










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkUniversitas Psychologica
Print version ISSN 1657-9267
Univ. Psychol. vol.9 no.3 Bogotá Sept./Dec. 2010
Efectos de la administración de LY354740, un agonista selectivo del grupo II de receptores metabotrópicos de glutamato, sobre la conducta agresiva en ratones*
Effects of LY354740, a Selective Agonist for Glutamate Metabotropic Receptors of Group II, on Aggressive Behaviour in Mice
VANESSA DE CASTRO **
MERCEDES MARTÍN-LÓPEZ ***
JOSÉ FRANCISCO NAVARRO ****
* Artículo de investigación.
** Universidad de Málaga, España, Departamento de Psicobiología. Facultad de Psicología. Campus de Teatinos s/n 29071. Málaga, España E-mail: navahuma@uma.es
*** Universidad de Málaga, España, Departamento de Psicobiología. E-mail: mmmartin@uma.es
**** Universidad de Málaga, España, Autor para correspondencia. E-mail: vanedcc@hotmail.com
Recibido: enero 10 de 2010 Revisado: abril 10 de 2010 Aceptado: abril 19 de 2010
Para citar este artículo
De Castro, V., Martín-López, M. & Navarro, J.F. (2010). Efectos de la administración de LY354740, un agonista selectivo del grupo II de receptores metabotrópicos de glutamato, sobre la conducta agresiva en ratones. Universitas Psychologica,9 (3), 617-625.
Resumen
Estudios recientes han demostrado una implicación de los receptores metabotrópicos mGlu1 y mGlu5 en la regulación de la conducta agresiva. Este trabajo examina el efecto de la administración de LY354740 (4-16 mg/ kg ip), un agonista selectivo de los receptores metabotrópicos del grupo II (mGlu2/3), en encuentros agonísticos entre ratones macho, utilizando un modelo de agresión inducida por aislamiento. Treinta minutos tras la administración del fármaco, se llevaron a cabo interacciones agonísticas de 10 min de duración entre un animal aislado y un oponente anósmico en un área neutral. Dichos encuentros fueron grabados, para su posterior análisis etológico, estimándose el tiempo pasado por los ratones en cada una de diez categorías conductuales. LY354740 (12 y 16 mg/kg) redujo significativamente las conductas ofensivas, sin afectar la motilidad, en comparación con el grupo control. Estos resultados sugieren una implicación de los receptores mGlu del grupo II, en la modulación de la agresión.
Palabras clave autores : Agresión, conducta agonística, mGlu2/3, ratón, receptores metabotrópicos de glutamato.
Palabras clave descriptor : Receptores de glutamato metabotrópico, agresividad (psicología), ratas-psicología.
Abstract
Recent studies have demonstrated that glutamate metabotropic receptors mGlu1 and mGlu5 are involved in the regulation of aggressive behaviour. This study examines the effect of the administration of LY354740 (4-16 mg/kg i.p.), a selective group II metabotropic receptors agonist (mGlu2/3), using an isolation-induced aggression model. Individually housed mice were exposed to anosmic opponents 30 min after drug administration. Ten min of diadic interactions were staged between a singly housed and an anosmic mouse in a neutral area. The encounters were videotaped and the accumulated time allocated by subjects to ten broad behavioural categories was estimated using an ethologically based analysis. LY354740 (12 and 16 mg/ kg) significantly reduced offensive behaviours, without affecting immobility, as compared with the control group. These results suggest an implication of mGlu group II receptors in the modulation of aggression.
Keywords authors : Aggression, Agonistic Behaviour, Metabotropic Glutamate Receptors, mGlu2/3, Mice.
Keywords plus : Receptors, Metabotropic Glutamate, Aggressiveness (Psychology), Rats-Psychology.
Introducción
El L-Glutamato es el principal neurotransmisor excitatorio del sistema nervioso central (SNC) (Nedergaard, Takano & Hansen, 2002). Media su acción a través de dos grupos de receptores: los receptores ionotrópicos (iGluR), que son receptores de canales iónicos controlados por ligando, responsables de la transmisión sináptica rápida (NMDA, AMPA y Kainato), y los receptores metabotropicos (mGluR), que forman parte de la superfamilia de receptores ligados a la proteína G y modulan la transmisión sináptica lenta (Kew & Kemp, 2005; Ozawa, Kamiya & Tsuzuki, 1998). Mediante técnicas de clonación molecular, se han identificado ocho subtipos de receptores metabotrópicos de glutamato (mGluR1-8). éstos se han clasificado en tres grupos en función de su homología de aminoácidos, sus mecanismos de transducción de señales y su farmacología (Ferraguti & Shigemoto, 2006). El grupo I incluye los receptores mGlu1 y mGlu5, y están ligados a la hidrólisis de fosfoinositoles; el grupo II está formado por los receptores mGlu2 y mGlu3, y están ligados negativamente a la adenilciclasa, mientras que el grupo III lo constituyen los receptores mGlu4, mGlu6, mGlu7 y mGlu8, que inhiben igualmente la formación de AMPc.
