine Associations). Also we thank to Dr. Niamh Thornton (University of Ulster, UK) and Dr. Luis Fernando Rodríguez-Ramos (Astrophysical Institute of the Canary Islands, Spain) for their support in translation and revision of this manuscript.
**Laboratorio de Psicología Experimental, Facultad de Psicología, Universidad Católica de Colombia, diagonal 47 # 15-50. Bogotá D. C., Colombia. Correo electrónico: ahernandez@ucatolica.edu.co
***Departamento de Psicobiología, Facultad de Psicologia, Universidad de La Laguna, Campus de Guajara. 38071 • La Laguna, Tenerife, Spain. Correo electrónico: eburunat@ull.es
***Laboratorio de Psicología Experimental, Facultad de Psicología, Universidad Católica de Colombia, Calle 47 # 13-54. Bogotá, D. C., Colombia. Correo electrónico: aldohb@ucatolica.edu.co
**** Corresponding Author. Programa de Psicología, Facultad de Medicina, Universidad del Rosario. Carrera 24 No. 63C-69, Quinta de Mutis, Bogotá. Correo electrónico: andres.perez15@urosario.edu.co
Recibido: octubre 17 de 2008 | Revisado: enero 6 de 2009 | Aceptado: enero 13 de 2009
ABSTRACT
The purpose of this study was to examine parade horses' auditory discrimination among four types of equine gaits: paso-fino ("fine step"), trote-reunido ("two -beat trot"), trocha ("trot"), and galope -reunido ("gallop"). Two experimentally naive horses were trained to discriminate the sound of their own gait (paso-fino or fine step), through an experimental module that dispensed food if the subject pressed a lever after hearing a sound reproduction of a particular gait. Three experimental phases were developed, defined by the period of exposure to the sounds (20, 10, and 5 seconds, respectively). The choice between pairs of sounds including the horse's own gait (fine step and two-beat trot; fine step and gallop; and fine step and trot) was reinforced differentially. The results indicate that the fine step horses are able to discriminate their own gait from others, and that receptivity to their own sounds could be included in their training regime.
Key words authors Instrumental Conditioning, Discriminative Learning, Auditory Stimuli, Equine Gaits, Horses.
Key words plus Operant Conditioning, Discrimination Learning, Auditory Stimulation, Horses.
RESUMEN
Esta investigación buscó examinar la discriminación auditiva de cuatro pasos equinos: paso-fino, trote-reunido, trocha y galope-reunido. Dos caballos de paso fino colombiano, experimentalmente ingenuos, fueron entrenados para discriminar el sonido de su propio paso (paso-fino), a través de un módulo experimental que dispensó comida si el sujeto presionaba una palanca, luego de escuchar la reproducción sonora de un paso particular. Se efectuaron tres fases experimentales, definidas por el período de exposición a los sonidos (20, 10 y 5 segundos respectivamente). Se reforzó diferencialmente la elección entre pares de sonidos que incluían el propio paso del caballo (paso-fino vs. trote-reunido; paso-fino vs. trocha; paso-fino vs. galope reunido). Los resultados indican que los caballos de paso fino son capaces de discriminar su propio paso de otros y que la receptividad a sus propios sonidos podría ser incluida en sus regímenes de entrenamiento, con el fin de evitar el uso de estimulación aversiva para tal fin.
Palabras clave autores Condicionamiento instrumental, aprendizaje de discriminación, estímulos auditivos, marchas equinas, caballos.
Palabras clave descriptores Condicionamiento instrumental, aprendizaje de discriminación, estímulos auditivos, caballos.
Introduction
Horses' ability to connect related environmental events has been experimentally evaluated through classical and instrumental conditioning experiments (Nicol, 2002). In particular, instrumental conditioning establishes correspondences between signals (discriminative stimuli), specific actions by the animal (response), and consequent rewards (reinforcers). These types of connections, carried out in controlled environments, simulate the environmental signals that control animal behaviour (see, Dinsmoor, 1995a and b). However, not all stimuli are equally conditionable and, in consequence, it is not always possible to the animal discriminate between them (see Mazur, 1998; Schwartz, 1989). Some researchers of animal behaviour (see, Klein & Mowrer, 2001; Staddon, 2003) have theorized about the biological influence of the stimuli in terms of their relevance on the phylogeny of the species.