Estudios inmunohistoquímicos y de hibridación in situ han mostrado diferentes patrones de distribución para los ocho subtipos de mGluR en el SNC (Luján-Miras, 2005). Los receptores del grupo II se encuentran localizados tanto a nivel presináptico como postsináptico, además de localizarse en la glía. Así, la activación presináptica de los receptores mGlu2/3 modula de forma negativa la liberación de glutamato previniendo su excesiva liberación -funcionaría como autorreceptor-(Anwyll, 1999), pero también puede regular la liberación de otros neurotransmisores (Cartmell & Schoepp, 2000). Además, la activación postsináptica puede regular la excitabilidad neuronal, a través de la modulación de las funciones de los canales iónicos (Anwyll, 1999). No obstante, es la actividad presináptica la que convierte a estos receptores en atractivas dianas terapéuticas (Schoepp, 2001). Por otro lado, los receptores metabotrópicos del grupo II se expresan principalmente en los cuerpos celulares de la mayoría de las estructuras claves involucradas en la recepción, procesamiento y transmisión de la información de las respuestas emocionales, como la ansiedad y la agresión (Gu et al., 2008).
Diversos sistemas de neurotransmisión han sido involucrados en la regulación de la conducta agresiva, incluyendo la serotonina (Navarro & Maldonado, 2004), dopamina (Manzaneque & Navarro, 1999; Martín-López, Puigcerver, Vera & Navarro, 1993), GABA (Martín-López & Navarro, 2002), opiáceos (Espert, Salvador, Navarro & Simón, 1993) y GHB (Navarro & Pedraza, 1996; Pedraza, Dávila, Martín-López & Navarro, 2007), entre otros. En los últimos años, el papel desempeñado por el glutamato en la modulación de la agresión, está recibiendo un interés creciente. Así, numerosos estudios han constatado una implicación de los receptores ionotrópicos de glutamato en la conducta agresiva (Belozertseva & Bespalov, 1999; Lumley et al., 2004; Navarro, Burón & Martín-López, 2007; Vekovischeva et al., 2004; Vekovischeva, Aitta-Aho, Verbitskaya, Sandnabba & Korpi, 2007). En contraste, el papel que juegan los receptores mGlu es menos conocido. Recientemente, se ha demostrado que los receptores del grupo I modulan la conducta agresiva. En concreto, la administración aguda de antagonistas selectivos de los receptores mGlu5 (MPEP; 5-25 mg/kg ip) y mGlu1 (JNJ16259685; 0.125-8 mg/kg) produjo una potente reducción de las conductas ofensivas (amenaza y ataque), sin afectar la motilidad, en un modelo animal de agresión inducida por aislamiento, indicando un papel de dichos receptores en la regulación de la agresión (Navarro, De Castro & Martín-López, 2008; Navarro, Postigo, Martín-López & Burón, 2006). Asimismo, Navarro, De Castro y Martín-López (2009b) han sugerido, recientemente, que los receptores mGlu7 (pero no los receptores mGlu8) podrían estar también implicados en la modulación de la agresión.
En la misma línea, los resultados obtenidos en un experimento preliminar con un ligando selectivo de los receptores mGlu2/3 (LY379268) sugieren que estos receptores podrían estar también implicados en la agresión (Navarro, Luque & Martín-López, 2009a). Así, encontraron un efecto antiagresivo tras la administración aguda de LY379268, pero con un margen muy estrecho de dosis. Concretamente, observaron una reducción significativa de la conducta de ataque con las dosis de 2 y 4 mg/kg pero, con la dosis más alta, el efecto antiagresivo se acompañó de un incremento significativo de la inmovilidad, indicando una acción no selectiva del fármaco para esa dosis. El objetivo del presente trabajo es intentar confirmar la posible implicación de los receptores mGlu2/3 en la modulación de la conducta agresiva. Para ello utilizamos el compuesto LY354740, un potente y selectivo agonista de los receptores metabotrópicos del grupo II, con un EC50 de 5 y 24 nM para los receptores mGlu2 y mGlu3, respectivamente (Schoepp et al., 1997).
Método
Animales
Se utilizaron 168 ratones albinos machos de la cepa Swiss OF. 1 (Harlam Ibérica, Barcelona). A la llegada a nuestro laboratorio, todos los animales fueron alojados en grupos de 5, durante una semana, para permitir su adaptación a nuestras instalaciones y al ciclo de luz/oscuridad impuesto (luz: 20.00h -8.00h). Transcurrido el período de adaptación, la mitad de los animales fueron empleados como animales experimentales y controles, siendo alojados individualmente (durante un período de 30 días) en jaulas de plástico transparentes (Tecniplast-Letica, Madrid) de 24 x 13,5 x 13 cm. Una vez finalizado el período de aislamiento, estos animales fueron distribuidos aleatoriamente a las distintas condiciones experimentales. En total, se formaron cinco grupos: un grupo control (vehículo -suero fisiológico más DMSO-) y cuatro experimentales (vehículo con las diferentes dosis de fármaco). Los animales restantes se utilizaron como oponentes "anósmicos", y permanecieron alojados en grupos de 5 en jaulas de las mismas características que las anteriores, hasta el momento de la realización de la prueba conductual. Para todos los animales, la comida y la bebida se administraron ad libitum. Las condiciones ambientales del laboratorio donde estaban ubicados los animales fueron cuidadosamente controladas, manteniéndose una temperatura (20 °C) y humedad constantes.
Este experimento se ha realizado de acuerdo con las principales reglas de cuidado y uso de Animales de Laboratorio aprobado por el Consejo Directivo de las Comunidades Europeas del 24 de Noviembre de 1986 (86/609/EEC).
Modelo de agresión inducida por aislamiento y descripción de la anosmia
Existen numerosos modelos de agresión animal (Olivier & Young, 2002), siendo el modelo de agresión inducida por aislamiento uno de los más empleados en el ámbito de la psicofarmacología. El aislamiento durante un periodo de cuatro semanas induce el incremento de la agresión en un animal territorial, como el ratón, conducta que se pone de manifiesto cuando es enfrentado a un oponente anósmico (Brain, 1975; Navarro, 1997). La prueba conductual tiene una duración de 10 minutos, y durante la misma los ratones muestran todo el repertorio de conductas agonísticas, incluidas las conductas ofensivas y defensivas (Miczek, Maxson, Fish & Faccidomo, 2001).
La anosmia se realizó mediante la administración intranasal de una solución al 4% de sulfato de zinc (Laboratorios Sigma) en los días 1 y 3 anteriores al test conductual. Los animales anósmicos son un tipo de "oponente" que elicita las conductas de amenaza y ataque, pero muy rara vez inician estas conductas (Brain, Benton, Childs & Parmigiani, 1981), convirtiéndose, por tanto, en un tipo de oponente estándar que permite evaluar de forma controlada la conducta del animal experimental.
El olfato juega un papel crucial para la discriminación social (Matochik, 1988). Probablemente, la detección de feromonas sea un factor determinante (Martínez, Calvo-Torrent & Paya-Cano, 1997). Así, en un encuentro con un ratón "anósmico" la lucha será siempre unidireccional y fácil de cuantificar (Liebenauer & Slotnock, 1996).
Administración del fármaco
El ligando LY354740 se adquirió comercialmente en los laboratorios Tocris (Madrid). El fármaco fue disuelto en suero fisiológico y dimetilsulfóxido (DMSO) a una concentración del 8.5%. Las dosis empleadas fueron: 4, 8, 12 y 16 mg/kg, seleccionadas con base en diversos estudios conductuales con animales de experimentación (Klodzinska et al., 1999; Moore, Rees & Monn, 1999; Schlumberger et al., 2009). La administración del vehículo y del fármaco se realizó por vía intraperitoneal (ip) en un volumen constante de 10 ml/kg.
Los animales aislados fueron distribuidos al azar, en cinco grupos. Un grupo control (n = 20) que recibió vehículo (suero salino (91.5%) más DMSO (8.5%), y cuatro grupos experimentales (n = 15-20 cada grupo) cuyos animales fueron tratados con las diferentes dosis del fármaco (4, 8, 12 y 16 mg/kg). La administración del fármaco y del vehículo se realizó siempre treinta minutos antes de la prueba conductual.