In the research on horse's discrimination abilities different types of auditory and visual stimuli have been applied, including some spatial markers (Bubnalittitz, Weinberger & Windischbauer, 1995; Smith & Goldman, 1999; Timney & Keil, 1992), size of the stimuli (Hall, Cassaday & Derrington, 2003), blinking (Geisbauer, Griebel, Schmid & Timney, 2004), selection of response key (Dougherty & Lewis, 1991) and stability of learning experiences over time (McCall & Burgin, 2002). According to the results of these experiments, horses are able to discriminate a wide variety of stimuli types in accordance with the formal conditions of the events.
Relational or symbolic learning has also been demonstrated in horses. For example, Hanggi (2003) investigated the longitude of concepts they can learn (relations among diverse characteristics in a collection of stimuli). His results suggest that these animals are capable of understanding the relation between five types of stimuli, exceeding the learning abilities that minders and trainers had assumed. Research studies in comparative biology and psychology have focused on proving the principles of associative learning, using contingent chains of stimuli, response and rewards (usually food). However, the effects of punitive stimuli have not been systematically evaluated, as well as self-discrimination and arbitrary rewards (Cooper, 1998).
Self-discrimination, just as perception in general, has evolutionary value in an animal adjustment to environmental conditions (Staddon, 2003). In this evolutionary sense, Minetti, Ardigo, Reinach and Saibene (1999) evaluate the relationship between physical work and energy consumption in different types of horse gaits (walk, trot, and gallop). Concluding from their experimental findings, the authors indicate that the type of gait is an evolutionary strategy which optimizes the relationship between speed and energy expended. In this study, results were taken from markers on the spinal column of the subjects. The movement patterns of these markers systematically changed in accordance with the animal's gait. These findings are consistent with those found by Klapp (2004), who compared the movement patterns of untrained and trained horses. Untrained horses presented greater frequency of transversal movements in their lumbar region, in contrast with those which had been trained.
Since Minetti et al. (1999) and Klapp's (2004) findings, the role of synesthesia in learning as well as the development of the gait in horses (Clayton, 1994; Ishii, Amano & Sakuraoka, 1989) has been taken into account. The gait is the combined rhythmic movement of the extremities of the horse. Walking, trotting and galloping are the most common gaits. However, in mountainous regions, people prefer horses whose gaits characterized by the speed of their movement (their four feet move at least two times per second), and by the fact that at any moment one or more of their extremities are in contact with the ground; this is the case of so-called fine step horses. Some of these horses spontaneously show their characteristic movement from youth, while others only demonstrate this after being trained by experienced riders.
Synesthesic self-discrimination, measured with receptors located in muscular joints, tendons and other joints (see, Santa Cruz & Burunat, 2004) plays a facilitating role in learning and the development of gaits in the training of fine step horses. Many other stimuli, like the sound generated by different movements, have not been taken into account in research on sensory discrimination.
The aim of this study was to evaluate the capability of fine step horses in auditory discrimination among different types of gaits (fine step, two beat trot, gallop and trot), evaluated through the speed of acquisition of controlled stimuli in an instrumentally conditioned situation. An eventual finding that these horses have the ability to distinguish between different gaits could provide, not only a better understanding of this species, but also useful elements in the training based on their self- discrimination.
Method Subjects
The experiment was carried out on two five year old horses (Equus caballus), experimentally naïve, registered and classified genotypically as Colombian fine-step horses by the Colombian National Federation of Equine Associations. At the beginning of the experiment, both horses walked adequately and the veterinary medical exam didn't show any sickness or damage in their auditory systems.
Apparatus and stimuli
The experiments took place in a 500 m2 enclosure with access to an experimental module of 9 m2, constructed in a farm near the School of Technology of the Catholic University of Colombia campus, with available water and kikuyu fodder (Pennisetum clandestinum Hoechst) ad libitum. In one corner of the module a metal lever was installed, measuring 20 cm long and 1.25 cm in diameter. This lever went through the wooden wall and was kept in place at the end of the last 10 cm by a spring on the outside. The horse could move the internal end of the lever by pushing against it using its upper incisors using a minimum force of 2 Kilograms onto a disc (5 cm long x 3.75 cm diameter), moving it 30 cm upwards (leverage). Every press-level activated a food dispenser installed besides the disc that dispenses 7 grams of food grains in a plastic container. The movement of the disc and the force required to move the lever were graduated thereby facilitating the operation of the instrument in the learning phase.