Evaluación conductual
Los encuentros agonísticos entre el animal aislado y el oponente anósmico, tuvieron lugar en un área neutral. Esta zona neutral consistía en un recipiente de cristal con medidas de 50 x 26 x 30 cm cuya base estaba cubierta de serrín fresco. Antes de comenzar la prueba, los animales permanecían separados en extremos opuestos del recipiente, durante un periodo de adaptación de 1 minuto, tras el cual se retiraba el separador y se daba comienzo el test conductual. Los encuentros fueron grabados con una cámara Sony-V8. Todas las pruebas se realizaron bajo una luz roja, entre la segunda y séptima horas de la fase oscura de la condición de luz. El serrín era renovado después de cada encuentro conductual.
Los encuentros agonísticos fueron analizados utilizando un microprocesador y un programa específico desarrollado para tal fin (Brain, McAllister & Walmsey, 1989), que facilitaba la estimación del tiempo pasado por los animales, en las diez categorías conductuales evaluadas. El análisis de la conducta fue llevado a cabo por un experimentador "ciego" al tratamiento administrado a los sujetos experimentales, siendo evaluada sólo la conducta exhibida por el animal aislado. Las categorías analizadas fueron las siguientes: 1. Cuidado corporal; 2. Escarbar; 3. Exploración no social; 4. Exploración a distancia; 5. Investigación social; 6. Amenaza; 7. Ataque; 8. Evitación/Huida; 9. Defensa/Sumisión; 10. Inmovilidad. Este procedimiento experimental permite una completa cuantificación de los elementos conductuales mostrados por los sujetos, durante el encuentro agonístico.
Análisis estadístico
Con el objeto de establecer si existían diferencias significativas entre los diferentes grupos experimentales (control y tratamiento), en cada una de las categorías conductuales evaluadas, se realizó un análisis de varianza no paramétrico (prueba de Kruskal-Wallis). Para realizar comparaciones entre pares de grupos, se utilizó una prueba no paramétrica para muestras independientes (prueba U de Mann-Whitney).
Resultados
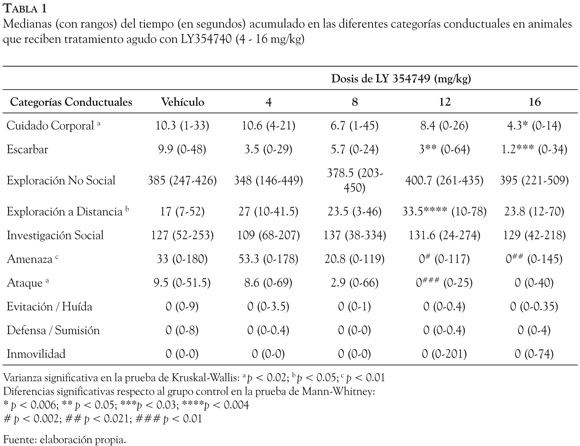
En la Tabla 1, se muestran las medianas (y los rangos) del tiempo acumulado (en segundos), en cada una de las categorías conductuales, para cada grupo de tratamiento. El análisis de Kruskall-Wallis indicó que existían diferencias estadísticamente significativas en las categorías de cuidado corporal, exploración a distancia, amenaza y ataque (p < 0.01 - p < 0.05). La prueba de U Mann-Whitney reveló que la administración de LY354740 reducía de forma significativa el tiempo empleado por los animales en conductas de cuidado corporal (16 mg/ kg: p < 0.006), escarbar (12 mg/kg: p < 0.05; 16 mg/kg: p < 0.03), amenaza (12 mg/kg: p < 0.002; 16 mg/kg: p < 0.021) y ataque (12 mg/kg: p < 0.01), en comparación con los animales del grupo control, mientras que las conductas de exploración a distancia se vieron significativamente incrementadas (12 mg/kg: p < 0.004). Las medianas para las categorías conductuales de evitación/huída, defensa/sumisión e inmovilidad fueron cero para todos los grupos.