To register the number of responses in the different sound intervals or in the periods of silence, two impulse counters were used (Lafayette Instrument model 58022). A GraLab model 171 timing device, alongside an electronic circuit adapted from a microcontroller Freescale model GP32, controlled the synchronization of the sound intervals and the rate of supply of the dispenser.
Auditory stimuli, the sounds generated by the fine step, two-beat trot, trot, and gallop horse gaits, were obtained through the use of a portable recorder installed onto the belly of the test horses while they were being ridden by professional riders. The best recordings were extracted and converted into Microsoft Windows Media Audio ® files which were played on a Hewlett Packard Pavilion Model ZV5000 laptop with a 1000 watt amplifier and two speakers located at either side of the food dispenser. The sound levels were kept between 60-70 decibels by a Yu Fong YF-20 sound level controller.
Procedure
Habituation and shaping
Before starting the experiment, the subjects stayed for 15 days in the enclosure with available feed and water. Once the experiment began the subjects were moved to the experimental module twice a day (from 06:00 to 07:00 and from 16:00 to 17:00) with 2 kg of grain in a trough. The horses were trained to push the lever to obtain food in three one-hour sessions, through the procedure of successive approximations to desired request, by gradually increasing the length of movement of the disc and the force of resistance of the lever.
Pre-training
During a pre -training period the connection of the sound of the fine-step gait and the food was established. The fine-step gait sound (Pf) was played in 30 second periods, during which time the movement of the lever was rewarded with 7 gm of grain. The Pf periods were separated by 30 seconds of silence; during which the press-lever was not rewarded with food. When the subject demonstrated the ability to discriminate 90% or more of the events, the phase was considered to be completed.
Initial training with response cost
In this phase periods of silence were replaced with sounds of other gaits: trot (Tc), two beat trot (Tt) and gallop (G), and intervals between periods were reduced from 30 to 20 seconds. Any response to non-Pf sound reinitiated the 20 seconds interval, resulting in a reduction of the reward. Each hour session included a pair of stimuli: Pf and Tt, Pf and Tc, or Pf and G, consecutively assigned to morning or afternoon sessions. When the animal discriminated at least 90% the phase was considered complete and the rest of training stages continue only with these pairs of sounds. Once finished the three stages with one pair, a new pair was introduced and the training continued. Unlike the other phases of the experiment, in this phase the session ended after 45 periods of 15 seconds of Pf (others last one hour).
Discrimination between gaits emitted in random periods of 10 seconds
In this phase 9 intervals of 10 seconds of Pf were randomly combined with 9 intervals of 10 seconds of one of the other gaits, keeping 18 intervals of 10 seconds of silence between the sound intervals. The aim of this phase was to counter-rest any possibility of a temporal learning effect based on the previous phase, given that the 20 seconds intervals were maintained during the entire session. To avoid repeating a sequence from the earlier phase, two groups of three different triads built on the order of the gaits: one made up of one Pf interval and two of one of another gait, and other made up of two Pf interval and one of the another gait (separated by intervals of silence). A sequence made of six triads randomly chosen was played alternating groups of triads. This procedure was repeated for each one of the three pairs composed by Pf and one of the other three gaits (Tc, Tt y G).
Discrimination among the sounds of gaits played at random intervals of 5 seconds
The final phase was based on a sequence played at random, made up of 12 intervals of 5 seconds of Pf, 12 intervals of 5 seconds of another gait, and 24 intervals of silence of 5 seconds which separated the sounds. To guarantee the balance of sounds over the sequence 6 groups of intervals were built, each one made up of two periods of Pf and two of another gait, separated by silences. The groups were mixed randomly to establish a sequence to be played during each one hour session until a 90% of discrimination was achieved. The procedure was iterated for each one of the 3 pairs of gaits: Pf and Tt, Pf and Tc, Pf and G.
Statistical analysis
For the analysis of the data a Spanish language edition SPSS version 11 database was created. Using this program, descriptive statistics were elaborated, in particular absolute frequencies which are expressed graphically in the tables below. Another type of differential statistics in the middle range was not analyzed because of the size of the sample and the quantitative characteristic of the variables.
Ethical issues
This experiment did not imply the use of aversive stimuli or any kind of invasive treatments. Possible discomfort to the animals was minimized, following the international ethical regulations to the experimental research with animals (Sherwin et al., 2003).
Results
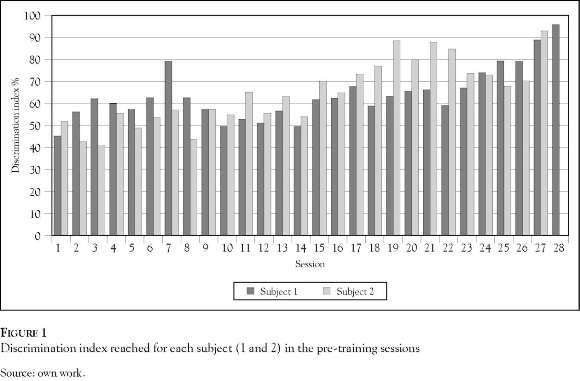
The experimental subjects finished their pre-training after 28 sessions (subject 1) and 27 sessions (subject 2), over the course of two weeks. Figure 1 illustrates the percentage of precision (index of discrimination) in the discrimination of the Pf sound. Although Subject 1 took one more session to exceed the 90% discrimination, the average of the two subjects over the course of the sessions is very similar, 63.8% and 64.5% respectively. Both subjects maintained the index of discrimination above 70% after session 23.
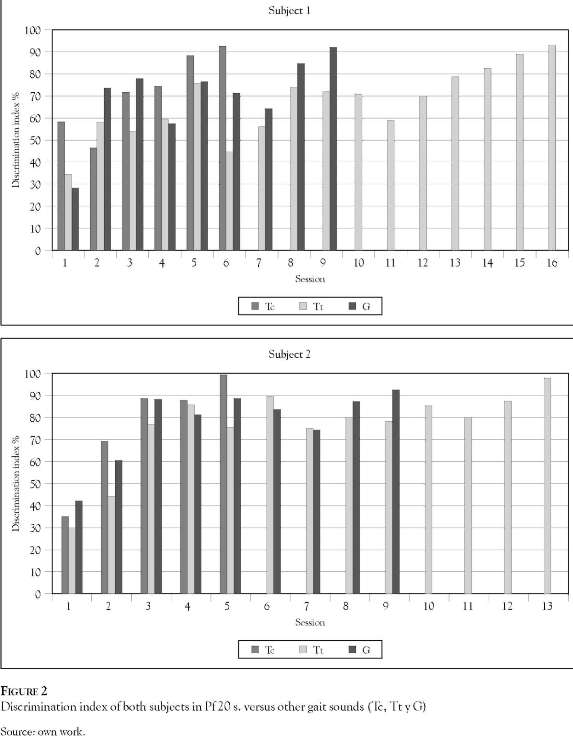
The discrimination of sounds lasting 20 seconds of Pf and of other gaits (Tt, Tc and G) is illustrated in Figure 2. Subject 1 (panel A) and subject 2 (panel B) display similar results discriminating other gaits from the Pf sound. The complete discrimination of the Tt sound was achieved between sessions 4 and 6, for subjects 1 and 2 respectively, while in session 9 both subjects achieved stability in the discrimination of sound G. The horses required longer training to distinguish Tc from the Pf - between 13 and 16 sessions.
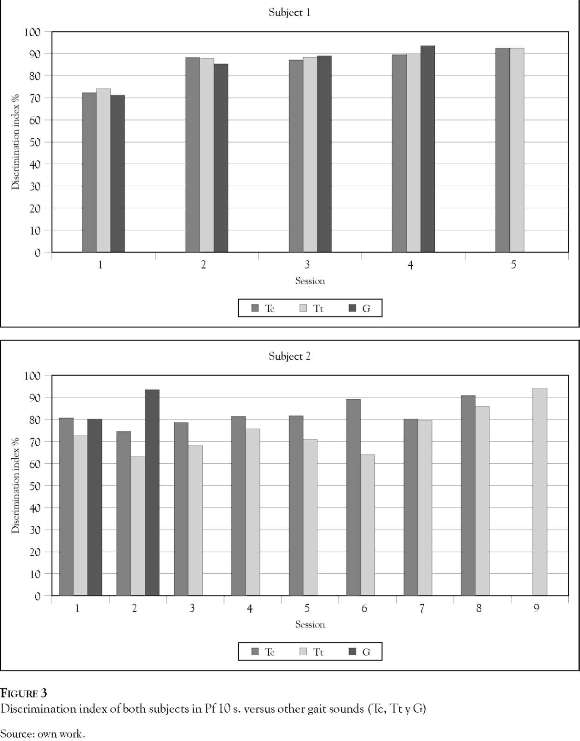
Panel A Figure 3 shows the sessions required by subject 1 to achieve an index of discrimination above 90% with stimuli periods of 10 seconds. The Tt and Tc gaits required 5 sessions, while G gait required 4 sessions. That is one session difference in achieving discrimination. Subject 2 (Panel B Figure 3) required more sessions to reach the level of discrimination between gaits: 2 for G, 8 for Tt and 9 sessions for Tc. However, it was expected that both subjects would discriminate G first, taking longer to discriminate between the other two (Tt and Tc); it is worth noting that discrimination by both subjects between Tt and Tc gaits occurred in sequential or identical sessions.