Discusión
Los receptores metabotrópicos del grupo II se encuentran localizados tanto post como presinápticamente (Klodzinska et al., 1999). Así, se ha propuesto que la activación de los receptores presinápticos mGlu localizados en los terminales nerviosos glutamatérgicos, causa una disminución de la liberación de glutamato, inhibiendo la transmisión excitatoria. De ahí que los compuestos que actúan sobre los autorreceptores presinápticos, puedan ejercer un antagonismo funcional sobre el sistema glutamatérgico. Además, la estimulación postsináptica de los receptores mGlu2/3 provoca una inhibición de la acumulación de AMPc, en el cerebro. Por lo tanto, independientemente de su localización sináptica, la estimulación de los receptores mGlu del grupo II produce respuestas inhibitorias en el cerebro (Luján-Miras, 2005).
Diversos estudios utilizando técnicas histoquímicas y de autorradiografía han evidenciado una extensa distribución a lo largo del sistema nervioso central (SNC) del ARNm e inmunorreactividad para los receptores mGlu2/3. El ARNm para los receptores mGlu2 se distribuye en regiones más limitadas del SNC, en comparación con el mGlu1, mGlu5 y mGlu3, con una elevada expresión en los cuerpos celulares de la mayoría de las estructuras del sistema límbico del prosencéfalo (incluyendo algunos núcleos talámicos, giro dentado del hipocampo, núcleo mamilar medial y amígdala), así como en la corteza cerebelosa y el bulbo olfatorio. Aunque existe cierto solapamiento, como en el hipocampo y en la amígdala, la expresión del AR-Nm para los receptores mGlu3 parece estar más dispersa (Gu et al., 2008). Muchas de estas regiones cerebrales están comúnmente asociadas con trastornos de ansiedad (Cryan & Dev, 2008). La amígdala es la principal estructura que procesa el condicionamiento de miedo. En modelos animales de estrés agudo y crónico, las neuronas del núcleo basolateral de la amígdala muestran un aumento de respuesta que contribuye al almacenamiento de la memoria de miedo, incrementando la ansiedad y la agresión (Gu et al., 2008). En este sentido, una creciente evidencia indica que la estimulación de los receptores metabotrópicos del tipo II, podría reducir la ansiedad (Bespalov et al., 2008). Concretamente, diferentes estudios muestran que los agonistas de los receptores mGlu2/3, como el LY354740, representan un prometedor tratamiento para la ansiedad y trastornos relacionados (Lee, Duman & Marek, 2006; Linden, Greene, Bergeron & Schoepp, 2004; Schoepp, Wright, Levine, Gaydos & Potter, 2003). No obstante, las implicaciones clínicas de los receptores mGlu2/3 no se circunscriben sólo a la ansiedad/estrés, sino que abarca un amplio número de trastornos psiquiátricos y neurológicos (Marek, 2004), incluyendo la agresividad. En este sentido, estructuras límbicas como la amígdala, la formación hipocampal, el área septal, la corteza prefrontal y el giro cingulado anterior, ejercen también importantes funciones moduladoras sobre la conducta agresiva, siendo el hipotálamo y la sustancia gris periacueductal (PAG) los principales responsables de su expresión (Siegel, Bhatt, Bhatt & Zalcman, 2007; Siever, 2008).
En nuestro estudio, la administración aguda del ligando selectivo LY35470 produjo una reducción significativa de las conductas ofensivas con la dosis de 12 mg/kg (amenaza y ataque), y de 16 mg/ kg (amenaza), sin un aumento concomitante de las conductas de inmovilidad, sugiriendo un perfil antiagresivo específico. Asimismo, la reducción de las conductas ofensivas se acompañó de una disminución de las conductas de escarbar (12 y 16 mg/kg). La categoría conductual de escarbar esté involucrada en la conducta agresiva junto con amenaza y ataque. De hecho, normalmente existe una correlación positiva entre estas categorías conductuales (Kudryavtseva, Bondar & Alekseyenko, 2000). En concordancia con estos resultados, podemos citar el trabajo preliminar de Navarro et al. (2009a), quienes utilizando el ligando LY379268, un agonista selectivo de los receptores metabotrópicos mGlu2/3, describieron una acción antiagresiva con las dosis de 2 y 4 mg/ kg (aunque sólo selectiva con la dosis de 2 mg/ kg), empleando también el modelo de agresión inducida por aislamiento. Además, dicha acción antiagresiva se acompañaba igualmente de una reducción en las conductas de escarbar. Por lo que respecta a la conducta de cuidado corporal, ésta representa una respuesta etológica relevante dentro del repertorio conductual del animal puesto que es altamente influenciable por factores endógenos y exógenos, interpretándose como una reacción al estrés, mientras que la categoría de exploración a distancia es considerada como un anticipo de la conducta social (Gómez, Carrasco & Redolat, 2008). Helton, Tizzano, Monn, Schoepp y Kallman (1998) constataron que el fármaco LY354740 muestra actividad ansiolítica, en modelos animales que son sensibles a las benzodiazepinas, sin producir los indeseables efectos colaterales de éstas, mientras que Klodkinska et al. (1999) observaron que dosis intermedias de LY354740 (0.5 y 1 mg/kg), pero no dosis más altas (2 y 4 mg/kg), tenían un efecto ansiolítico evaluado en el test de Vogel. No obstante, en nuestro trabajo, con las dosis utilizadas (4, 8, 12 y 16 mg/kg), no apreciamos una acción ansiolítica, puesto que las conductas de exploración no social y, especialmente, de investigación social, cuyo incremento se asocia tradicionalmente a una reducción de la ansiedad (Brain, Kusumorini & Benton, 1991; Navarro & Maldonado, 2004), no se han visto significativamente afectadas con ninguna de las dosis empleadas.