In Figure 4 Panel A it is clear that subject 1 only needed 3 sessions to discriminate between Tc and G and 2 sessions for Tt, while subject 2 (Panel B) reached a level of discrimination of over 90% for G and Tc in one session, and by the second session could discriminate Tc.
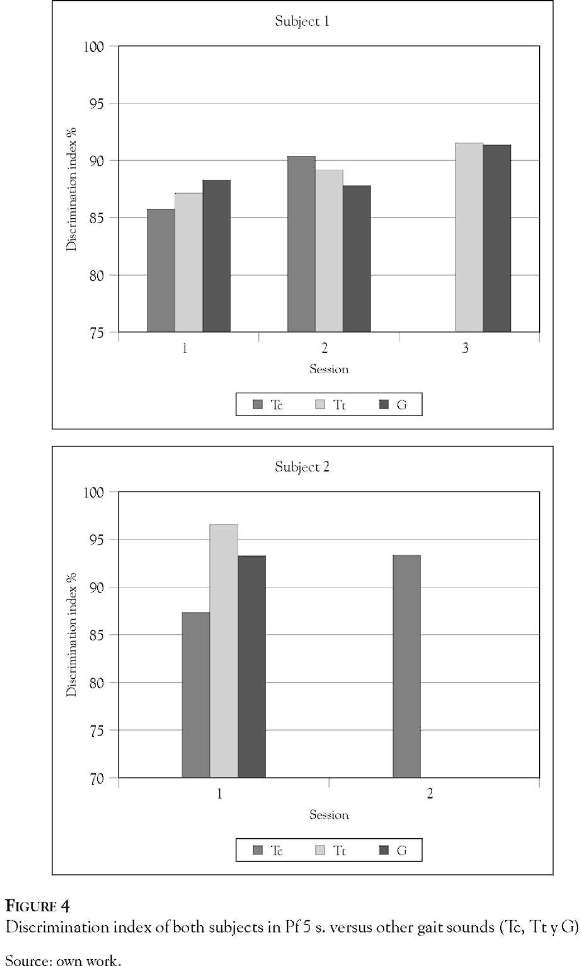
Figure 5 illustrates that the progressive reduction in periods of exposure to discriminative stimuli (20, 10 and 5 seconds corresponding to the different training phases) reduced the number of training sessions needed by the subjects to reach a 90% level of discrimination. In addition, it is possible to see higher levels of difficulty for both subjects to discriminate the sound of trot in contrast with that for trot and gallop.
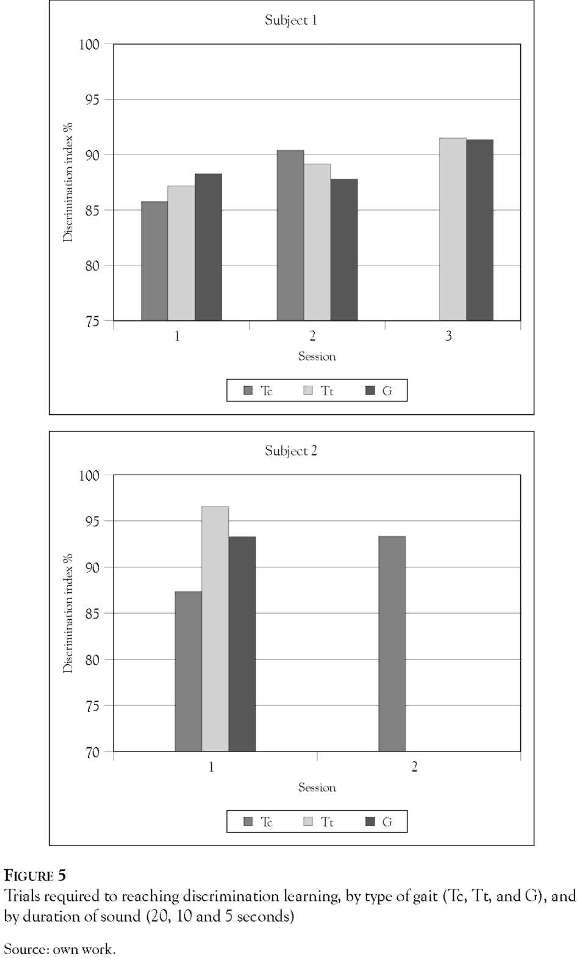
Discussion
Previous studies have shown that horses can discriminate pure tone frequencies (Heffner & Heffner, 1983, 1986); however, the sound stimuli which control the horses' conduct are complex; neighs, whinnies, hooves' stomping, swishing tails, rider's sounds, etc. The results of this study lead us to conclude that, on one hand the horses' sensitivity to the sound generated by the fine step gait can lead them to control the conduct of the press-lever with 90% efficacy (Figure 1). On the other hand, fine step horses can distinguish the sound that they produce walking, from those generated by other gaits (trot, two beat trot and gallop) (Figures 2, 3 and 4). In these last illustrations it is obvious that the trot and fine step sounds are very similar for the horse, since both subjects required a greater number of training sessions to distinguish both sounds 90% of the time. This difficulty in distinguishing between the two gaits can be attributed to the similarity in the number of sound pulses per second; 11 for fine step and 13 for the trot. However, the difference is based on the origins of the sound; fine step is generated through the striking of the ground of each lateral pair of legs one after the other, fore-leg first and then hind-leg, while the trot is a result of a diagonal rhythm of the striking of the right fore-leg followed by that of the left hind-leg or vice versa.
The three phases of training resulted in a progressive reduction in the presentation time of the sound stimuli (20, 10 and 5 seconds), which resulted in an increase in the specificity of the discriminatory stimuli, which also meant an increase in the reinforcing rewards in food and a decrease in the level of awareness to environmental stimuli which accompanied the sound of the gaits. Despite the increasing complexity of the tasks over the course of the training, the subjects maintained the level of learning achieved in the first phase, composed of 20 seconds of fine step and 20 seconds of another gait. This leads us to believe that the subjects achieved 90% in the level of discrimination between press -lever in function of the characteristics of the sound and not as a result of learning the timings of these intervals.
Both subjects demonstrated similar learning curves; in other words no major differences were evident between the results of the two subjects during their training period (p>0.01). This allows us to conclude that fine step horses have the capacity to learn how to distinguish between different gaits to those of a regular walk, which facilitates the possibility to implement a methodological strategy to evaluate the innate reactions of the individual towards these sound stimuli, in function with the learning speed, and, thereby, find a level of innate alertness that could be the closest description of what is called 'brio' in the equine world.
A two weeks period is a normal time interval in animal behaviour research. It corresponds to the underlying neurobiological pattern. Even in early studies such as Pavlov's classical conditioning, this period was found to be usual (Pavlov, 1927), and alongside other factors which have been described in our experiments on the result of pharmacological stimuli on the central nervous system as "pharmacological conditioning" (Burunat, Diaz-Palarea, Castro & Rodriguez, 1987, 1988).
Certain methodological features need to be taken into account when studying fine step horses. The type of lever is one of them. In some preparatory experiments a similar model to that of Dougherty and Lewis (1991) was used (5.1 cm wide, 1.3 cm thick and 1.75 cm long). However, the participating fine step horse subjects simply pushed impulsively the press-lever in some periods. Then the animals went away from both the feed area and the lever without paying attention to the sonic changes in the environment. To protect the teeth and mouth of the animals the metal lever was covered with hard wood and then the size was increased by 10 cm in length and width, despite this the animals continued to move the lever without stopping until they heard the sound periods which matched the delivery of food. In this case the vertical movement of the lever was increased by 30 cm, which meant that the subject had to move the neck and head further; consequently the challenge was increased. The horse had to stay in the feed area where the sound source was located; the high frequencies of the press-lever in irregular periods practically disappeared and the animals were conditioned to the operation of the lever sooner.