En conclusión, los resultados obtenidos en el presente trabajo sugieren que los receptores metabotrópicos de glutamato mGlu2/3 podrían estar implicados en la modulación de la agresión. Son necesarios estudios adicionales para intentar clarificar el papel diferencial de cada uno de los receptores por separado (mGlu2 vs. mGlu3), una vez podamos disponer de ligandos altamente selectivos para cada subtipo de receptor.
Referencias
Anwil, R. (1999). Metabotropic glutamate receptors: Electrophysiological properties and role in plasticity. Brain Research Reviews, 29, 83-120. [ Links ]
Belozertsseva, I. V. & Bespalov, A. Y. (1999). Effects of NMDA receptor channel blockade on aggression in isolated male mice. Aggressive Behavior, 25, 381-396. [ Links ]
Bespalov, A. Y., van Gaalen, M. M., Sukhotina, I., A., Wicke, K., Mezler, M., Schoemaker, H. & Gross, G. (2008). Behavioral characterization of the mGlu group II/III receptor antagonist, LY-341495, in animal models of anxiety and depression. European Journal of Pharmacology, 592, 96-102. [ Links ]
Brain, P F. (1975). What does individual housing mean to a mouse? Life Sciences, 16, 187-200. [ Links ]
Brain, IP F., Benton, D., Childs, G. & Parmigiani, S. (1981). The effect of the type of opponent in tests of murine aggression. Behavioural Processes, 6, 319-327. [ Links ]
Brain, P. F., Kusumorini, N. & Benton, D. (1991). "Anxiety" in laboratory rodents: A brief review of some recent behavioural developments. Behavioural Processes, 25, 70-80. [ Links ]
Brain, P. F., McAllister, K. H. & Walmsey, S. (1989). Drug effects on social behaviour: Methods in ethopharmacology. In A. A. Boulton, G. B. Baker & A. J. Greenshaws (Eds.), Neuromethods (pp. 687-739). NJ: Humana Press. [ Links ]
Cryan, J. F. & Dev, K. K. (2008). The glutamatergic system as a potencial therapeutic target for the treatment of anxiety disorders. En R. J. Blanchard, D. C. Blanchard, G. Griebel & D. Nutt (Eds.), Handbook of Anxiety and Fear, 17, 269-301. [ Links ]
Espert, R., Salvador, A., Navarro, J. F. & Simon, V. M. (1993). Effects of morphine hydrochloride on social encounters between male mice. Aggressive Behavior, 19, 377-383. [ Links ]
Ferraguti, F. & Shigemoto, R. (2006). Metabotropic glutamate receptors. Cell and Tissue Research, 326, 483-504. [ Links ]
Gómez, M. C., Carrasco, M. C. & Redolat, R. (2008). Differential sensitivity to the effects of nicotine and bupropion in adolescent and adult male OF1 mice during social interaction tests. Aggressive Behavior, 34, 369-379. [ Links ]
Gu, G., Lorrain, D. S., Wei, H., Cole, R. L., Zhang, X., Daggett, L. P et al. (2008). Distribution of metabotropic glutamate 2 and 3 receptors in the rat forebrain: Implication in emotional responses and central disinhibition. Brain Research, 1197, 47-62. [ Links ]
Helton, D. R., Tizzano J. P, Monn J. A., Schoepp D. D. & Kallman, M. J. (1998). Anxiolytic and side-effect profile of LY354740: A potent, highly selective, orally active agonist for group II metabotropic glutamate receptors. Journal of Pharmacology and Experimental Therapeutics, 284, 651-660. [ Links ]
Klodzinska, A., Chojnacka-Wojcik, E., Palucha, A., Bransky, P., Popik, P. & Pilc, A. (1999). Potential anti-anxiety, anti-addictive effects of LY 354740, a selective group II glutamate metabotropic receptors agonist in animal models. Neuropharmacology, 38, 1831-1840. [ Links ]
Kew, J. N. & Kemp, J. A. (2005). Ionotropic and metabotropic glutamate receptor structure and pharmacology. Psychopharmacology, 179, 4-29. [ Links ]