The Colombian fine step horses are descents from the Barb line of North Africa. These are considered hot-blooded in their interactions with others of their breed, riders and towards environmental variations. Possibly this condition (which had been label 'brio' in fine step horses) is the reason why these animals push and bite at the lever so hard. This was not found by Dougherty and Lewis (1991) in their experiment.
Heffner and Heffner (1983, 1984, 1986) used a training system whereby the horse had to push its head against a plate and remove it on hearing a sound matched with water in the drinking fountain. Water deprivation induces colic, a cause of death in a large number of horses, and could reduce the commercial value of the fine step horses when become reproductive (because affects phenotype and movements). Therefore, this type of conditioning was abandoned.
Periods of deprivation of food grain used in this experiment were the same as those that the animals were accustomed to when in stables. Given that the session took place from 06:00 to 07:00 and 16:00 to 17:00 the night period of food deprivation exceeded that of the daytime by 4 hours. This difference, which could have generated clear evidence of motivational changes in the frequency of press-lever between the morning and afternoon sessions, meant that the Pf and Tt, Pf and Tc, Pf and G sound variations were equally distributed between AM and PM. Comparisons between the frequencies of press-lever in the morning and afternoon over the course of the training period did not display significant differences, which could be put down to the greater amount of night time motor activity, the daytime need to eat, combined with greater alertness which balanced itself out.
The balanced assignation of pairs of sounds, randomly mixed and assigned morning and evening sessions in the period of discrimination between gaits, prevented any prospective effect in learning one stimuli on the pattern presented in another session, when similar patterns appeared one after another.
Another methodological feature to be controlled relates to discrimination stability after reaching the criteria of 90%. In this sense, once reached this level, two more sessions were carried out where the discrimination levels were over 90%. For this reason they were not taken into account in the analysis of the results.
Our results lead us to conclude that the horses are able to discriminate their own sound sequences when they change from their characteristic gaits. This ability in horses suggests new research questions related to the role that listening may have in the motor movements of a fine step horse training or re-training. This ability to discriminate the sounds that characterize their gait can also assist in their training when young, and could allow selection between the correct motor sequences, which is further facilitated by the genetic makeup of their parents. The ability to discriminate between sounds could be understood as a selection of their own behaviour, a basic form of self-discrimination present in different species (Perez-Acosta, Ben-jumea & Navarro, 2001). To achieve a definitive demonstration of self-awareness, another study is required in which the specific stimuli (horse gait in this case) is emitted by the subject itself and later associated with an arbitrary symbol or "label of the own behaviour" (Dymond & Barnes, 1997).
As a consequence of this research, whose theoretical and economic outcomes cannot yet be fully quantified, we hope to contribute to the improvement of fine step horses training, thus avoiding the usual practice of more aversive and less efficient techniques that cause difficulties in learning that impend or retard training.
References
Bubnalittitz, H., Weinberger, H. & Windischbauer, G. (1995). Determination of the upper auditory threshold in horses by operant conditioning. Wiener Tierarztliche Monatsschrift, 82, 259-263. [ Links ]
Burunat, E., Diaz-Palarea, M. D., Castro, R. & Rodriguez, M. (1987). Conditioned rotational response in nigrostriatal system-lesioned rats: The origin of undrugged rotational response. Life Sciences, 41, 1861-1866. [ Links ]
Burunat, E., Castro, R., Diaz-Palarea, M. D. & Rodriguez, M. (1988). Conditioning of the early behavioral response to apomorphine in the rotational model of Parkinson's disease. European Journal of Pharmacology, 145, 323-327. [ Links ]
Clayton, H. M. (1994). Comparison of the stride kinematics of the collected, working, medium, and extended trot in horses. Equine Veterinary Journal, 26, 173-174. [ Links ]
Cooper, J. J. (1998). Comparative learning theory and its application in the training of horses. Equine Veterinary Journal, 27, Supplement, 39-43. [ Links ]
Dinsmoor, J. A. (1995a). Stimulus control: Part I. The Behavior Analyst, 18, 51-68. [ Links ]
Dinsmoor, J. A. (1995b). Stimulus control: Part II. The Behavior Analyst, 18, 253-269. [ Links ]
Dymond, S. & Barnes, D. (1997). Behavior analytic approaches to self-awareness. Psychological Record, 47, 181-200. [ Links ]
Dougherty, D. M. & Lewis, P (1991). Stimulus generalization, discrimination learning, and peak shift in horses. Journal of the Experimental Analysis of Behavior, 56, 97-104. [ Links ]
Geisbauer, G., Griebel, U., Schmid, A. & Timney, B. (2004). Brightness discrimination and neutral point testing in the horse. Canadian Journal of Zoology, 82, 660-670. [ Links ]
Hall, C. A., Cassaday, H. J. & Derrington, A. M. (2003). The effect of stimulus height on visual discrimination in horses. Journal of Animal Science, 81, 1715-1720. [ Links ]
Hanggi, E. B. (2003). Discrimination learning based on relative size concepts in horses (Equus caballus). Applied Animal Behaviour Science, 83, 201-213. [ Links ]
Heffner, H. E. & Heffner, R. S. (1984). Sound localization in large mammals: Localization of complex sound by horses. Behavioral Neuroscience, 98, 541-555. [ Links ]
Heffner, R. S. & Heffner, H. E. (1983). Hearing in large mammals: Horses (Equus caballus) and cattle (Bos taurus). Behavioral Neuroscience, 97, 299-309. [ Links ]
Heffner, R. S. & Heffner, H. E. (1986). Localization of tones by horses: Use of binaural cues and the superior olivary complex. Behavioral Neuroscience, 100, 93-103. [ Links ]
Ishii, A., Amano, K. & Sakuraoka, H. (1989). Kinematic analysis of horse gait. Bulletin of Equine Research Institute, 26, 1-9. [ Links ]
Klapp, M. A. (2004). Influence of the rider on the variability of the equine gait. Human Movement Science, 23, 663-671. [ Links ]
Klein, S. B. & Mowrer, R. R. (Eds.). (2001). Handbook contemporary learning theories. Mahwah, NJ: Erlbaum. [ Links ]
Mazur, J. E. (1998). Learning and behavior. Englewood Cliffs, NJ: Prentice-Hall. [ Links ]
McCall, C. A. & Burgin, S. E. (2002). Equine utilization of secondary reinforcement during response extinction and acquisition. Applied Animal Behaviour Science, 78, 253-262. [ Links ]
Minetti, A. E., Ardigó, L. G., Reinach, E. & Saibene, F. (1999). The relationship between mechanical work and energy expenditure of locomotion in horses. Journal of Experimental Biology, 202, 23292338. [ Links ]
Nicol, C. J. (2002). Equine learning: Progress and suggestions for future research. Applied Animal Behaviour Science, 78, 193-208. [ Links ]
Pavlov, I. (1927). Conditioned reflexes: an investigation of the physiological activity of the cerebral cortex. Downloaded on October 10, 2008, from htt://psychclassics.yorku.ca/Pavlov/ [ Links ]
Pérez-Acosta, A. M., Benjumea, S. & Navarro, J. I. (2001). Autoconciencia animal: estudios sobre autodiscriminación condicional en varias especies [Animal self-awareness: Research on conditional self-discrimination in several species]. Revista Latinoamericana de Psicología, 33, 311-327. [ Links ]
Santa Cruz, R. & Burunat, E. (2004). Bases biológicas y fisiológicas del movimiento [Biological and physiological bases of movement]. Tenerife, Spain: Arte Comunicación Visual. [ Links ]
Schwartz, B. (1989). Psychology of Learning and Behavior. New York: Norton. [ Links ]
Sherwin, C. M., Christiansen, S. B., Duncan, I. J., Erhard, H., Lay, D., Mench, J., O'Connor, C. & Petherick, C. (2003). Guidelines for the ethical use of animals in applied animal behaviour research. Applied Animal Behaviour Science, 81, 291-305. [ Links ]
Smith, S. & Goldman, L. (1999). Color discrimination in horses. Applied Animal Behaviour Science, 62, 13-25. [ Links ]
Staddon, J. E. R. (2003). Adaptive behavior and learning. Cambridge, MA: Cambridge University Press. [ Links ]
Timney, B. & Keil, K. (1992). Visual acuity in the horse. Vision Research, 32, 2289-2293. [ Links ]