Kudryavtseva, N. N., Bondar, N. P. & Alekseyenko, O. V. (2000). Behavioral correlates of learned aggression in male mice. Aggressive Behavior, 26, 386-400. [ Links ]
Lee, Y., Duman, R. S. & Marek, G. J. (2006). The mGlu2/3 receptor agonist LY354740 suppresses immobilization stress-induced increase in rat prefrontal cortical BDNF mRNA expression. Neuroscience Letters, 398, 328-332. [ Links ]
Liebenauer, L. L. & Slotnick, B. M. (1996). Social organization and aggression in a group of olfactory bulbectomized male mice. Physiology and Behaviour, 60, 403-409. [ Links ]
Linden, A. M., Greene, S. J., Bergeron, M. & Schoepp, D. D. (2004). Anxiolytic activity of the mGlu2/3 receptor agonist LY354740 on the elevated plus maze is associated with the suppression of stress-induced c-Fos in the hippocampus and increases in c-Fos induction in several other stress-sensitive brain regions. Neuropsychopharmacology, 29, 502513. [ Links ]
Lujén-Miras, R. (2005). Receptores metabotrópicos de glutamato: nuevas dianas moleculares de enfermedades neurológicas y psiquiétricas. Revista de Neurología, 40, 43-53. [ Links ]
Lumley, L. A., Robinson, C. L., Slusher, B. S., Wozniak, K., Dawood, M. & Meyerhoff, J. L. (2004). Reduced isolation-induced aggressiveness in mice following NAALADase inhibition. Psychopharmacology, 171, 375-381. [ Links ]
Manzaneque, J. M. & Navarro, J. F. (1999). Behavioral profile of amisulpride in agonistic encounters between male mice. Aggressive Behavior, 25, 225-232. [ Links ]
Marek, G. J. (2004). Metabotropic glutamate 2/3 receptors as drugs targets. Current Opinion in Pharmacology, 4, 18-22. [ Links ]
Martín-López, M. & Navarro, J. F. (2002). Antiaggresive effects of zolpidem and zopiclone in agonistic encounters between male mice. Aggressive Behavior, 28, 416-425. [ Links ]
Martín-López, M., Puigcerver, A., Vera, F. & Navarro, J. F. (1993). Sulpiride shows an antiaggressive specific effect after acute treatment in male mice. Medical Science Research, 21, 595-596. [ Links ]
Martínez, M., Calvo-Torrent, A. & Paya-Cano, J. L. (1997). Effects on the behaviour of standard nonaggressive male mice opponents during agonistic encounters. Aggressive Behavior, 23, 179-181. [ Links ]
Matochik, J. A. (1988). Role of the main olfactory system in recognition between individual spiny mice. Physiology and Behavior, 42, 217-222. [ Links ]
Miczek, K. A., Maxson, S. C., Fish, E. W. & Faccidomo, S. (2001). Aggresive behavioral phenotypes in mice. Behavioural Brain Research, 125, 167-181. [ Links ]
Moore, N. A., Rees, G. & Monn, J. A. (1999). Effects of the group II metabotropic glutamate receptor agonist, LY354740 on schedule-controlled behaviour in rats. Behavioural Pharmacology, 10, 319-325. [ Links ]
Navarro, J. F. (1997). An ethoexperimental analysis of the agonistic interactions in isolated male mice: Comparison between OF.1 and NMRI strains. Psicothema, 9, 333-336. [ Links ]
Navarro, J. F. & Maldonado, E. (2004). Effects of acute, subchronic and intermittent MDMA (ecstasy) administration on agonistic interactions between male mice. Aggressive Behavior, 30, 84-91. [ Links ]
Navarro, J. F. & Pedraza, C. (1996). An ethopharmacological assessment of the effects of gammahydroxybutyrate (GHB) on agonistic interactions in male mice. Medical Science Research, 24, 817-819. [ Links ]
Navarro, J. F., Postigo, D., Martín-López, M. & Burón, E. (2006). Antiaggressive effects of MPEP, a selective antagonist of mGlu5 receptor, in agonistic interactions between male mice. European Journal of Pharmacology, 551, 67-70. [ Links ]
Navarro, J. F., Burón, E. & Martín-López, M. (2007). Antiaggressive effects of topiramate in agonistic encounters between male mice. Methods and Findings in Experimental and Clinical Pharmacology, 29, 195-198. [ Links ]
Navarro, J. F., De Castro, V. & Martín-López, M. (2008). JNJ16259685, a selective mGlu1 antagonist, suppresses isolation-induced aggression in male mice. European Journal of Pharmacology, 586, 217-220. [ Links ]
Navarro, J. F., Luque, M. J. & Martín-López, M. (2009a). Effects of L379268, a selective agonist of mGlu2/3 receptors, on isolation-induced aggression in male mice. The Open Pharmacology Journal, 3, 17-20. [ Links ]
Navarro, J. F., De Castro, V. & Martín-López, M. (2009b). Behavioural profile of selective ligands for mGlu7 and mGlu8 glutamate receptors in agonistic encounters between mice. Psicothema, 21, 475-479. [ Links ]
Nedergaard, M., Takano, T. & Hansen, A. (2002). Beyond the role of glutamate as a neurotransmitter. Nature, 3, 748-755. [ Links ]
Olivier, B. & Young, L. J. (2002). Animal Models of Aggression. En K. L. Davis, D. Charney, J. T. Coyle & C. Nemeroff (Eds.), Neuropsychopharmacology: The Fifth Generation of Progress (pp. 1699-1708). Philadelphia: Lippincott Williams & Wilkins. [ Links ]
Ozawa, S., Kayima, H. & Tsuzuki, K. (1998). Glutamate receptors in the mammalian central nervous system. Progress in Neurobiology, 54, 581-618. [ Links ]
Pedraza, C., Dévila, G., Martín-López, M. & Navarro, J. F. (2007). Anti-aggressive effects of GHB in OF.1 strain mice: Involvement of dopamine D2 receptors. Progress in Neuropsychopharmacology & Biological Psychiatry, 31, 337-342. [ Links ]
Schlumberger C, Schéfer, D, Barberi, C., More, L., Nagel, J., Pietraszek, M. et al. (2009). Effects of a metabotropic glutamate receptor group II agonist LY354740 in animal models of positive schizophrenia symptoms and cognition. Behavioural Pharmacology, 20, 56-66. [ Links ]
Schoepp, D. D., Johnson, B. G., Wright, R. A., Salhoff, C. R., Mayne, N. G., Wu, S. et al. (1997). LY354740 is a potent and highly selective group II metabotropic glutamate receptor agonist in cells expressing human glutamate receptors. Neuropharmacology, 36, 1-11. [ Links ]
Schoepp, D. D. (2001). Unveiling the functions of presinaptic metabotropic glutamate receptors in the central nervous system. Journal of Pharmacology and Experimental Therapeutics, 299, 12-20. [ Links ]
Schoepp, D. D., Wright, R. A., Levine, L. R., Gaydos, B. & Potter, W. Z. (2003). LY354740, an mGlu2/3 receptor agonist as a novel approach to treat anxiety/ stress. Stress, 6, 189-197. [ Links ]
Siegel, A., Bhatt, S., Bhatt, R. & Zalcman, S. (2007). The neurobiological bases for development of pharmacological treatments of aggressive disorders. Current Neuropharmacology, 5, 135-147. [ Links ]
Siever, L. J. (2008). Neurobiology of aggression and violence. American Journal of Psychiatry, 165, 429-442. [ Links ]
Vekovischeva, O. Y., Aitta-Aho, T., Echenko, O., Kankaanpaa, A., Seppala, T., Honkanen, A. et al. (2004). Reduced aggression in AMPA-type glutamate receptor GluR-A subunit-deficient mice. Genes, Brain and Behavior, 3, 253-265. [ Links ]
Vekovischeva, O. Y., Aitta-Aho, T., Verbitskaya, E., Sandnabba, K. & Korpi, E. R. (2007). Acute effects of AMPA-type glutamate receptor antagonists on intermale social behavior in two mouse lines bidirectionally selected for offensive aggression. Pharmacology Biochemistry & Behavior, 87, 241-249. [ Links ]

